G蛋白信號調節因子-13通過調控β-catenin抑制結直腸癌細胞增殖、遷移和侵襲
2022-06-21 01:45:08項友群方冠金亦翔楊凱潘貽飛
溫州醫科大學學報 2022年6期
項友群,方冠,金亦翔,楊凱,潘貽飛
溫州醫科大學附屬第一醫院結直腸肛門外科,浙江 溫州 325015
結直腸癌(colorectalcancer, CRC)是世界范圍內最常見的消化系統腫瘤之一[1]。2015 年我國CRC新發病例近38萬例,病死近19萬例,發病率和病死例數均居世界首位[2]。CRC的進展通常伴隨著癌基因和抑癌基因的突變,表達量的變化以及表觀遺傳學變化等[3-4]。雖然對CRC進展過程中基因水平的表達調控已有了廣泛的研究,但是CRC在分子水平的發生發展機制仍有待闡明。明確CRC增殖、遷移和侵襲的相關分子與相關信號通路,對于CRC的診斷與治療具有重要的意義。G蛋白信號調節因子-13(regulatorofGproteinsignaling13, RGS13)基因位于人類1號染色體長臂3區1帶,編碼19.1kDa的蛋白質。RGS13是Gαi和Gαo的GTPase激活蛋白,使結合的GTP轉變為GDP,從而調節G蛋白的活性來抑制G蛋白的信號轉導[5-6]。作為一個與細胞信號通路密切相關的因子,其在腫瘤發生發展中的作用尚不完全清楚。本研究通過TCGA數據庫發現RGS13在人類CRC組織中低表達,并在CRC臨床組織樣本和細胞系中進行驗證,探究了其在CRC細胞增殖、遷移和侵襲中的作用及其可能的下游機制。
1 材料和方法
1.1 材料
1.1.1 細胞:結直腸正常細胞系FHC(CRL-1831)購自美國模式培養物集存庫(Americantypeculturecollection, ATCC);CRC細胞系SW480(CBP60019)、SW620(CBP60036)、DLD-1(CBP60037)和HCT116(CBP60028)購自南京科佰生物科技有限公司,均經過STR分型鑒定且鑒定無誤。細胞均使用ATCC推薦的培養基培養,培養基均補充有10%胎牛血清、1%雙抗和1%L-谷氨酰胺,并置于37℃、5%CO2細胞培養箱。
1.1.2 質粒:β-catenin、RGS13過表達質粒購自武漢淼靈生物科技有限公司。慢病毒法構建穩定轉染的細胞株。將適量293T細胞接種于6孔板中,使用PolyJetTMDNA體外轉染試劑按照說明書操作。使用工作濃度的嘌呤霉素篩選穩定轉染的細胞。
1.1.3 抗體:RGS13抗體(PA5-68626)購自美國ThermoFisher公司,β-catenin(51067-2-AP)、Flag(66008-4-Ig)、GAPDH(60004-1-Ig)抗體購自美國Proteintech公司。
1.1.4 引物:RGS13引物(正向:5’-AGATTTACATCCAGCCACAGTCCC-3’,反向:5’-GACTTTAGAAATCTGGGGTAGGAAT-3’),細胞周期蛋白-D1(cyclinD1,CCND1)引物(正向:5’-GCTGCGAAGTGGAAACCATC-3’,反向:5’-CCTCCTTCTGCACACATTTGAA-3’),c-Myc引物(正向5’-CAGTAGCACTGCGCGATAGA-3’,反向:5’-TGCTTCAGACCTTCGTGACC-3’),基質金屬蛋白酶7(matrixmetalloproteinases7,MMP7)引物(正向:5’-GAGT GAGCTACAGTGGGAACA-3’,反向:5’-CTATGACGCGGGAGTTTAACAT-3’),均由上海桑尼生物科技有限公司合成。
1.1.5 臨床組織樣本:實驗使用的134對CRC組織及其癌旁組織樣本取自溫州醫科大學附屬第一醫院結直腸肛門外科,均為通過病理學檢測確診為CRC,通過外科手術治療取得的臨床組織。所有程序均通過溫州醫科大學附屬第一醫院倫理委員會審查[編號:(2021)第(063)號]。
1.1.6 數據庫組織樣本:下載TCGA 數據庫(https://tcga-data.nci.nih.gov/)中的結腸癌(TCGA-COAD)的exon-SeqV2測序數據(Level3,rawcount)和臨床數據。其中,數據庫中可用于差異分析的CRC與癌旁配對樣本共41對,用于生存分析含無病生存期(disease-freesurvival, DFS)信息的樣本共379例。
1.2 方法
1.2.1 免疫組織化學染色:將制作好的切片脫蠟、復水、抗原修復,1×Tris緩沖液(1×TBS)清洗后3%H2O2封閉30min后用5%BSA封閉30min,甩去后4℃孵育一抗過夜,回收一抗,然后37℃下孵育二抗30min,清洗后鏈霉親和素-生物素復合物(SABC)孵育30min,滴加DAB顯影液于組織上顯色合適時間,ddH2O終止后蘇木素染色,自來水靜置10min,脫色和透明后制片,光學顯微鏡下觀察蛋白的染色情況,軟件分析單位面積下的IOD值(陽性著色)。
1.2.2 蛋白質印跡法(Westernblot):配好10%的十二烷基硫酸鈉-聚丙烯酰胺(SDS-PAGE)膠后,收集細胞,處理并定容蛋白樣品,上樣20μL,濃縮膠80V電泳15min,分離膠120V電泳1.5h,使用快速轉膜儀轉膜15min,5%脫脂牛奶封閉1h,1×TBS清洗PVDF膜后4℃孵育一抗過夜后回收一抗,4℃孵育二抗3h后回收二抗,使用ECL顯影法顯影。Typhoon7000掃膜儀掃描PVDF膜,保存分析圖像結果。
1.2.3 實時熒光定量聚合酶鏈反應(real-timequantitativepolymerasechainreaction, RTqPCR):TRIzol法提取總RNA并使用SuperScriptTMIV反轉錄試劑盒,按照說明書反轉錄出cDNA后,使用SYBRGreen試劑盒按照說明書進行qPCR,以GAPDH作為內參。最后置于Q6qPCR儀中檢測并用2-ΔΔCt法分析。
1.2.4 ATP細胞活力檢測:先細胞種板,96 孔板中每孔加入1000個細胞,用0.1%培養基處理細胞12h。之后按照對應天數用10%培養基培養細胞,棄培養基后將各25μL的ATP試劑和PBS混合加入孔中。振蕩后1200r/min離心5min。靜置后轉至黑色96孔板,加樣后生物發光儀檢測,保存數值并分析結果。
1.2.5 軟瓊脂克隆集落形成實驗:先在6孔板中鋪2mL下層瓊脂膠,凝固后鋪1mL混有1×104個細胞的上層膠,6孔板放入37℃、5%CO2培養箱中培養。用倒置顯微鏡5倍鏡拍照并計數含32個細胞以上的克隆,計算細胞集落數形成率并分析結果。
1.2.6 細胞遷移侵襲實驗:專用小室上室加400μL0.1%FBS培養基,下室加700μL完全培養基。上室接種細胞5×104個,培養24h后,棄去培養基,之后用4%多聚甲醛固定細胞20min,甲醇通透20min,吉姆薩染色液染色20min,擦去未從上室穿出到下室的細胞,倒置顯微鏡拍照計數并統計分析。
1.2.7 蛋白降解實驗:先細胞種板,6孔板中每孔加入3×105個細胞,細胞貼壁后分別用0.1%FBS的培養基、MG132試劑(10mmol/L)處理細胞12h、8h后,加入放線菌酮(cycloheximide, CHX)試劑(50μg/mL)分別處理細胞0h、3h、6h、12h,Westernblot檢測蛋白降解情況。
1.3 統計學處理方法 采用SPSS18.0統計軟件進行數據分析,用GraphPadPrism5.0軟件作圖。計量資料以±s表示,兩獨立樣本數據比較采用非配對t 檢驗,兩配對樣本數據比較采用配對t 檢驗,多組間數據分析采用單因素方差分析,生存分析采用Log-rank法,非正態分布計量資料采用非參數檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 RGS13在CRC組織與細胞系中表達降低且與CRC患者的分期和無病生存期顯著相關 本研究選取了TCGA數據庫中41對配對的CRC組織樣本和癌旁正常組織樣本的轉錄組測序結果,發現在CRC組織樣本中RGS13的mRNA表達水平顯著低于癌旁正常組織(P<0.01),見圖1A;對收集到的134對CRC臨床組織樣本進行RT-qPCR實驗,得到了與上述分析一致的結果(P<0.01),見圖1B。在蛋白水平上,本研究通過對134對CRC臨床組織樣本進行免疫組織化學實驗并分析,發現在CRC組織樣本中,RGS13蛋白表達量顯著低于癌旁正常組織(P<0.01),見圖1C。為了方便進一步研究RGS13的功能,本研究在常用的CRC細胞系及正常結直腸細胞系中進行了Westernblot檢測。結果表明,相較于正常結直腸細胞系FHC,在HCT116、SW480等CRC細胞系中RGS13的蛋白表達水平均顯著下調,見圖1D。
為了進一步研究RGS13的表達量與CRC惡性程度的關系,本研究對134對臨床組織免疫組織化學實驗的結果按照病例的臨床分期分組并分析。結果表明,CRC低分期(I+II期,70例)病例的RGS13的蛋白表達量高于CRC高分期(III+IV期,64例)病例的RGS13的蛋白表達量(P<0.01),見圖1E。為了進一步闡明RGS13的表達量與CRC患者預后的關系,本研究對TCGA數據庫中的379例CRC患者的DFS數據進行分析,發現RGS13高表達組(相對表達量大于均值0.758,86例)的DFS顯著高于RGS13低表達組(相對表達量小于均值0.758,293例,P<0.01),見圖1F。
2.2 RGS13的下調可顯著促進CRC細胞的增殖、遷移與侵襲 為了在生物學功能上驗證RGS13的下調是否促進CRC的惡性進展,本研究選用RGS13下調較為顯著的HCT116和SW480細胞系,構建了RGS13的過表達穩轉細胞株,見圖2A。ATP實驗表明,過表達RGS13的HCT116和SW480細胞,其增殖指數顯著低于對照空載細胞(P<0.01),見圖2B、圖2C。進行軟瓊脂克隆集落形成實驗檢測細胞的錨定非依賴生長能力,結果顯示,過表達RGS13的HCT116和SW480細胞,其細胞克隆集落的形成能力顯著低于對照空載細胞(P<0.01),見圖2D、圖2E。細胞遷移侵襲實驗結果顯示,過表達RGS13的HCT116和SW480細胞,其遷移與侵襲能力顯著低于對照空載細胞(P<0.01),見圖2F、圖2G。
2.3 RGS13可下調β-catenin的表達從而影響相關轉錄因子的表達,進而抑制CRC細胞系的增殖、遷移與侵襲 Westernblot結果表明,在過表達RGS13的HCT116和SW480細胞中,β-catenin的表達量低于對照空載細胞,見圖3A。使用蛋白酶體阻斷劑MG132預處理過表達RGS13的HCT116細胞和對照空載細胞后,再使用蛋白合成抑制劑CHX處理不同的時間。結果發現,過表達RGS13的HCT116細胞的β-catenin的降解速率高于對照空載細胞,見圖3B。
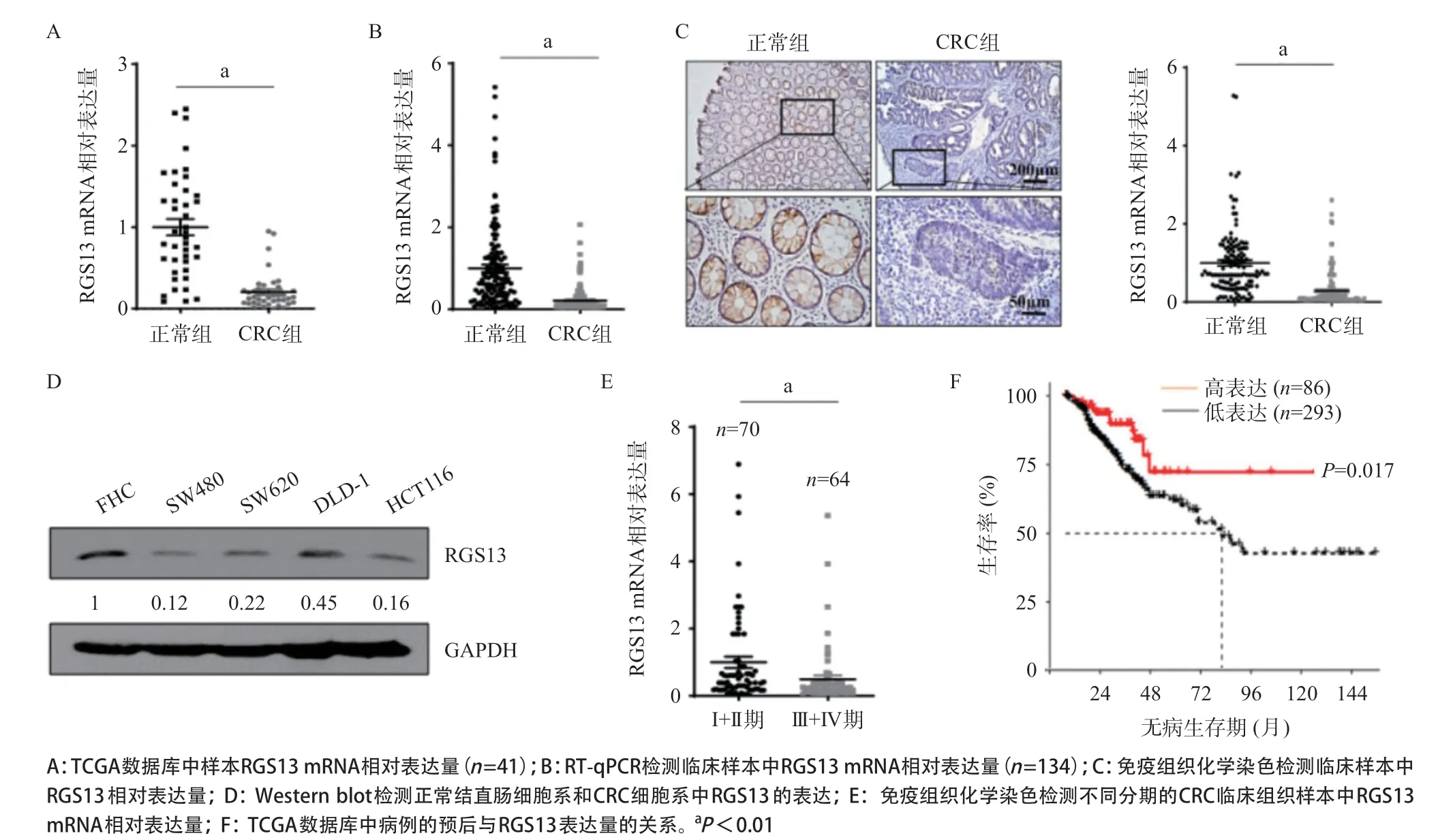
圖1 RGS13在CRC組織與細胞系中表達降低且與CRC患者分期和無病生存期相關
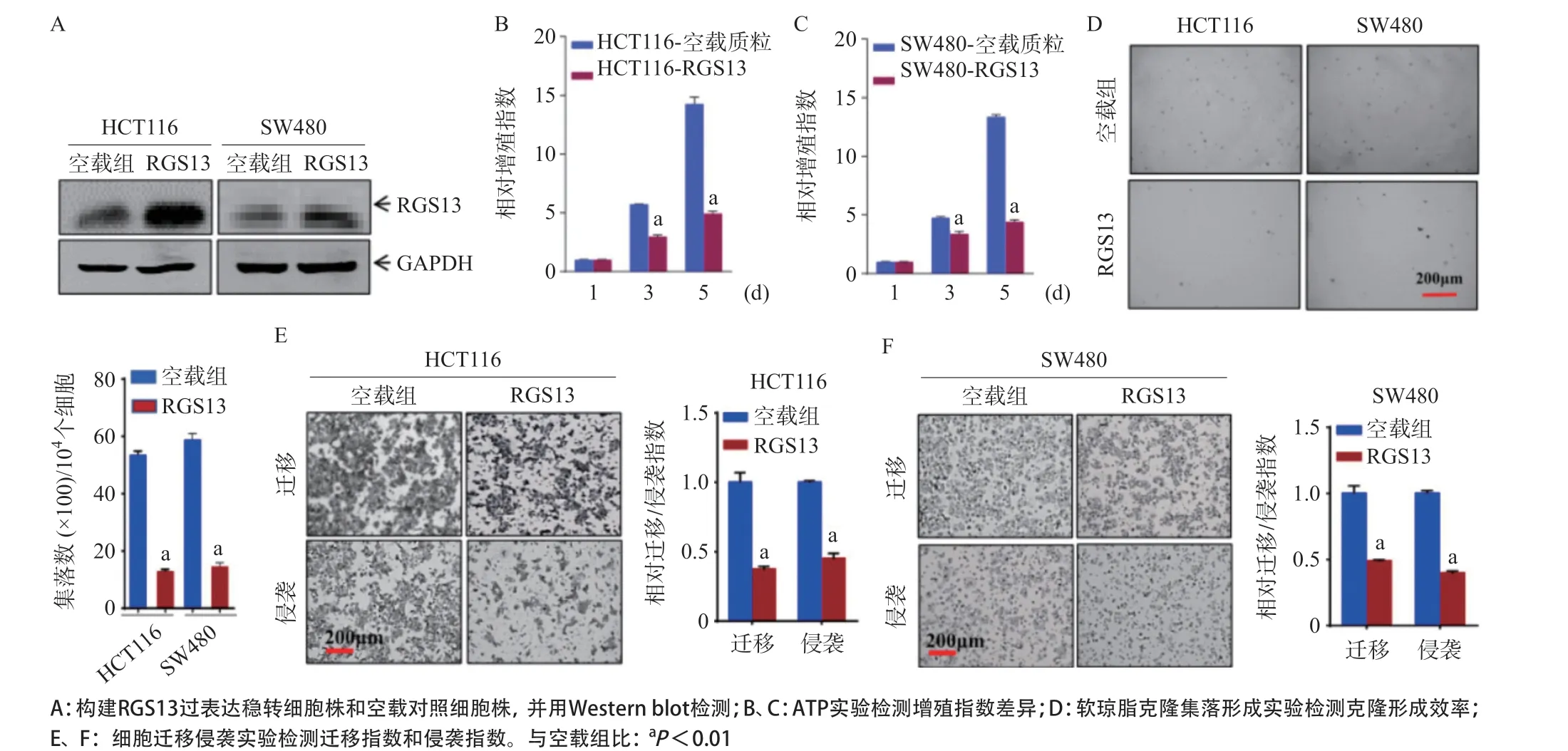
圖2 RGS13的下調可顯著促進CRC細胞的增殖、遷移與侵襲
RT-qPCR結果表明,在過表達RGS13的HCT116細胞中,CCND1、c-Myc和MMP7的mRNA水平較對照空載細胞均發生了顯著下調(P<0.01),見圖3C。為了進一步驗證,本研究在過表達RGS13的HCT116細胞中回轉了β-catenin的表達,見圖3D。再次進行RTqPCR發現,相較僅過表達RGS13的HCT116細胞,過表達β-catenin逆轉了CCND1、c-Myc和MMP7的mRNA水平(P<0.01),見圖3E。在生物學功能方面,ATP實驗、軟瓊脂克隆集落形成實驗和細胞遷移侵襲實驗共同表明,過表達β-catenin的HCT116-RGS13細胞,其增殖活力、集落形成能力和遷移與侵襲能力相較HCT116-RGS13對照空載細胞均得到了提高(P<0.01),見圖3F、圖3G、圖3H。
3 討論
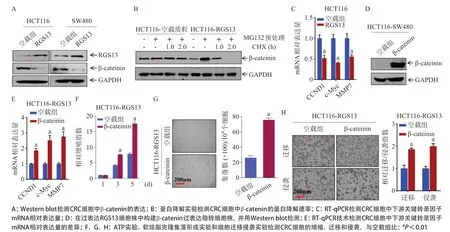
圖3 RGS13下調β-catenin的表達以影響相關轉錄因子的表達而抑制CRC細胞系的增殖、遷移與侵襲
異常的Wnt/β-catenin信號在CRC等癌癥研究中被大量報道,并且普遍認為這種異常的信號級聯是CRC發生發展的關鍵因素[7],90%的CRC患者都可以發現該通路的突變[8]。胞質中的β-catenin通常需要Wnt信號的激活,包括磷酸化、泛素化等一系列過程,導致β-catenin破壞復合物解聚,抑制β-catenin降解[9],導致其在核內積累,并促進許多癌基因如c-Myc、MMP7和CCND1的轉錄[10-11]。近年來,有研究發現了調節β-catenin穩定性的不依賴Wnt的機制,如在人CRC中發現APC和RNF43的功能喪失突變,以及CTNNB1和RSPO2/3的功能獲得性突變等[12]。本研究發現RGS13在CRC中表達下調,增強了β-catenin的穩定性,這提示RGS13可能通過不依賴Wnt的途徑調節β-catenin的表達,并且,本研究發現β-catenin的表達升高增加了癌基因c-Myc、MMP7和CCND1的轉錄活性,且促進了CRC細胞的增殖、遷移與侵襲,這可能是RGS13下調促進CRC惡性進展的重要分子機制。
RGS13是G蛋白信號(RGS)家族的調節因子,包含一個特征性的保守RGS結構域。研究表明axin2RGS域中的一個位點能阻止axin2 聚合體的形成,對該位點的點突變會導致axin2聚合體的形成,從而增強對β-catenin信號的抑制;同時該研究發現了一種短肽,其可通過促進聚合體的形成從而促進β-catenin的降解,抑制β-catenin下游基因的轉錄,進而抑制CRC細胞的增殖[13]。另外一項研究也發現了一種小分子KYA1797K,其可以結合axin的RGS結構域,誘導形成β-catenin破壞復合物,促進β-catenin降解,從而抑制結直腸癌細胞的增殖[14]。β-catenin的降解速率調控是Wnt/β-catenin信號通路的關鍵調控方式,本研究發現含RGS結構域的RGS13的過表達可促進β-catenin降解,從而影響下游相關因子的表達,這提示RGS13可能通過RGS結構域促進β-catenin的降解,具體的作用機制有待進一步的驗證。
綜上,本研究認為RGS13或通過直接結合或間接方式下調β-catenin的胞質穩定性,進而降低下游關鍵癌基因的轉錄活性,從而在CRC的進展中發揮抑癌作用,而RGS13的下調將導致CRC的惡性進展。隨著研究的不斷深入,RGS13有望作為分子靶點用于CRC的診斷與治療。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
發明與創新(2016年38期)2016-08-22 03:02:52
