人氨基肽酶N在豬丁型冠狀病毒感染HEK293細胞中的作用
2022-05-30 07:33:00趙玉佳宋代麗張路文李施倩文翼平杜森焱顏其貴文心田曹三杰黃小波
畜牧獸醫學報 2022年5期
趙玉佳,陳 汭,宋代麗,張路文,肖 黛,李施倩,文翼平,伍 銳,趙 勤,杜森焱,顏其貴,文心田,曹三杰,2,3,黃小波,2,3*
(1.四川農業大學動物醫學院豬病研究中心,成都 611130;2.農業農村部獸用藥物與獸醫診斷技術四川科學觀測實驗站,成都 611130;3.四川農業大學國家級動物類實驗教學示范中心,成都 611130)
丁型冠狀病毒(deltacoronavirus)是新發現的冠狀病毒科(),丁型冠狀病毒屬()成員,可以感染哺乳動物和禽類。2009年,Woo等發現3種禽丁型冠狀病毒:夜鶯冠狀病毒HKU11、畫眉冠狀病毒HKU12和文鳥冠狀病毒HKU13。2012年,Woo等在豬和鳥群中鑒定出7種新型丁型冠狀病毒,對豬冠狀病毒HKU15-44和HKU15-155毒株13、和基因序列分析顯示其與亞洲豹貓冠狀病毒相似性高達99.8%,說明PDCoV在野生小型哺乳動物和豬之間可能存在跨種傳播。2014年,美國暴發豬丁型冠狀病毒(porcine deltacoronavirus,PDCoV)感染。隨后,韓國、加拿大、泰國、越南、日本和中國等國家也報道PDCoV引起的仔豬腹瀉性疾病。當前,已報道的PDCoV毒株全基因組序列相對保守,序列分析表明,PDCoV可能起源于麻雀丁型冠狀病毒(sparrow deltacoronavirus,SpCoV)。此外,美國報道的4種新型SpCoV與PDCoV親緣關系更為密切,表明PDCoV在豬和禽類之間也可能發生跨種傳播。
PDCoV是引起豬腸道疾病的主要病原,可感染各年齡階段的豬,感染仔豬出現嘔吐、腹瀉、脫水和死亡等臨床癥狀,影響全球養豬業的健康發展。人工接種PDCoV還可感染牛、雞和火雞等多種動物。體外試驗證實,病毒也可感染豬源細胞(LLC-PK、PK15、ST、IPEC和IPI-2I)、人源細胞(Huh7和Hela)、禽源細胞(LMH和DF-1)、牛源細胞(PBK和PBH)、猴源細胞(Vero-CCL81)和犬源細胞(MDCK)等多種細胞,顯示PDCoV具有廣泛的跨宿主傳播風險。
氨基肽酶N(aminopeptidase N,APN),又稱CD13,是一種膜結合的金屬蛋白酶,可與冠狀病毒S蛋白結合,介導病毒入侵宿主細胞。甲型冠狀病毒中,傳染性胃腸炎病毒(transmissible gastroenteritis virus,TGEV)、貓冠狀病毒(feline coronavirus,FCoV)、犬冠狀病毒(canine coronavirus,CCoV)和人冠狀病毒229E(human coronavirus 229E,HCoV-229E)被鑒定出以APN作為功能受體。此外,APN作為冠狀病毒受體具有種屬特異性。有研究表明,HCoV-229E以hAPN作為受體,而不能以豬APN(porcine APN,pAPN)作為受體。貓APN(feline APN,fAPN)可與TGEV、CCoV、HCoV-229E和FCoV結合。但是,APN在PDCoV入侵宿主細胞的過程中是否發揮受體功能存在爭議。Li等構建APN基因敲除細胞證明APN是PDCoV感染多種動物細胞的功能受體,病毒可通過S蛋白的S1結構域與APN的催化區域發生互作。Wang等也證明,pAPN在PDCoV入侵細胞中發揮受體功能。與之相反,部分研究結果卻證明,APN不是PDCoV的受體。也有研究顯示,APN雖不是PDCoV的關鍵受體,但其能影響PDCoV的早期感染和增殖過程。另外,Stoian等卻認為APN是PDCoV感染細胞的一個非必需細胞受體。
為驗證hAPN在PDCoV復制中的作用,本研究首先證實PDCoV可感染HEK293細胞,再進一步構建hAPN基因敲除細胞系和hAPN過表達質粒,驗證hAPN在PDCoV復制中的作用。通過同源建模和分子對接模擬PDCoV S 蛋白與hAPN蛋白的相互作用,為闡述PDCoV的細胞入侵機制和跨種傳播提供新的理論依據。
1 材料與方法
1.1 細胞、病毒和主要試劑
HEK293、HEK293T和ST細胞由本實驗室凍存;PDCoV四川分離株CHN-SC2015(GenBank收錄號:MK355396.1)由本實驗室分離、鑒定和保存。pCMV-SPORT6-ANPEP質粒由本實驗室構建保存;兔抗ACTB多克隆抗體(AC026),HRP-羊抗兔IgG(AS014),HRP-羊抗鼠IgG(AS003):武漢Abclonal公司;鼠抗ANPEP單克隆抗體(sc-166105):Santa Cruz公司;兔抗PDCoV N多克隆抗體由本實驗室制備保存。
1.2 PDCoV感染HEK293細胞
將PDCoV以MOI=0.1接種60 mm培養皿中的HEK293細胞,37 ℃吸附1 h,PBS洗2遍,加維持液繼續培養。于0、12、24、36和48 h取樣,通過RT-qPCR和Western blot檢測PDCoV感染HEK293細胞后的病毒含量;將PDCoV感染HEK293細胞24 h后的細胞培養物凍融3次后,連續傳至4代,通過RT-PCR檢測PDCoV在HEK293細胞上的增殖情況,TCID檢測不同代次的病毒滴度。
1.3 hAPN基因敲除細胞系的構建與鑒定
參考hAPN基因(NC_000015)序列,利用網絡在線工具(http://chopchop.cbu.uib.no/)設計一對sgRNA(表1)。合成的sgRNA經退火處理,連接到B Ⅰ酶切的線性化載體lenti CRISPR v2,構建同時表達Cas9和sgRNA的慢病毒轉移質粒。用Lipofectamine 3000將質粒按重組質粒∶psPAX2∶pMD2.G=5∶3∶2的比例共轉染至HEK293T細胞中,48 h后收上清。待T25細胞瓶中HEK293細胞長至50%時,感染慢病毒,36 h后,用含1 μg·mL嘌呤霉素(puromycin)的完全培養基進行抗性篩選;有限稀釋法挑選hAPN基因敲除細胞系,將其命名為hAPN。通過測序、RT-qPCR和Western blot鑒定hAPN在HEK293細胞上的敲除情況。
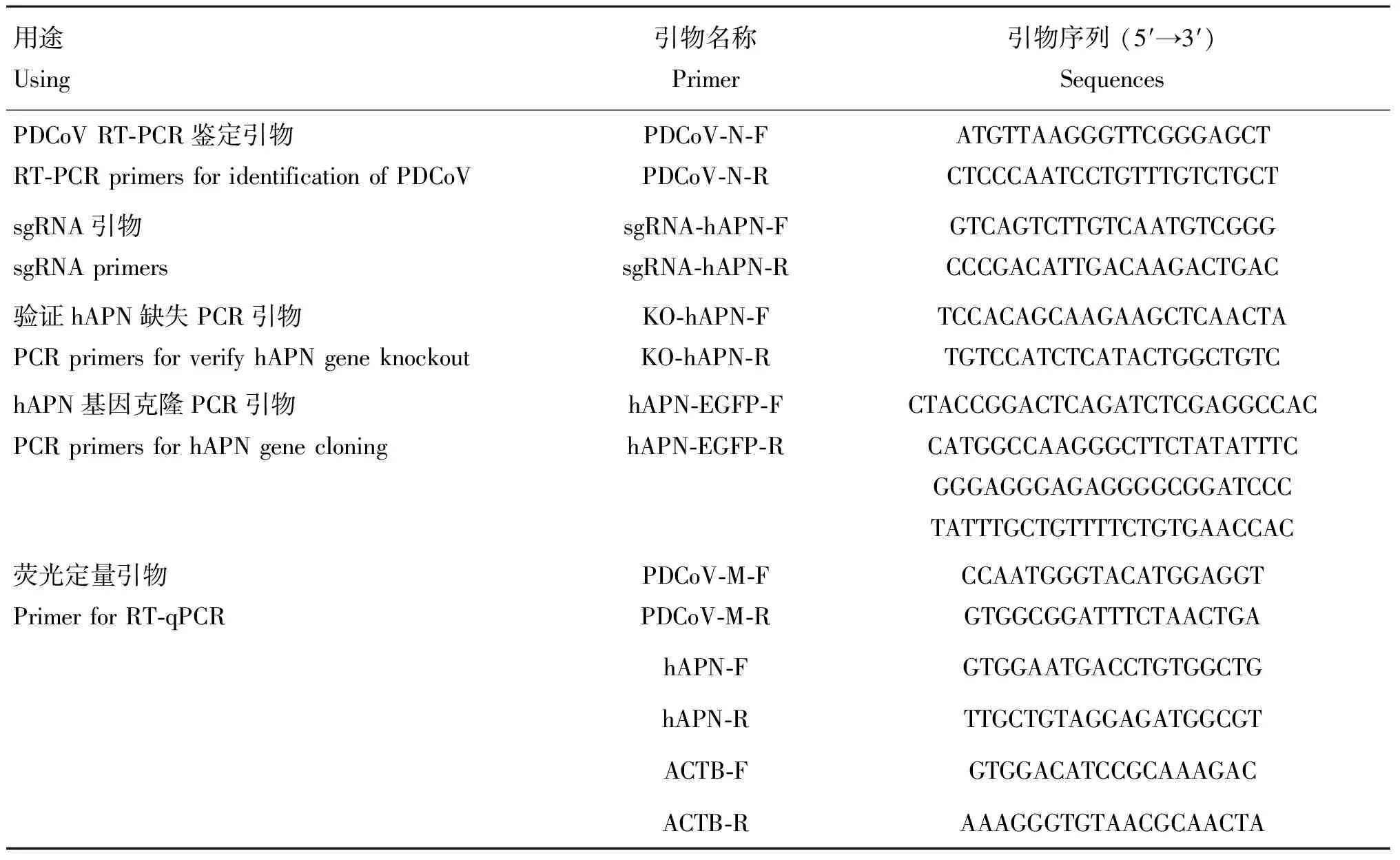
表1 相關引物序列
1.4 細胞活性鑒定
按照10·孔的細胞數將野生型細胞(hAPN)和敲除細胞(hAPN)接種96孔細胞板,24 h后,避光條件下,每孔加入10 μL CCK-8試劑,37 ℃孵育1 h,測定450 nm的吸光度。計算細胞存活率,細胞存活率=[(As-Ab)/(Ac-Ab)]×100%,As:試驗孔(hAPN細胞孔);Ac:對照孔(hAPN細胞孔);Ab:空白孔(培養基)。
1.5 敲除hAPN對PDCoV復制的影響
待60 mm細胞培養皿中hAPN和hAPN長滿單層后,PBS洗2遍,將PDCoV以MOI=0.1的劑量感染細胞,37 ℃孵育1 h,棄病毒液,每孔加入4 mL維持液,于24 h收集細胞懸液和蛋白,細胞懸液用RT-qPCR檢測基因轉錄水平;蛋白樣品用Western blot檢測N蛋白表達水平。
1.6 過表達hAPN對PDCoV復制的影響
針對hAPN基因的CDs區設計引物,經PCR擴增后連接到載體pIRES2-EGFP,構建hAPN過表達真核載體(命名為pIRES2-EGFP-hAPN)。待HEK293細胞長至50%時,用Lipofectamine 3000分別轉染4 μg質粒pIRES2-EGFP和pIRES2-EGFP-hAPN至HEK293細胞;轉染24 h后,觀察綠色熒光蛋白表達情況;收集細胞和細胞蛋白,分別用RT-qPCR和Western blot檢測hAPN表達。以MOI為0.1的PDCoV感染轉染pIRES2-EGFP-hAPN的HEK293細胞,同時設轉染pIRES2-EGFP空載的HEK293細胞為對照,24 h后,收集細胞懸液和蛋白,細胞懸液通過RT-qPCR檢測基因轉錄水平;蛋白樣品通過Western blot檢測N蛋白表達水平。
1.7 同源建模與分子對接
hAPN蛋白(PDB ID:4FYQ)、PDCoV S蛋白(PDB ID:6B7 N)和HCoV-229E S(PDB ID:6U7H)的整體結構從蛋白質數據庫獲取(https://www.rcsb.org)。用SWISS-MODEL(https://swissmodel.expasy.org/)手動構建PDCoV S蛋白和HCoV-229E S蛋白的單體結構和受體結構域(receptor binding domain,RBD),用PyMOL對hAPN、PDCoV S和HCoV-229E S蛋白的三維結構進行可視化分析;用MEGA6、ESPrit 3.0(http://espript.ibcp.fr/ESPript/ESPript/index.php)和PyMOL的align功能對PDCoV和HCoV-229E S蛋白的RBD進行序列和結構比對。用分子對接方法模擬PDCoV S1蛋白與hAPN蛋白的相互作用。
1.8 TCID50測定
取100 μL病毒液,按照10倍梯度進行倍比稀釋,共稀釋7個梯度,每個稀釋度設置8個重復,同時設置空白對照。待96孔細胞板中的ST細胞長滿單層后,PBS洗2遍,加入100 μL稀釋好的病毒液,37 ℃孵育1.5 h,棄病毒液,加入150 μL含5 μg·mL胰酶的維持液,37 ℃繼續培養。每日觀察細胞病變,連續觀察4 d,按照Reed&Muench方法計算TCID。
1.9 統計學分析
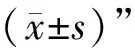
2 結 果
2.1 PDCoV在HEK293細胞中的增殖情況
將PDCoV以MOI=0.1感染HEK293細胞,收取0、12、24、36和48 h的樣品,RT-qPCR檢測病毒含量。結果發現,病毒感染細胞12~36 h時,病毒快速增殖,36 h達到頂峰;36~48 h,出現下降趨勢(圖1A);此外,在0~24 h,病毒N蛋白表達水平也迅速增加(圖1C)。將PDCoV在HEK293細胞上連續傳至4代,RT-PCR檢測發現當傳至第3代時,只能檢測到很弱的條帶,而傳至第4代時,檢測不到條帶(圖1B);Western blot 檢測發現,PDCoV傳至2代時,仍可檢測到很弱的病毒N蛋白表達水平(圖1D);TCID結果表明,PDCoV在HEK293細胞上第1代滴度約為4.36 lg TCID·mL,傳至2代時,病毒滴度稍微下降,約為3 lg TCID·mL(圖1E),表明PDCoV可感染HEK293細胞,但是不能在HEK293細胞上穩定傳代。
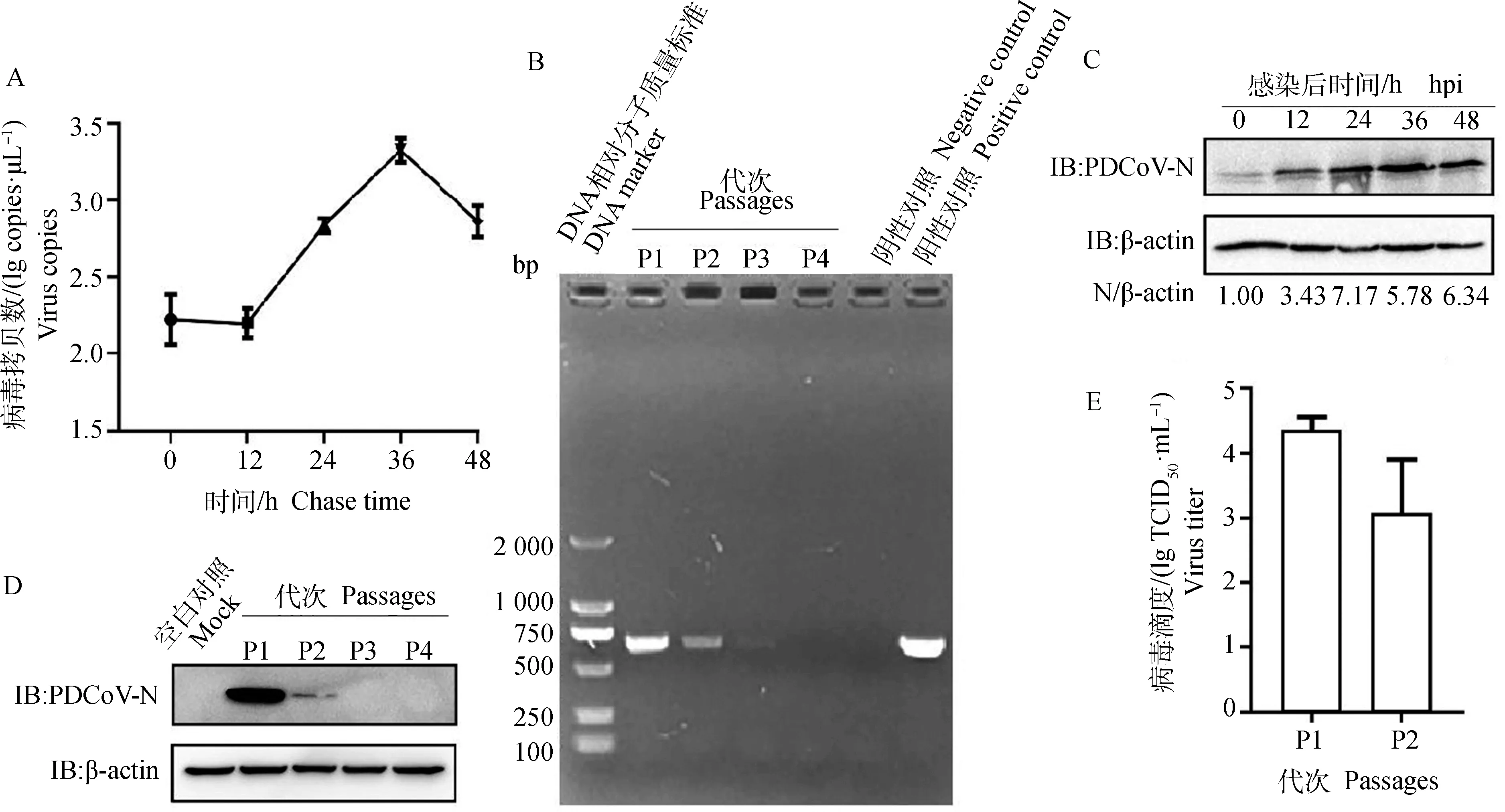
A.RT-qPCR檢測PDCoV在HEK293細胞上的增殖情況;B.RT-PCR檢測PDCoV在HEK293細胞上的傳代;C.Western blot檢測PDCoV感染細胞不同時間點的N蛋白表達水平;D.Western blot檢測PDCoV感染HEK293細胞不同代次的N蛋白表達水平;E.TCID50檢測PDCoV感染HEK293細胞不同代次的病毒滴度
2.2 hAPN基因敲除細胞系的構建和鑒定
用CRISPR/Cas9技術構建hAPN基因敲除細胞系,通過測序、RT-qPCR和Western blot檢測hAPN在HEK293細胞上的敲除情況。測序結果顯示,hAPN細胞系在PAM序列前出現明顯的重疊峰,且存在23個堿基的連續缺失(圖2A);RT-qPCR和Western blot結果顯示,在hAPN細胞上幾乎檢測不到hAPN基因表達(圖2B、C),表明hAPN在HEK293細胞上缺失成功。細胞活性檢測發現,hAPN和hAPN的活性無差異(圖2D),表明敲除hAPN對HEK293細胞活性無影響。
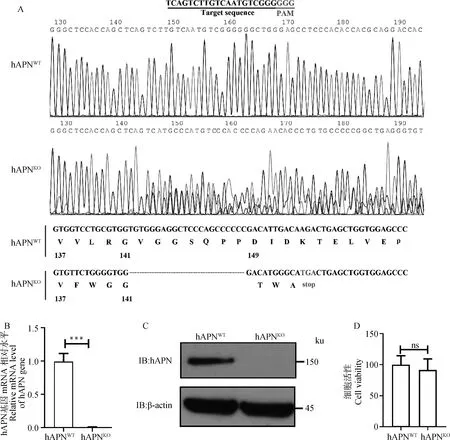
A.hAPNWT和hAPNKO細胞中hAPN基因的序列測定;B.RT-qPCR檢測hAPNKO細胞的hAPN基因mRNA水平(***.P<0.001);C.Western blot檢測hAPNKO細胞的hAPN蛋白表達水平;D.CCK-8檢測hAPNWT和hAPNKO細胞的細胞活性(ns.P>0.05)
2.3 敲除hAPN可降低PDCoV復制
為驗證hAPN敲除對PDCoV復制的影響,將PDCoV以MOI=0.1感染hAPN和hAPN細胞,24 h后,測定基因轉錄水平和N蛋白表達水平。結果表明,hAPN基因敲除后,導致基因mRNA水平下調約75%(圖3 A),N蛋白表達水平下降約66%(圖3 B),證實hAPN敲除能夠顯著抑制PDCoV復制。
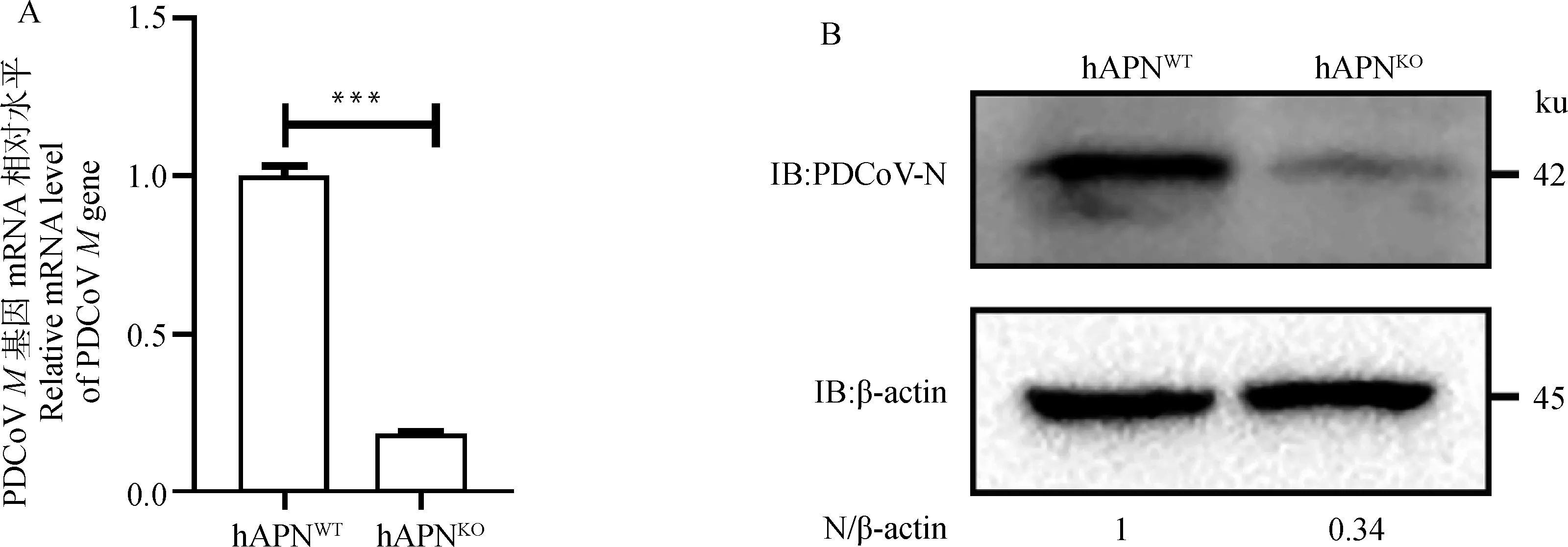
A.RT-qPCR檢測敲除hAPN后PDCoV M基因mRNA水平(***.P<0.001);B.Western blot檢測敲除hAPN后PDCoV N蛋白表達水平
2.4 過表達hAPN可促進PDCoV復制
為進一步驗證hPAN過表達對PDCoV復制的影響,構建hAPN真核表達質粒pIRES2-EGFP-hAPN,轉染至HEK293細胞中,24 h后,可以觀察到明顯的綠色熒光(圖4 A)。RT-qPCR和Western blot檢測發現,hAPN在HEK293細胞中成功表達(圖4 B、C)。將pIRES2-EGFP和pIRES2-EGFP-hAPN分別轉染HEK293細胞24 h后,接種PDCoV,病毒感染24 h后,RT-qPCR和Western blot檢測表明,過表達hAPN導致M基因mRNA水平上調2.7倍,N蛋白表達水平上調1.7倍(圖4 D、E),表明過表達hAPN促進PDCoV復制。
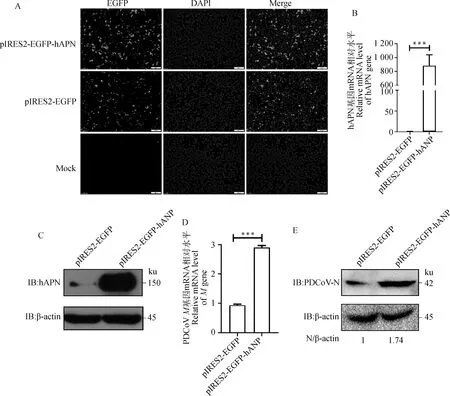
A.熒光顯微鏡觀察綠色熒光蛋白的表達(標尺=100 μm);B.RT-qPCR檢測HEK293細胞轉染pIRES2-EGFP-hAPN后hAPN基因mRNA水平(***.P<0.001);C.Western blot檢測HEK293細胞轉染pIRES2-EGFP-hAPN后hAPN蛋白表達水平;D.RT-qPCR檢測過表達hAPN后M基因mRNA水平(***.P<0.001);E.Western blot檢測過表達hAPN后N蛋白表達水平。掃描文章首頁OSID碼可查看彩圖
2.5 hAPN與PDCoV S蛋白的互作分析
為分析hAPN和PDCoV S蛋白的互作關系,從PDB數據庫獲取hAPN和PDCoV S的三聚體結構(圖5 A、G);PDCoV S蛋白的單體結構及其主要結構域包括C-末端結構域(C-terminal domain,CTD)、N-末端結構域(N-terminal domain,NTD)、融合肽(fusion peptide,FP)、七肽重復序列(heptad repeat,HR)(圖5 B)。HCoV-229E和PDCoV S1 RBD,包含6個β-折疊和3個短而不連續的環組成的受體結合基序(receptor binding motifs,RBM)(圖5 C、D)。序列和結構比對發現PDCoV和HCoV-229E S1 RBD具有相似的空間結構,表明PDCoV RBD與hAPN的結合可能類似HCoV-229E RBD和hAPN的結合(圖5 E、F)。分子對接結果表明,PDCoV S1 RBD能夠結合hAPN結構域Ⅱ,主要是S1蛋白的RBM1氨基酸殘基TYR、THR、THR、PHE和MET通過氫鍵與hAPN的氨基酸殘基PHE、GLN、ARG和SER結合(圖5 G、H)。
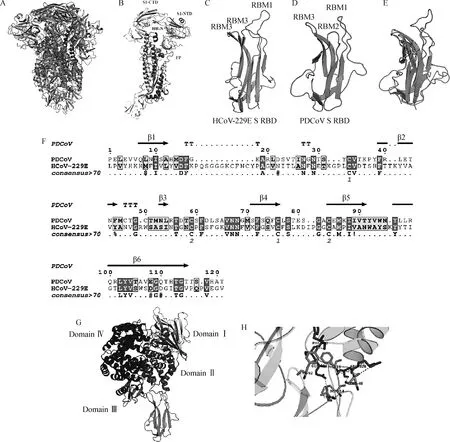
A.PDCoV S 蛋白整體結構;B.PDCoV S 蛋白單體結構;C.HCoV-229E S1 RBD;D.PDCoV S1 RBD;E.HCoV-229E和PDCoV S1 RBD結構比對;F.HCoV-229E和PDCoV S1 RBD序列比對;G~H.分子對接模擬PDCoV S1 RBD和hAPN蛋白的互作。掃描文章首頁OSID碼可查看彩圖
3 討 論
PDCoV是近年來新發現的一種豬冠狀病毒,人工接種PDCoV能夠感染豬、牛、雞和火雞等多種動物。PDCoV感染仔豬后引起明顯的腹瀉癥狀,而感染小雞后腹瀉癥狀較輕,接種PDCoV的小牛甚至未出現腹瀉和其他臨床癥狀。Lednicky等在兒童血清樣品中檢測到變異的PDCoV,首次報道了PDCoV可感染人。鑒于PDCoV在豬群的廣泛流行和跨宿主傳播風險,研究宿主細胞蛋白在PDCoV復制中的作用具有重要意義。過表達hAPN、fAPN、pAPN、cAPN顯著增加細胞對PDCoV的易感性。APN在不同腸段中的差異表達與PDCoV腸道組織嗜性也密切相關。本研究發現敲除hAPN降低PDCoV復制,而過表達hAPN促進PDCoV復制,表明hAPN是影響PDCoV復制的重要宿主細胞因子,豐富了PDCoV跨種傳播和致病機制理論。
S蛋白是冠狀病毒入侵宿主細胞的關鍵蛋白,在病毒感染宿主細胞的過程中,S蛋白構象的變化能促進病毒囊膜與宿主細胞膜的融合。此外,宿主細胞內的酸性環境和蛋白水解酶對S蛋白的活化也是病毒囊膜與宿主細胞膜融合所必需的。胰蛋白酶可通過增強細胞與細胞之間的融合而促進PDCoV增殖,但在病毒入侵細胞的過程中不發揮關鍵作用。此外,PDCoV通過胰蛋白酶介導的細胞膜表面入侵ST和IPI-2I細胞的效率明顯高于內吞體途徑。HEK293細胞是由剪切過的5型腺病毒DNA轉染的人胚腎細胞形成的細胞系,HEK293 T細胞是能夠穩定表達SV40 T抗原的HEK293衍生細胞系。PEDV可以感染HEK293細胞,且與Vero細胞上的增殖特點相似。此外,黃病毒科成員,日本腦炎病毒(Japanese encephalitis virus,JEV)、寨卡病毒(Zika virus,ZIKV)和黃熱病毒(yellow fever virus,YFV)也可感染HEK293T細胞。由于HEK293細胞貼壁能力弱,對胰蛋白酶的耐受程度較弱,本研究在不添加胰蛋白酶的情況下,發現PDCoV可在HEK293細胞至少傳至2代,表明PDCoV可不依賴于胰蛋白酶入侵HEK293細胞。
CRISPR/Cas9系統是新發現的一種基因編輯工具,廣泛應用于研究宿主因子在病毒復制中的作用。近年來,CRISPR/Cas9系統也被應用于篩選調節病毒復制的相關宿主因子。關于APN是否為PDCoV入侵宿主細胞的受體存在爭議。本研究為驗證hAPN在PDCoV毒株CHN-SC2015復制中的作用,通過CRISPR/Cas9技術構建hAPN敲除細胞系,發現hAPN敲除導致PDCoV基因mRNA水平下調約75%,N蛋白表達水平下降約66%。本研究還通過hAPN過表達驗證其對病毒復制的影響,結果發現過表達hAPN可導致PDCoV基因mRNA水平上調2.7倍,N蛋白表達水平上調1.7倍,證實hAPN可明顯促進PDCoV復制。
冠狀病毒S蛋白主要由S1和S2結構域組成,其中S1主要負責識別受體,而S2介導宿主細胞膜與病毒囊膜的融合。有研究表明,冠狀病毒S1 RBD可以與APN結合,介導病毒入侵宿主細胞。HCoV-229E與hAPN的胞外域Ⅱ結合,而TGEV主要與pAPN的胞外域Ⅳ結合。pAPN結構域Ⅶ(581—967 aa)在PEDV復制中發揮重要作用。在PDCoV研究中,Zhu等發現PDCoV S1-CTD蛋白可結合pAPN。本研究通過序列和結構比對發現PDCoV與HCoV-229E S1 RBD具有相似的空間結構,表明PDCoV可能與hAPN結合。因此,本研究通過分子對接方法模擬PDCoV S1 RBD與hAPN的結合,發現PDCoV S1 RBD能夠通過RBM1與hAPN結構域Ⅱ結合。
4 結 論
PDCoV可以感染HEK293細胞,宿主細胞因子hAPN表達可促進PDCoV復制。此外,PDCoV S1 RBD可通過RBM1氨基酸殘基TYR、THR、THR、PHE和MET與hAPN蛋白的氨基酸殘基PHE、GLN、ARG和SER結合,證實了hAPN是影響PDCoV復制的一個重要宿主細胞因子。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
