淹水底泥反硝化消納面源氮潛力的影響因子分析
2022-06-25 09:51:32史禎琦周佳華張文娟佘冬立
中國農村水利水電 2022年6期
史禎琦,房 凱,周佳華,張文娟,佘冬立
(1.河海大學農業科學與工程學院,南京 210098;2.江蘇省宿遷市節約用水管理服務中心,江蘇 宿遷 223800;3.江蘇省水文水資源勘測局連云港分局,江蘇連云港 222004)
0 引 言
反硝化作用是活化氮循環的最后一步,其基本過程是:NO3-→NO2-→NO →N2O →N2[1]。微生物主導的反硝化作用能夠將生態系統固定的氮和人為活性氮以N2的形態返回到大氣氮庫中,因而反硝化過程是濕地生態系統氮素消納的主要途徑之一。反硝化過程影響因素眾多,目前關于溫度、pH、Eh、有機碳與活化氮有效性等環境因素的影響已開展較為豐富的研究[2,3],而對于水土界面生物變化影響下反硝化規律及其機制研究還有待深入。淹水土壤生態系統水土界面普遍生長有一層周叢生物,其系統內各種生物和非生物物質交織在一起,具有很強的生態穩定性,是反硝化發生的熱點區域[4]。周叢生物因素在反硝化過程中扮演雙重角色,一方面在新陳代謝過程中直接主導反硝化過程,另外一方面自身對周邊環境的作用反過來又影響其活性[5]。因此,分析反硝化過程影響因素時,不僅要考慮環境因子,還要綜合考慮水土界面周叢生物因子的影響及其共線性關系,探究反硝化過程主控因子[6]。
由于模型結構簡單、變量間關系易于說明等特點,多元線性回歸模型被廣泛運用于生態系統過程影響因子的分析。淹水底泥生態系統各組成部分相互影響、相互作用,導致量化后的環境因子之間不可避免的存在共線性,導致多元回歸模型穩定性不強。伍德與阿巴諾[7]在1983年提出的偏最小二乘回歸分析方法(Partial Least Square Regression)PLSR 模型兼具主成分分析、典型相關分析和一般最小二乘多元回歸分析等多種傳統多元統計方法的特點,可提取出對因變量解釋性最強的自變量,預測精度較高,結果更為可靠且整體性較強,能較好的解決環境因子、生物因子之間的共線性問題。因此,本研究通過培養實驗測定上覆水-周叢生物-底泥界面反硝化特征,運用PLSR 模型分析反硝化消納面源氮潛力的影響因子,提取出主控因子,為面源污染防控提供一定理論基礎。
1 材料和方法
1.1 供試土壤與實驗設計
培養實驗在河海大學江寧校區節水園區(N31°86′,E118°60′,海拔144 m)進行。供試的底泥pH(H2O)為7.6,砂粒(0.02~2 mm)、粉粒(0.002~0.02 mm)、黏粒(0~0.002 mm)的體積分數分別為31.5%、39.6%、28.9%,有機質含量為5.8 g/kg,總氮含量為9.3 mg/kg,總磷含量為79 mg/kg,總鉀含量為201 g/kg。
為研究淹水底泥反硝化消納面源氮潛力的影響因素,對供試底泥進行覆水培養。將供試底泥風干研磨過4 mm 篩網,去除石塊和大顆粒土塊,并將其充分攪拌,以獲得均勻的底泥樣品。將底泥樣品均分為56 個子樣品,以1.3 g/cm3的容重,30 cm的深度,填入直徑20 cm、深度40 cm 的培養盆中,加水覆蓋,水層深度5 cm,整個試驗周期中保持水層深度不變,每一培養盆隨機添加N 營養液(按照0、50、100、200、400、600、900 和1 300 kg/hm2尿素梯度添加,各N 營養液處理設置7 次重復隨機擺設)。在溫室中(25 ℃)培養60 d 后,水土界面持續生長形成一層明顯的周叢生物,其生物群落結構穩定,且由于水土界面N營養水平的差異,周叢生物膜生物量梯度差異明顯。
1.2 反硝化速率與環境因子測定
影響反硝化速率的因素主要從上覆水、周叢生物和底泥3部分考慮。試驗培養60 d 后,采用無擾動沉積物采樣器(有機樹脂玻璃管,內徑8 cm,外徑9 cm,高30 cm)采取各個處理上覆水5 cm、土層30 cm的土柱,柱底用黑色橡膠塞密封。將柱樣帶回實驗室后,沿玻璃棒緩慢加入與上覆水氮素含量一致的硝酸鉀溶液直至柱體內充滿液體,擰好蓋子,調整直至柱體中沒有任何氣泡。垂直放置于模擬原位淹水環境的培養裝置中,液面高出柱子約4~5 cm,然后連接進水管和出水管。采用膜進樣質譜法[8]測定柱樣反硝化速率,具體方法示意見圖1。

圖1 膜進樣質譜法測定反硝化速率示意圖Fig.1 Schematic diagram of measuring of denitrification rates by Membrane Inlet Mass Spectrometry
反硝化測定后,采用便攜式多參數測試儀(Hach Company,Loveland)測定上覆水中pH、溶解氧(DO);將水樣過濾后采用流動分析儀(Skalar Analytical,Breda,The Netherlands)測定上覆水體氨態氮(NH4+-N)和硝態氮(NO3--N)含量;采用液態碳氮元素分析儀(Vario Toc)測定可溶性有機碳(DOC)和總氮(TN(W))。將上覆水緩慢傾倒,收集水土界面周叢生物,測定參數包括群落結構比例、葉綠素(CHI)含量。利用消毒后的薄刀片將一定面積(3×3 cm 大小,約2 mm 厚)的周叢生物從載體表面剝離,放置于烘箱105 度烘干至恒重即為生物量(BIO)[9]。通過測序分析發現在前10 種優勢物種中,假單胞菌(Pseudomonas)與反硝化速率成顯著相關關系,因此將其納入上覆水影響因子中。輕輕刮取0.5 g 周叢生物膜樣品,采用FastDNA?SPIN Kit For Soil(MP Biomedicals,Santa Ana,CA)試劑盒提取土壤微生物總DNA,并采用高通量測序儀Miseq對其進行測序,使用16sr DNA進行高通量測序分析,測定底泥土壤反硝化細菌基因豐度(NIRK、NOSZ)、氨氧化古菌(AOA)、氨氧化細菌(AOB);取底泥土樣20 g,用2 mol/L KCl 溶液浸提后,采用流動分析儀(Skalar Analytical,Breda,The Netherlands)測定氨態氮(NH4+-N)、硝態氮(NO3
--N)。將土樣風干后研磨,過100 目篩網,使用CNS 元素分析儀(型號Vario MAX)測定全氮[TN(S)]。
1.3 數據處理
在構建PLSR 模型之前,運用SPSS statistics 25.0 進行Zscore 數據標準化、皮爾遜相關性分析,滿足模型的建立要求。在偏最小二乘回歸運算過程中,將交叉驗證作為確定PLSR 組分的標準,進行如下計算。
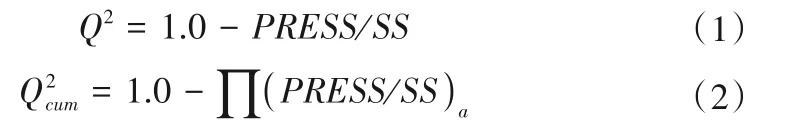
式中:Q2為每一成分可解釋的因變量方差比;Q2cum為偏最小二乘回歸中所有成分可解釋的因變量方差比;PRESS為預測誤差平方和;SS為剩余平方和;a為偏最小二乘回歸成分數量。
運用均方根誤差(Root Mean of Squared Error,RMSE)來對模型進行校正。PLSR 模型的建立和計算均在SIMCA 14.1 軟件中進行,將整理好的數據導入SIMCA 中,設定反硝化速率為因變量,環境影響因子為自變量。應用Autofit 對模型進行自動擬合,擬合過程中軟件會自動進行交叉驗證,使最佳解釋能力(R2)與模型預測能力(Q2)相平衡。偏最小二乘回歸建模過程中使用變量投影重要性指標(VIP)表示自變量對因變量預測的重要程度。回歸系數(RCs)是反映偏最小二乘回歸模型中自變量對因變量影響的方向和程度。
2 結果與分析
2.1 反硝化速率與環境因子變異特征
淹水底泥添加N 營養液且經過60 d 培養后,水土界面環境與生物因子均發生了顯著變化(p<0.05)(表1)。上覆水和底泥中氮含量(NO3--N、NH4+-N 和總氮TN)隨添加N 營養液梯度呈顯著遞增趨勢(p<0.05),且各氮含量因子均呈中等變異強度。在N 營養激化作用下,底泥中參與氮素循環過程的微生物群落同樣發生顯著變化,底泥反硝化細菌(NIRK 亞硝酸鹽還原酶基因和NOSZ 氧化亞氮還原酶基因)變化范圍分別為0.53~47.63×105copies/ug 和2.58~389.33×105copies/μg,氨氧化古菌(AOA)和氨氧化細菌(AOB)變化范圍分別為0.03~1.05×105copies/μg和0.47~10.48×105copies/μg。與此同時,淹水底泥在經過60 d培養后,水土界面生長有一層穩定的生物聚集體,即周叢生物,其系統內各種生物,如微生物(包括細菌和真菌)和小型動植物等生物群與非生物物質(如鐵錳氧化物等)交織形成自然的一層生物膜。通過對周叢生物生物量(BIO)、假單胞菌(PSE)和葉綠素(CHI)含量的分析表明,周叢生物的生長受水土界面氮營養狀況影響顯著,隨初始添加氮營養濃度增大,周叢生物各指標均顯著增大。
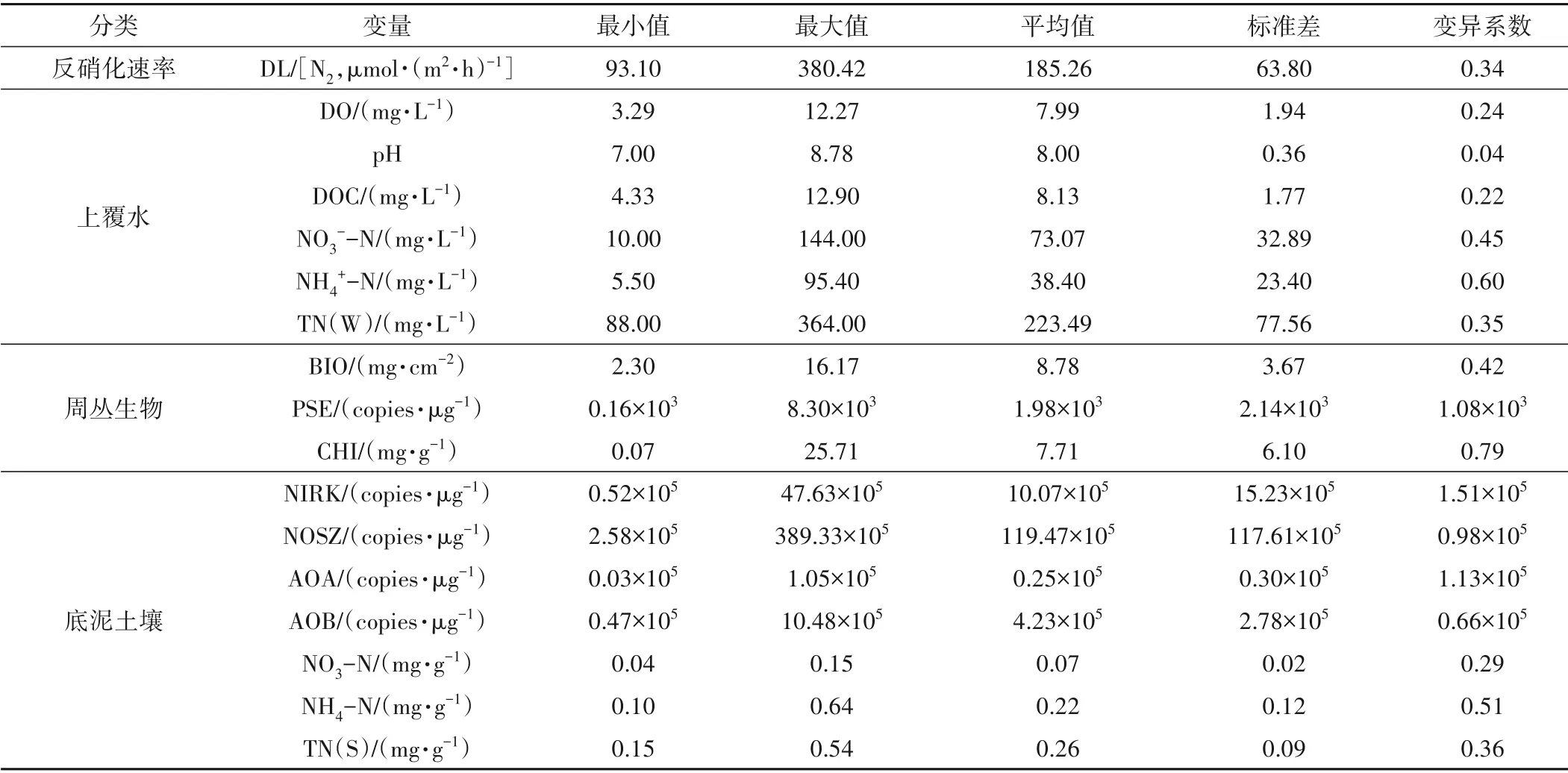
表1 反硝化速率和環境影響因子數據特征Tab.1 Statistical summary of denitrification loss and associated factors
淹水底泥氮素營養狀況及周叢生物的變化顯著影響土柱水土界面的反硝化速率(p<0.05)。反硝化速率的變化范圍為93.1~380.42 μmol/(m2·h),平均值為185.26 μmol/(m2·h),屬于中等變異強度。相關分析結果表明,反硝化速率與氮營養梯度和周叢生物各指標均呈顯著正相關關系(p<0.05)。不同于對傳統“上覆水-沉積物”兩相界面脫氮過程的認識,本研究也進一步表明,水土界面的周叢生物也是淹水底泥反硝化發生的熱點區域。
2.2 反硝化速率變異PLSR模型構建
根據培養試驗結果,構建了反硝化速率與水土界面各環境因子間的PLSR 模型(表2)。模型提取了2 個偏最小二乘回歸成分,第一成分解釋反硝化速率75.86%變異,第二成分解釋10.97%變異,模型累計變異性解釋度為86.83%,表明該模型能夠反映反硝化速率變異整體信息,Qcum2>0.5,模型能較好的對數據集進行模擬和預測。PLSR 權重圖展示了反硝化速率和環境影響因子之間的關系,權重越大的因子與反硝化速率相關度越高(圖2)。將2 個主成分權重綜合分析,得出各因子對反硝化過程的影響作用。上覆水中pH、DOC、NO3--N 和NH4+-N 對反硝化過程起促進作用且與反硝化速率相關度較高,DO 則抑制反硝化過程;周叢生物各指標均在第一主成分系統中表現出對反硝化速率的促進作用,而底泥中NIRK、NO3--N 和TN(S)在兩個主成分系統中均表現出對反硝化速率顯著正向貢獻作用。
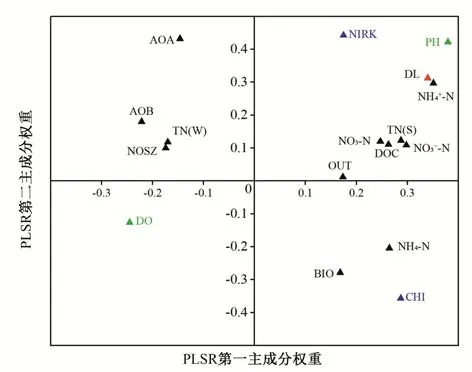
圖2 反硝化速率PLSR模型第一、第二主成分權重圖Fig.2 Weight plots of the first and second PLSR components for denitrification rate
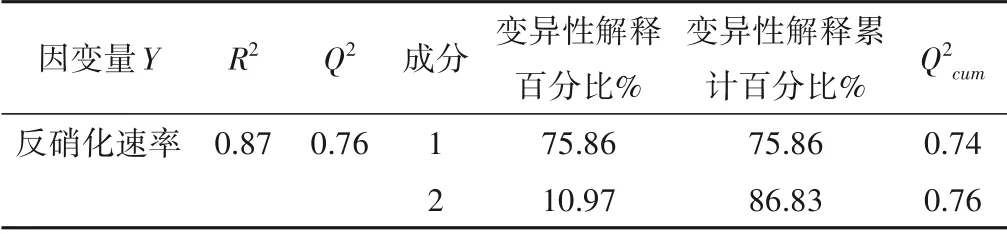
表2 反硝化速率變異的PLSR模型Tab.2 PLSR model of Denitrification Loss
2.3 反硝化速率的影響因子分析
在PLSR 模型中,VIP值表示自變量對因變量預測的重要程度(圖3)。根據VIP值將各變量分為高影響因子(VIP值>1)、中影響因子(0.8<VIP值<1)和低影響因子(VIP值<0.8)。“上覆水-周叢生物-沉積物”三相界面中,各界面影響反硝化速率的主要因素不同。上覆水中,高影響因子主要有酸堿度(pH,VIP=1.531,回歸系數=0.261)、氨根離子濃度(NH4+-N,VIP=1.378,回歸系數=0.212)、硝酸根離子濃度(NO3--N,VIP=1.146,回歸系數=0.135)和溶解性有機碳(DOC,VIP=1.014,回歸系數=0.124),上覆水中高影響因子皆對反硝化過程起促進作用;周叢生物中影響反硝化速率的高影響因子為葉綠素含量(CHI,VIP=1.219,回歸系數=-0.014);底泥沉積物中各環境因子VIP值均<1,其中5個因子屬于中等影響因子,包括底泥氨態氮(NH4-N,VIP=0.989,回歸系數=0.026)、全氮[TN(S),VIP=0.951,回歸系數=0.036]、硝態氮(NO3-N,VIP=0.949,回歸系數=0.085)、反硝化細菌(亞硝酸鹽還原酶基因)(NIRK,VIP=0.91,回歸系數=0.197)、氨氧化細菌(AOB,VIP=0.9,回歸系數=-0.019)和氧化亞古菌(AOA,VIP=0.809,回歸系數=0.085),底泥中各影響因子對反硝化過程主要為正向影響。
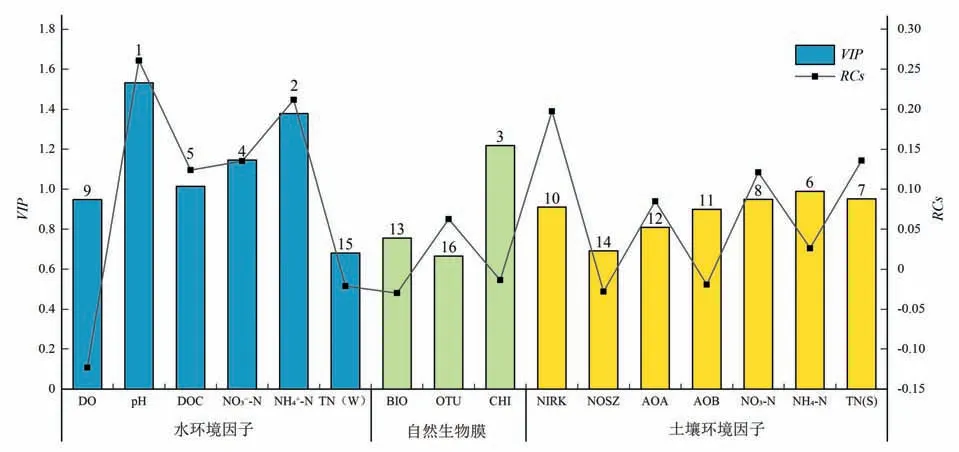
圖3 反硝化速率PLSR模型中各因子VIP及RCs圖Fig.3 Variable importance for the projection and regression coefficient of each predictor of PLSR for denitrification rate
3 討 論
上覆水中,pH、DOC、NO3--N 和NH4+-N 濃度對反硝化過程有正向影響,DO 對反硝化有負向影響。DO 的平均值為7.99 mg/L(變異系數24%),屬于富氧環境,而反硝化細菌是化能異養兼性菌,當水環境中DO>6.4 mg/L 時,會使得分子氧直接供氧[10],從而導致反硝化速率降低;Cho 等[11]發現在淹水土壤中,當O2消耗完全后,才會產生N2O。Fillery 等[12]發現當pH 上升時,N2O 還原酶活性降低,而Aamer 等[13]則認為當pH>6.6 時,對N2O 還原酶的影響會減弱;陳曦等[2]研究認為當pH 在7.80 左右時,厭氧氨氧化速率最高;本研究培養實驗pH 變化幅度較小(均值為8),進一步驗證了當pH 趨于穩定后(大于6.6),對N2O還原酶的抑制減弱促進了反硝化作用[14]。絕大多數反硝化細菌是化能異養型,Lescure等[15]發現單糖對反硝化細菌的刺激更強,DOC 則為反硝化細菌同時提供了電子受體及能量來源,而易分解的DOC又刺激了土壤微生物呼吸作用,加速了厭氧環境的形成。因此,DOC 從提供能量和創造厭氧環境兩個方面促進反硝化速率[16]。NO3--N和NH4+-N直接參與反硝化過程,NO3--N濃度的提高激發了微生物活性,加速底泥土壤的反硝化進程,驗證了佘冬立等[6]研究結果,同時二者作為反應物和氧源影響著化學反應方向及微生物的呼吸作用[17]。底泥沉積物中,NO3--N 對反硝化過程有較大的正向影響。NO3--N 作為反應電子受體及反應產物,直接影響土壤反硝化速率;而在pH 值較高的環境中,NO3--N 含量過高則會抑制N2O 還原酶(此抑制作用可逆)[18],提高反硝化氣體產物的N2O/N2的比率。底泥NIRK、AOA、AOB 屬于中影響因子,其中NIRK 對反硝化速率具有顯著的正向作用。
相對于單一微生物群落,周從生物具有多種特殊的“集體功能”,如具有多種酶促協同作用,周叢生物含有多種酶蛋白如堿性磷酸酶、脲酶和過氧化氫酶等[19],這些酶活性能直接或間接影響反硝化速率。周叢生物中CHI 為影響反硝化速率的主要因子。CHI 是衡量藻類數量的指標,日間藻類的光合作用造成水體富氧以及pH 的升高,夜間藻類自身及其他微生物的呼吸作用又會降低DO,DO 的具體數值實際上是生態系統中藻類光合作用與其他微生物生命活動耗氧平衡的結果。其生長過程中產生的胞外多糖[20]為反硝化細菌提供了好氧—缺氧界層,在它大量凋亡分解的過程中又需要較多的氧氣甚至會形成缺氧環境[21],同時還會提高DOC 和氨氮濃度[22],促進反硝化反應。此外,Thind 和Rowell 的研究表明[3],藻類所吸收的氮素的40%會在體內迅速礦化,最后釋放到環境中刺激作物生長,這意味著藻類保護了部分氮素阻止其參與反硝化過程。同時,周叢生物可以通過影響上覆水和底泥環境因子而導致反硝化速率發生變化。例如,周叢生物光合作用釋放大量氧氣[4],提高上覆水和表層沉積物氧化還原電位,從而有利于硝化反應;硝態氮擴散到毗鄰的厭氧層,在反硝化細菌的作用下轉化成氮氣[23],最終損失到大氣中。
4 結 論
(1)淹水底泥水土界面氮素營養狀況及周叢生物的變化顯著影響沉積物反硝化速率(p<0.05)。上覆水各因子的影響作用顯著,其中,pH、NH4+-N 、NO3--N 和DOC 對反硝化過程起促進作用且與反硝化速率相關度較高,DO 則抑制反硝化過程;周叢生物各指標均在第一主成分系統中表現出對反硝化速率的促進作用,而底泥中NIRK、NO3--N 和TN(S)在兩個主成分系統中均表現出對反硝化速率顯著正向貢獻作用。
(2)“上覆水-周叢生物-沉積物”三相界面中影響反硝化速率的主要因素和程度不同。上覆水整體對反硝化過程的影響程度較大,周叢生物次之,底泥沉積物影響程度相對較小。主成分分析表明各因子之間互相影響,其中周叢生物同時影響上覆水和底泥土壤,因此在淹水底泥反硝化消納氮素的研究過程中,應重視并厘清生物界面的影響機制及方式。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
當代陜西(2020年13期)2020-08-24 08:22:02
航空世界(2020年10期)2020-01-19 14:36:20
制造技術與機床(2017年5期)2018-01-19 02:49:17
濰坊學院學報(2016年2期)2016-12-01 13:00:11
中國塑料(2016年3期)2016-06-15 20:30:00
