植物生長調節劑優化不同品種冬小麥成熟胚再生體系
2019-07-02 12:10:30姚富泉董魯浩王悅李興國王芳別曉敏
山東農業科學 2019年5期
姚富泉 董魯浩 王悅 李興國 王芳 別曉敏


摘要:小麥愈傷組織再生能力是遺傳轉化的重要基礎。為優化小麥成熟胚再生體系,以科農199、輪選987和濟麥22三個冬小麥品種的成熟胚為外植體,在愈傷組織分化階段添加不同濃度的植物生長調節劑TDZ(N-苯基-N′-1,2,3-噻二唑-5-脲,thidiazuron),統計和分析分化率。結果表明,TDZ最適響應濃度存在品種間差異,科農199、輪選987和濟麥22成熟胚愈傷組織分化的最適TDZ濃度分別為1.5、1.0、0.5mg/L,愈傷組織分化率分別為84.61%、71.88%和71.77%,均極顯著高于對照。本研究結果可為優化冬小麥成熟胚愈傷組織再生體系提供參考。
關鍵詞:植物生長調節劑;TDZ;成熟胚;冬小麥;再生體系
中圖分類號:S512.1+1文獻標識號:A文章編號:1001-4942(2019)05-0019-05
隨著分子生物學的發展,生物技術育種手段日趨完善,但再生效率低、遺傳轉化困難仍是提高小麥(TriticumaestivumL.)遺傳轉化效率的瓶頸。已有研究發現,影響小麥組織培養再生頻率的主要因素包括外植體[1,2]、基因型[3,4]、培養基類型[5-7]等。
改良培養基是提高小麥再生頻率的手段之一,添加植物生長調節劑可以達到提高小麥成熟胚再生頻率的目的。作為植物生長調節劑,N-苯基-N′-1,2,3-噻二唑-5-脲(thidiazuron,TDZ)對體細胞胚胎發生等具有重要作用,還具有調節其他植物激素和生理活性物質的作用[8]。Huetteman和Preece[9]發現低濃度TDZ可通過去除頂端優勢誘導器官發生,促進不定芽或側芽的形成。在植物離體培養過程中,無論是外植體脫分化形成愈傷組織,還是愈傷組織經過器官發生途徑或胚狀體途徑形成再生植株,都是激素協同作用的結果[10]。在多種植物培養體系中添加TDZ都能誘導愈傷組織的形成,而且細胞增殖速率大大高于添加其他植物生長調節物質[11]。
目前小麥遺傳轉化所用外植體主要有幼胚、成熟胚、花藥和胚軸等[1,2,12-16]。與其他外植體相比,成熟胚具有取材方便、不受季節限制、可大量獲得、操作簡單等優點,已得到廣泛應用,然而再生頻率較低是其應用的主要限制因素之一[14]。有研究表明植物生長調節劑預處理對小麥成熟胚的出愈率無顯著影響,但能夠顯著促進愈傷組織分化[17]。培養基中添加TDZ對冬小麥成熟胚愈傷組織再生能力影響的研究尚未見報道。本試驗通過研究TDZ對科農199、輪選987和濟麥22成熟胚分化效率的影響,明確小麥成熟胚再生的最適TDZ濃度,為提高小麥成熟胚再生效率提供參考。
1材料與方法
1.1試驗材料
選用冬小麥商業栽培品種科農199、輪選987和濟麥22,均由山東農業大學生命科學學院植物發育分子生物學實驗室保存并提供。秋季在山東農業大學試驗基地種植,第二年六月份收獲成熟種子進行試驗。
1.2培養基成分和TDZ濃度設置
成熟胚預培養用Adi培養基,愈傷組織誘導用XCSX培養基,愈傷組織分化用XCFH培養基[7](表1)。在小麥成熟胚分化階段設置添加不同濃度TDZ處理,分別為0.5、1.0、1.5、2.0、2.5mg/L,以不添加TDZ的培養基做對照,每個濃度梯度設置3個重復,每個重復接種25個外植體。所有培養基均在1.1kg/cm2(121℃)條件下高壓滅菌15min。
1.3成熟胚培養
選取成熟飽滿的小麥種子,在超凈工作臺上先用70%乙醇表面消毒10min,無菌水沖洗1次,再用25%次氯酸鈉滅菌25min,無菌水沖洗3次,置于25℃培養箱中過夜。次日早晨,再用25%次氯酸鈉滅菌15min,無菌水沖洗5次,將成熟胚刮碎后接種于Adi培養基上,25℃黑暗條件下培養7d后,轉移到XCSX培養基上弱光培養14d,胚性愈傷組織及時轉移到XCFH培養基上進行光照培養(光照強度3500lx,光周期為16h光照、8h黑暗)[7]。
1.4數據統計
小麥成熟胚愈傷組織在分化培養基培養14d后統計分化率,并進行方差分析。愈傷組織分化率(%)=分化出綠點的愈傷組織數/接種外植體數×100。
試驗數據利用DPS6.85軟件采用Tukey法進行統計分析。
2結果與分析
2.1不同濃度TDZ對科農199成熟胚愈傷組織分化率的影響
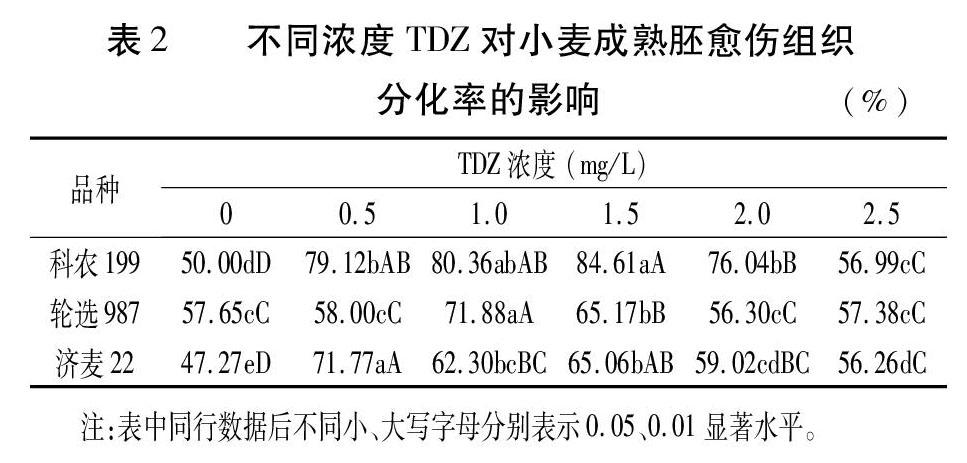
添加TDZ處理的科農199小麥成熟胚愈傷組織分化率均極顯著高于對照,且隨著TDZ添加濃度的升高,愈傷組織分化率呈現先高后低的趨勢,在添加濃度為1.5mg/L時愈傷組織分化率達到84.61%的最高值,與2.0、2.5mg/L濃度處理相比有極顯著差異,與0.5mg/L濃度處理相比達到顯著性差異水平(表2,圖1)。表明TDZ在0.5~2.5mg/L范圍內對科農199成熟胚愈傷組織誘導出芽具有正向調控作用,0.5~1.5mg/L低濃度的促進效果更顯著。
2.2不同濃度TDZ對輪選987成熟胚愈傷組織分化率的影響
隨著TDZ添加濃度的升高,輪選987的愈傷組織分化率呈現先高后低的趨勢,在添加濃度為1.0mg/L時愈傷組織分化率達到71.88%的最高值,與其他添加濃度相比達到極顯著差異水平;其次為1.5mg/L添加濃度,愈傷組織分化率為65.17%,也極顯著高于其他濃度。添加濃度為0.5、2.0、2.5mg/L時,愈傷組織分化率與對照無顯著性差異(表2,圖2)。可見,輪選987成熟胚愈傷組織在1.0~1.5mg/LTDZ處理時分化率顯著提高。
2.3不同濃度TDZ對濟麥22成熟胚愈傷組織分化率的影響
添加TDZ可以顯著提高濟麥22成熟胚的愈傷組織分化率(表2,圖3)。在添加濃度為0.5mg/L時,愈傷組織分化率達到71.77%的最高值,顯著高于其他處理;其次為1.5、1.0mg/L添加濃度,愈傷組織分化率明顯高于其余濃度處理;添加濃度為2.0mg/L和2.5mg/L的愈傷組織分化率較低。結果表明,添加0.5~1.5mg/LTDZ對濟麥22成熟胚愈傷組織分化率的提高幅度較大。
3討論與結論
TDZ是人工合成的植物生長調節劑,具有促進細胞分裂和植株再生的能力,有利于不定芽或側芽的形成,可以代替生長素或者細胞分裂素來誘導體細胞胚胎發生[8,9,18]。作為一種有效的形態發生調節劑在植物組織培養快繁體系中得到了廣泛應用,應用范圍遍及草本植物和木本植物,許多被認為難以誘導植株再生的植物種類都可以對TDZ產生應答[19]。
單獨使用TDZ即可誘導花生(Arachishypogaea)體胚的發生,表明TDZ有類似生長素的生物學功能[20]。Mok等[21]以棉豆(Phaseoluslunatus)和煙草(Nicotianatabacum)愈傷組織為外植體研究發現,TDZ具有相當于甚至超過腺嘌呤類細胞分裂素的活性。在誘導天竺葵(Pelargoniumhortorum)體細胞胚胎發生時,TDZ的添加引起乙烯水平升高,推測TDZ可能通過乙烯途徑影響植物再生能力[22]。此外,徐華松等[23]發現在TDZ誘導體細胞胚胎發生時脯氨酸也起到關鍵作用,推測TDZ誘導體細胞胚胎發生的作用機制也可能與脅迫反應有關。以上研究說明,TDZ在促進植物再生過程中,通過調節內源植物生長激素起作用[11,24]。
本研究以3個冬小麥品種的成熟胚為外植體,在誘導愈傷組織出芽過程中添加0.5~2.5mg/L的TDZ,發現TDZ低濃度添加可以明顯提高3個冬小麥品種的愈傷組織分化能力,但不同品種對TDZ添加濃度的響應不同。在本試驗條件下,科農199、輪選987、濟麥22的最適TDZ添加濃度分別為1.5、1.0、0.5mg/L。本試驗結果可為提高冬小麥成熟胚愈傷組織再生能力提供參考。
參考文獻:
[1]葉興國,徐惠君,趙樂蓮,等.組織培養途徑改良定型小麥品種的研究[J].作物學報,1998,24(3):310-314.
[2]武麗敏,鄭有良,魏育明,等.小麥轉基因研究進展[J].四川農業大學學報,2003,21(2):176-181.
[3]FennellS,BohorovaN,GinkelM,etal.Plantregenerationfromimmatureembryosof48eliteCIMMYTbreadwheats[J].TheoreticalandApplidGenetics,1996,92(2):163-169.
[4]MamruthaH,KumarR,VenkateshK,etal.Genetictransformationofwheat-presentstatusandfuturepotential[J].JournalofWheatResearch,2014,6(2):107-119.
[5]CarmanJG,JeffersonNE,CampbellWF.InductionofembryogenicTriticumaestivumL.calli.I.Quantificationofgenotypeandculturemediumeffects[J].PlantCell,TissueandOrganCulture,1987,10:101-103.
[6]RamageC,WilliamsR.Mineralnutritionandplantmorphogenesis[J].InVitroCellular&DevelopmentalBiology-Plant,2002,38(2):116-124.
[7]別曉敏,杜麗璞,徐惠君,等.培養基中CuSO4和Fe鹽濃度對小麥胚培養再生效果的影響[J].植物遺傳資源學報,2011,12(6):975-981.
[8]陳云鳳,張春榮,黃霞,等.TDZ對植物體細胞胚胎發生的作用[J].植物生理學通訊,2006,42(1):127-133.
[9]HuettemanC,PreeceJ.Thidiazuron:apotentcytokininforwoodyplanttissueculture[J].PlantCell,TissueandOrganCulture,1993,33(2):105-119.
[10]LoschiavoF,PittoL,GiulilanoG.DNAmethylationofembryogeniccarrotcellculturesanditsvariationascausedbymutation,differentiation,hormonesandhypomethylatingdrugs[J].TheoreticalandAppliedGenetics,1989,77(3):325-331.
[11]徐曉峰,黃學林.TDZ:一種有效的植物生長調節劑[J].植物學通報,2003,20(2):227-237.
[12]HakamN,UdupaS,RabhaA,etal.Efficientcallusinductionandplantletsregenerationinbreadwheatusingimmatureandmatureembryos[J].InternationalJournalofBiotechnologyResearch,2015,3(1):1-9.
[13]郭志江,張鳳路,王金明,等.農桿菌介導小麥成熟胚體系的優化[J].中國農學通報,2008,24(4):181-185.
[14]陶麗莉,殷桂香,葉興國.小麥成熟胚組織培養及遺傳轉化研究進展[J].麥類作物學報,2008,28(4):713-718.
[15]ShariatpanahiM,BelogradovaK,HessamvaziriL,etal.Efficientembryogenesisandregenerationinfreshlyisolatedandculturedwheat(TriticumaestivumL.)microsporeswithoutstresspretreatment[J].PlantCellReports,2006,25(12):1294-1299.
[16]YuH,WangW,WangY,etal.Highfrequencywheatregenerationfromleaftissueexplantsofregeneratedplantlets[J].AdvancesinBioscienceandBiotechnology,2012,3(1):46-50.
[17]王麗,陳耀鋒,張月琴,等.有機添加物對小麥成熟胚愈傷組織分化特性的影響[J].西北農業學報,2014,23(3):45-49.
[18]ThomasJ,KattermanF.Cytokininactivityinducedbythidiazuron[J].PlantPhysiology,1986,81(2):681-683.
[19]MurthyB,SaxenaP.Somaticembryogenesisandplantregenerationofneem(AzadirachtaindicaA.Juss.)[J].PlantCellReports,1998,17(6/7):469-475.
[20]MurchS,SaxenaP.Theroleofprolineinthidiazuroninducedsomaticembryogenesisofpeanut[J].InVitroCellular&DevelopmentalBiology-Plant,1999,35(1):102-105.
[21]MokM,MokD,ArmsstrongD.CytokinactivityofN-phenye-N′-1,2,3-thidiazol-5-ylurea(thidiazuron)[J].Phytochemistry,1982,21(7):1509-1511.
[22]HutchinsonM,SaxenaP.Roleofpurinemetabolisminthidiazuron-inducedsomaticembryogenesisofgeranium(Pelargonium×hortorum)hypocotylcultures[J].PhysiologiaPlantarum,1996,98(3):517-522.
[23]徐華松,徐九龍.TDZ在植物組織培養中的作用[J].廣西植物,1996,16(1):77-80.
[24]周俊彥,郭扶興.苯基脲衍生物的細胞分裂素活性[J].植物生理學通訊,1990(4):7-13.
