火炬松體細(xì)胞胚胎發(fā)生體系的優(yōu)化
2022-05-24 06:57:14楊春霞谷振軍肖平江李火根
林業(yè)科學(xué)研究 2022年3期
關(guān)鍵詞:研究
胡 珊,楊春霞,谷振軍,杜 強(qiáng),肖平江,李火根
(1. 江西省林業(yè)科學(xué)院,江西 南昌 330013;2. 南京林業(yè)大學(xué)南方現(xiàn)代林業(yè)協(xié)同創(chuàng)新中心,江蘇 南京 210037;3. 安福縣武功山林場(chǎng),江西 吉安 343200)
火炬松(Pinus taedaL.)作為我國(guó)成功引種的國(guó)外松樹種之一,因其生長(zhǎng)快、用途廣、適應(yīng)性強(qiáng)等特性,成為我國(guó)南方地區(qū)的重要用材樹種和生態(tài)樹種。然而,火炬松育種和生長(zhǎng)周期長(zhǎng),常規(guī)有性繁殖子代表型變異大,且種子園建設(shè)成本和維護(hù)成本高、更新慢,在短期內(nèi)不能滿足生產(chǎn)上對(duì)火炬松苗木的需求,限制了火炬松規(guī)模化推廣應(yīng)用。無(wú)性快繁不僅可在短期內(nèi)保質(zhì)保量地規(guī)模生產(chǎn)無(wú)性系苗木,而且還可縮短育種周期,加速育種進(jìn)程。
植 物 體 細(xì) 胞 胚 胎 發(fā) 生 ( Somatic embryogenesis)具有繁殖系數(shù)高、遺傳穩(wěn)定性好、生產(chǎn)周期短、便于規(guī)模化生產(chǎn)等優(yōu)點(diǎn),是植物良種規(guī)模化繁殖的有效手段之一,在提高森林生產(chǎn)力、可持續(xù)性和林產(chǎn)品一致性方面發(fā)揮重要作用[1]。早在1987 年,美國(guó)學(xué)者[2]開始火炬松體胚發(fā)生技術(shù)研究,初步建立了懸浮培養(yǎng)體系并獲得再生植株。隨著Becwar 等[3]、Li 等[4]、Pullman[5]等學(xué)者對(duì)體胚發(fā)生技術(shù)研究的深入,體胚技術(shù)逐漸成熟,火炬松一些優(yōu)良基因型的體胚誘導(dǎo)率和植株再生率顯著提升,并成功用于商業(yè)化生產(chǎn)[6]。國(guó)內(nèi)對(duì)火炬松體胚發(fā)生方面的研究相對(duì)較少,主要集中在20 世紀(jì)90 年代[7],雖然近年來(lái)也開展過(guò)一些研究,但效果不理想[8-9]。無(wú)論是國(guó)外還是國(guó)內(nèi),火炬松體胚發(fā)生技術(shù)研究中仍存在一些共性的、難以克服的技術(shù)問(wèn)題,如某些優(yōu)良基因型的胚性愈傷組織誘導(dǎo)率低、增殖穩(wěn)定性差、體胚成熟分化同步性低及體胚萌發(fā)率低等[6],造成大多數(shù)火炬松優(yōu)良基因型目前尚不能通過(guò)體胚發(fā)生途徑進(jìn)行規(guī)模化生產(chǎn)。
為此,本研究以江西省推廣應(yīng)用的火炬松良種[贛R-CS0(1)-PT-001-2020(5)]的未成熟種子為材料,對(duì)胚性愈傷組織誘導(dǎo)與增殖、體胚成熟與萌發(fā)、體胚苗移栽等相關(guān)技術(shù)進(jìn)行了優(yōu)化,以期提高火炬松胚性愈傷組織誘導(dǎo)率、體胚成熟分化及植株再生效率,為江西省火炬松良種體胚繁育奠定了基礎(chǔ),并為其規(guī)模化生產(chǎn)體胚苗提供技術(shù)支撐。
1 試驗(yàn)材料
未成熟球果采自江西省安福縣武功山林場(chǎng)的第1 代火炬松無(wú)性系種子園中6 個(gè)長(zhǎng)勢(shì)好、結(jié)實(shí)量高、且子代速生特性明顯的無(wú)性系(武林1、武林2、武林3、武林16、榮山52、湖南39)。自2019年7 月7 日開始采集,8 月19 日結(jié)束,每周采集1 次,共采集7 次,每株每次采集20~30 個(gè)未成熟球果,隨機(jī)取3 個(gè)球果鏡檢,剩余的放置4 ℃冰箱保存?zhèn)溆谩?/p>
2 方法
2.1 外植體消毒與接種
未成熟種子先用無(wú)菌水清洗1 遍,然后用75%(v/v)酒精消毒30 s,無(wú)菌水清洗3 遍,再用0.1%(w/v)升汞消毒7~8 min,無(wú)菌水清洗5 遍,最后用無(wú)菌濾紙將種子表面多余的水分吸干,備用。
消毒后的種子用無(wú)菌手術(shù)刀和鑷子剝除種子的外種皮和內(nèi)種皮,將其接種至提前配好的胚性愈傷組織誘導(dǎo)培養(yǎng)基中,每皿10 粒。
2.2 胚性愈傷組織誘導(dǎo)與增殖
設(shè)置不同基因型、基本培養(yǎng)基類型、不同采樣時(shí)間、PGR 不同組合(NAA、6-BA 和KT)及濃度梯度等因素對(duì)火炬松胚性愈傷組織誘導(dǎo)影響的試驗(yàn)。
不同采樣時(shí)期、不同基因型的誘導(dǎo)培養(yǎng)基采用DCR 培養(yǎng)基 + 1.0 mg·L?12,4-D + 0.5 mg·L?16-BA + 1.0 mg·L?1NAA + 5.0 mg·L?1ABA;不同基本培養(yǎng)基類型采用基本培養(yǎng)基 + 1.0 mg·L?12,4-D +0.5 mg·L?16-BA + 1.0 mg·L?1NAA + 5.0 mg·L?1ABA;PGR 組合則采用4 因素3 水平正交試驗(yàn)設(shè)計(jì),在DCR 基本培養(yǎng)基上添加不同PGR 及其濃度的組合。以上培養(yǎng)基同時(shí)添加L-谷氨酰胺450 mg·L?1、肌醇500 mg·L?1、水解酪蛋白500 mg·L?1、2-嗎啉乙磺酸250 mg·L?1、蔗糖30 g·L?1、卡拉膠7.5 g·L?1。每個(gè)處理設(shè)置3 個(gè)重復(fù),(22±1)℃暗培養(yǎng), 8 周后統(tǒng)計(jì)胚性愈傷組織誘導(dǎo)率。
選擇生長(zhǎng)狀態(tài)良好的胚性愈傷組織,在增殖培養(yǎng)基中(MLP+2.0 mg·L?1NAA+0.5 mg·L?16-BA+0.5 mg·L?1KT+450 mg·L?1L-谷氨酰胺+500 mg·L?1肌醇+500 mg·L?1水解酪蛋白+30 g·L?1麥芽糖+7.5 g·L?1卡拉膠)進(jìn)行繼代,培養(yǎng)條件同誘導(dǎo)培養(yǎng)。
2.3 成熟分化培養(yǎng)
選擇增殖能力強(qiáng)、具有典型的胚性胚柄細(xì)胞團(tuán)結(jié)構(gòu)細(xì)胞系#R52(榮山52 無(wú)性系的未成熟種子誘導(dǎo)的一個(gè)細(xì)胞系),在增殖培養(yǎng)基上增殖10 d 后,稱取鮮質(zhì)量1 g,轉(zhuǎn)接至MLP 基本培養(yǎng)基,暗培養(yǎng)14 d 后再轉(zhuǎn)移至成熟分化培養(yǎng)基中進(jìn)行培養(yǎng),成熟培養(yǎng)基設(shè)置ABA 濃度(6、8、10 mg·L?1)、PEG 8 000 濃度(120、140、160 g·L?1)、麥芽糖濃度(20、30、40 g·L?1)3 因素3 水平的正交設(shè)計(jì)試驗(yàn),同時(shí)添加450 mg·L?1L-谷氨酰胺 、500 mg·L?1肌 醇、 500 mg·L?1水 解 酪 蛋 白、 250 mg·L?12-嗎啉乙磺酸、30 g·L?1麥芽糖、7.5 g·L?1卡拉膠,8 周后統(tǒng)計(jì)成熟胚的數(shù)量。
2.4 萌發(fā)與移栽
將細(xì)胞系#R52 成熟的體細(xì)胞胚胎接種于萌發(fā)培養(yǎng)基G0 和G1(G0=WPM + 肌醇500 mg·L?1+L-谷氨酰胺450 mg·L?1+ 水解酪蛋白 500 mg·L?1+2-嗎啉乙磺酸 250 mg·L?1+ 麥芽糖30.0 g·L?1+ 結(jié)冷膠10 g·L?1,G1=G0 + 3.5 g·L?1活性炭)中進(jìn)行萌發(fā),先暗培養(yǎng)7 d,然后置光照下培養(yǎng)。組織培養(yǎng)間的環(huán)境控制為:光照強(qiáng)度為50 μmol·m?2·s?2、光照16 h/黑暗8 h,溫度(22±1)℃,濕度為60%~65%。將體胚生根且長(zhǎng)出真葉判定為萌發(fā),統(tǒng)計(jì)8 周內(nèi)的萌發(fā)率。
體胚萌發(fā)后,將其放在瓶裝的萌發(fā)培養(yǎng)基上繼續(xù)培養(yǎng)4~6 周,室溫下開口煉苗1 周后移栽,移栽基質(zhì)按照泥炭土∶珍珠巖=3∶1 的比例配置,光照16 h 的光周期/黑暗8 h、光照強(qiáng)度為50 μmol·m?2·s?2,溫度25 ℃,濕度前兩周控制為85%,后續(xù)兩周濕度控制75%。8 周后統(tǒng)計(jì)移栽成活率。
2.5 數(shù)據(jù)統(tǒng)計(jì)分析
使用Excel 進(jìn)行數(shù)據(jù)的計(jì)算,采用SPSS 22.0軟件對(duì)數(shù)據(jù)進(jìn)行方差分析和多重比較。采用單因素的一般線性模型進(jìn)行方差分析,模型為:
Y=u+F+e
式中:u為平均值,F(xiàn)為處理,e為誤差。
對(duì)各處理的試驗(yàn)結(jié)果平均值進(jìn)行Duncan 多重比較。
胚性愈傷組織誘導(dǎo)率=誘導(dǎo)出胚性愈傷組織的未成熟合子胚數(shù)/接種的未成熟合子胚數(shù) × 100%。
萌發(fā)率=已萌發(fā)的體胚數(shù)量/萌發(fā)試驗(yàn)體胚起始數(shù)量 × 100%。
移栽成活率=成活的體胚苗數(shù)量/移栽的體胚苗總數(shù)量 × 100%。
3 結(jié)果與分析
3.1 火炬松胚性愈傷組織誘導(dǎo)
3.1.1 采集時(shí)期 將火炬松3 個(gè)無(wú)性系不同時(shí)期采集的未成熟種子(圖1a)鏡檢后對(duì)應(yīng)合子胚發(fā)育期,剝除內(nèi)外種皮后(圖1b)接種于誘導(dǎo)培養(yǎng)基中,胚性愈傷組織(圖1c)誘導(dǎo)率區(qū)別很大,且均以7 月20 日采集的未成熟種子的胚性愈傷組織誘導(dǎo)率最高(表1)。由此可推斷,火炬松最佳采樣時(shí)期是7 月20 日,最佳的合子胚發(fā)育期處于裂生多胚期。

注:a. 火炬松球果及其種子;b. 去除種皮的未成熟合子胚;c. 誘導(dǎo)胚性愈傷組織;d. 繼代一次的胚性愈傷組織;e. 體細(xì)胞胚胎的成熟分化培養(yǎng);f. 成熟的體細(xì)胞胚胎;g. 萌發(fā)試驗(yàn);h. 壯苗培養(yǎng);i. 煉苗移栽。Notes: a. Cones and seeds of loblolly pine; b. Immature zygotic embryo with seed coat removed; c. Mature differentiation of embryogenic callus; d. Embryogenic callus subcultured once; e.Maturation culture of T somatic embryos; f. Mature somatic embryos;g. Germination test; h. Strong seedling culture; i. Seedling refining and transplanting.
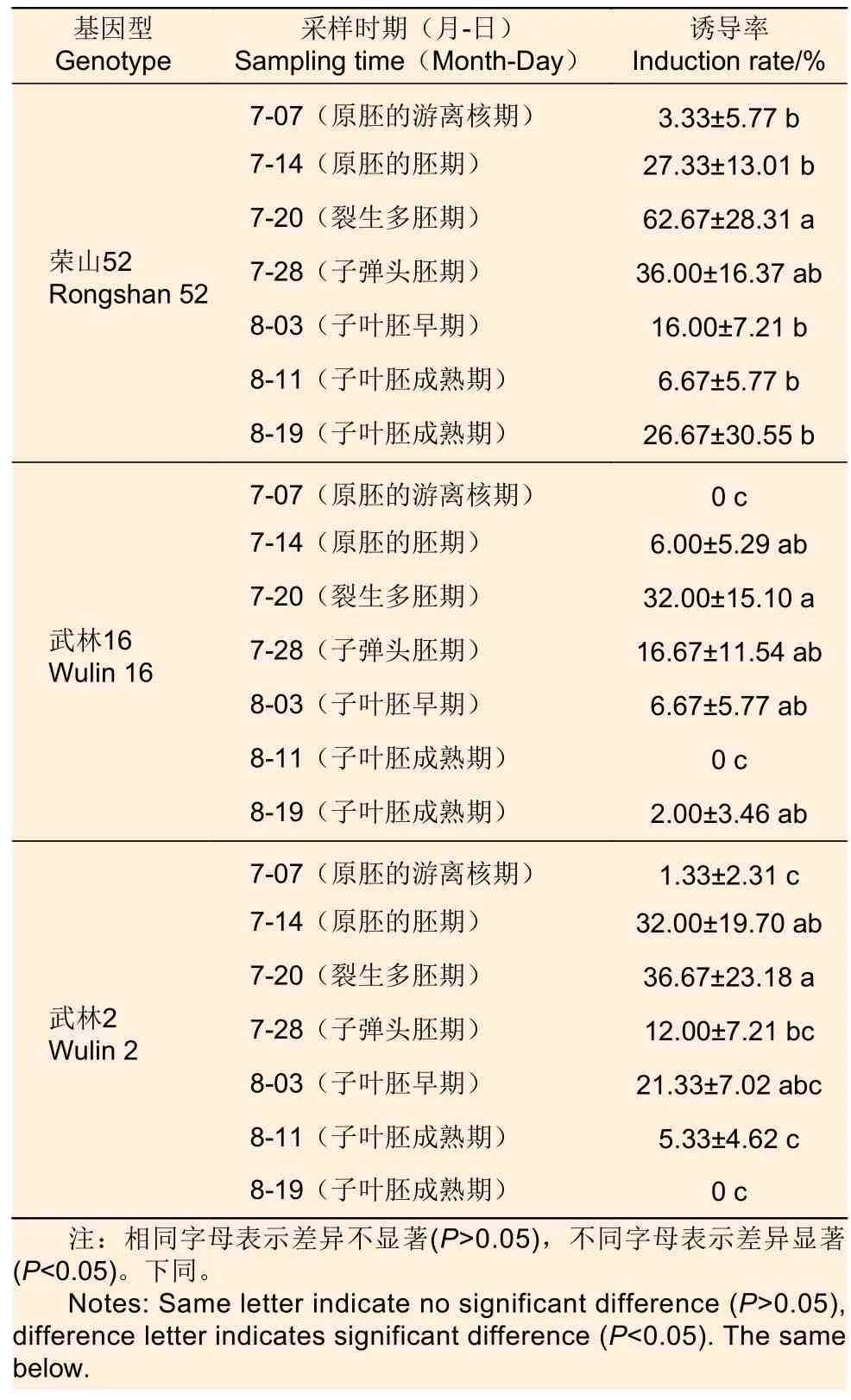
表1 不同基因型未成熟合子胚在不同采樣時(shí)期的胚性愈傷組織誘導(dǎo)率Table 1 Effect of different genotypes of immature zygotic embryos on embryogenic callus induction rate at different sampling stages
3.1.2 基因型 將7 月20 日采集的6 個(gè)不同基因型火炬松未成熟種子接種于誘導(dǎo)培養(yǎng)基中進(jìn)行胚性愈傷組織誘導(dǎo),發(fā)現(xiàn)胚性愈傷組織誘導(dǎo)率差別明顯(表2),其中,榮山52 號(hào)未成熟合子胚的胚性愈傷組織的誘導(dǎo)率最高,達(dá)63.33%;湖南39 號(hào)次之,愈傷組織誘導(dǎo)率達(dá)60.00%;武林1 號(hào)胚性愈傷組織的誘導(dǎo)率最低,僅為10%。
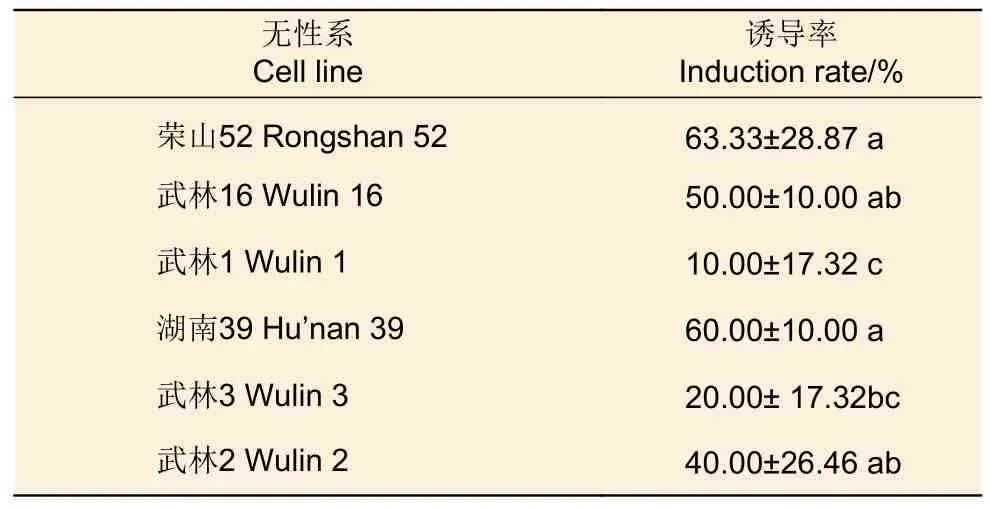
表2 不同基因型對(duì)胚性愈傷組織的影響Table 2 Effects of different genotype on embryonic callus introduction
3.1.3 基本培養(yǎng)基類型 以武林2 號(hào)未成熟合子胚(7 月20 日)為材料開展不同基本培養(yǎng)基上胚性愈傷組織誘導(dǎo)試驗(yàn)(表3),發(fā)現(xiàn)DCR 培養(yǎng)基上胚性愈傷組織的誘導(dǎo)率最高,達(dá)到40.00%,且與其他培養(yǎng)基間差異顯著;其次WPM 和MLP 培養(yǎng)基,胚性愈傷組織誘導(dǎo)率分別為20.00%和13.33%;而MLV、WV5 和B5 培養(yǎng)基中誘導(dǎo)率較低。
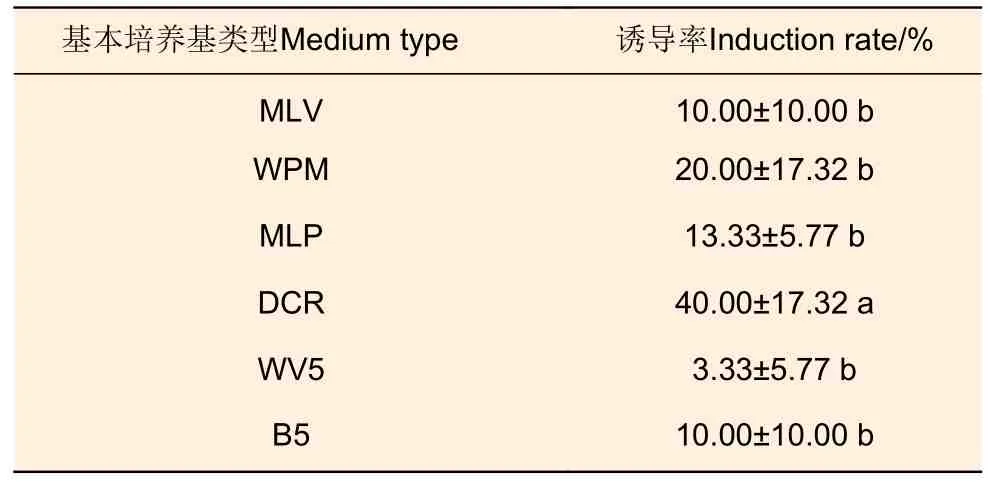
表3 不同基本培養(yǎng)基對(duì)胚性愈傷組織誘導(dǎo)的影響Table 3 Effects of different basic media on embryogenic callus induction
3.1.4 不同PGR 組合及其濃度 榮山52 的未成熟合子胚接種于不同濃度的6-BA、2,4-D、NAA和ABA 的正交設(shè)計(jì)處理的9 種培養(yǎng)基中,培養(yǎng)基配方為DCR + PGR 組合,結(jié)果(表4)發(fā)現(xiàn):處理3的胚性愈傷組織誘導(dǎo)率最高,達(dá)50.00%;其次是處理7,胚性愈傷組織誘導(dǎo)率達(dá)18.33%;處理6 的胚性愈傷組織的誘導(dǎo)率最低。從正交設(shè)計(jì)的方差分析可得,6-BA 濃度、NAA 濃度以及ABA 濃度都對(duì)胚性愈傷組織的誘導(dǎo)率有顯著影響。
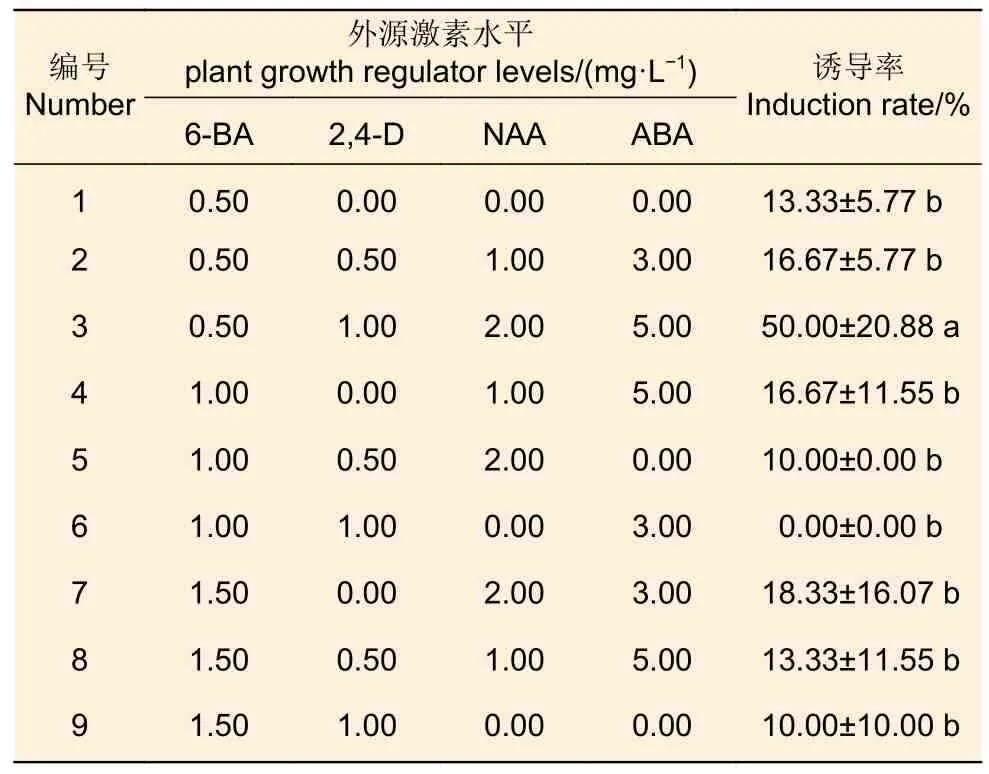
表4 不同PGR 組合及濃度對(duì)榮山52 胚性愈傷組織誘導(dǎo)率的影響Table 4 Effects of different PGR combinations and concentrations on embryogenic callus induction rate of Rongshan 52
3.2 體胚成熟分化培養(yǎng)
選擇半透明、粘連狀,且具有典型ESM 結(jié)構(gòu)的#R52 胚性愈傷組織(圖1d)進(jìn)行成熟分化試驗(yàn)。#R52 胚性愈傷組織在成熟分化培養(yǎng)基培養(yǎng)1 周后,胚性愈傷組織表面即可觀察到原胚,此后原胚不斷生長(zhǎng)分化,子葉開始形成,2 個(gè)月左右體細(xì)胞胚胎基本發(fā)育成熟(圖1e、f)。體胚成熟分化培養(yǎng)的9 種處理中體胚誘導(dǎo)率差別明顯(表5、圖2),處理5 對(duì)#R52 胚性愈傷組織成熟誘導(dǎo)效果最好,平均可達(dá)411 個(gè)·g?1成熟胚,最高可達(dá)448 個(gè)·g?1成熟胚;其次是處理3,成熟胚數(shù)量平均達(dá)257 個(gè)·g?1,最高可達(dá)327 個(gè)·g?1。

注: a. 處理1 的體胚;b. 處理2 的體胚;c. 處理3 的體胚;d. 處理4 的體胚;e. 處理5 的體胚;f. 處理6 的體胚;g. 處理7 的體胚;h. 處理8 的體胚;i. 處理9 的體胚。Notes: a. the somatic embryos treated with 1; b. the somatic embryos treated with 2; c. the somatic embryos treated with 3; d. the somatic embryos treated with 4; e. the somatic embryos treated with 5; f. the somatic embryos treated with 6; g. the somatic embryos treated with 7; h. the somatic embryos treated with 8; i. the somatic embryos treated with 9.
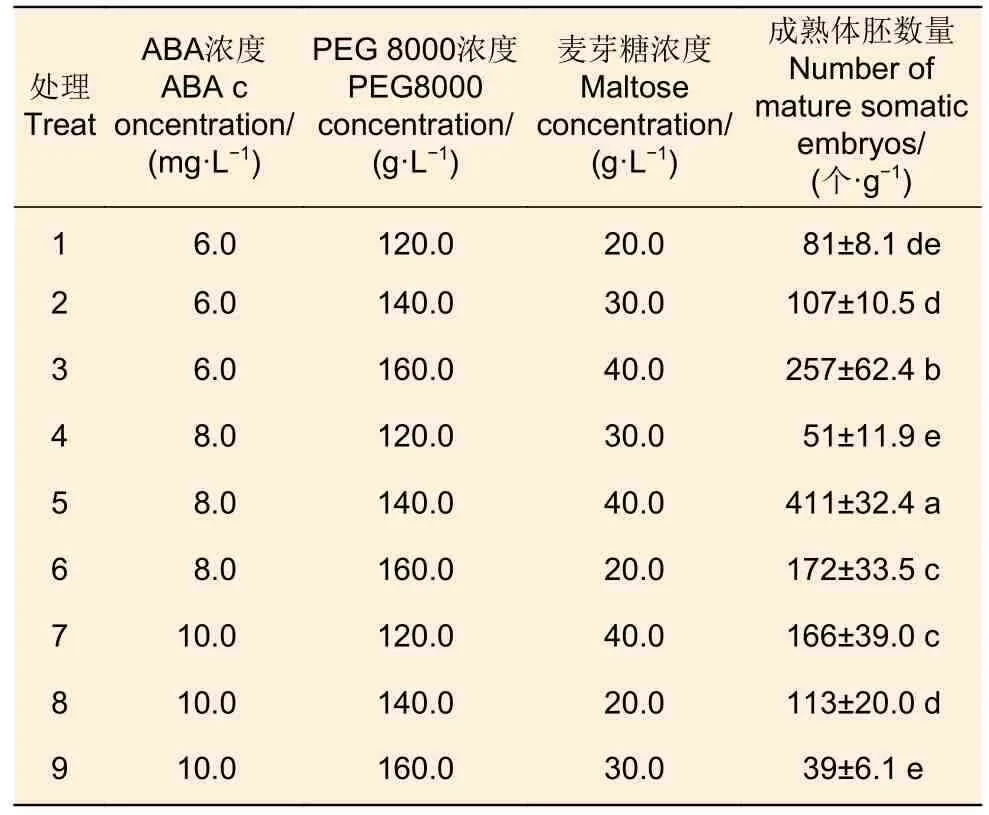
表5 培養(yǎng)基組合對(duì)#R52 體胚成熟分化培養(yǎng)的影響Table 5 Effect of medium combination on somatic embryo mature differentiation of #R52
對(duì)成熟分化培養(yǎng)的結(jié)果進(jìn)行方差分析,可得ABA 濃度、PEG 8 000 濃度以及麥芽糖濃度均對(duì)成熟分化培養(yǎng)的影響很大,且3 個(gè)因素影響效果的主次為:麥芽糖>PEG 8 000>ABA。
3.3 萌發(fā)與煉苗移栽
將火炬松成熟胚(圖1f)置于萌發(fā)培養(yǎng)基中萌發(fā),#R52 細(xì)胞系在添加3.5 g·L?1活性炭的G1 培養(yǎng)基中體胚萌發(fā)率可達(dá)93.33%(表6),優(yōu)于不添加活性炭的G0 培養(yǎng)基。體胚萌發(fā)(圖1g)后,在瓶裝基本培養(yǎng)基中繼續(xù)培養(yǎng)(圖1h)4~6 周后,選擇生長(zhǎng)旺盛、根系發(fā)達(dá)的的體胚苗移栽至基質(zhì)中,做好控溫控濕管理,移栽成活率可達(dá)86.30%(表6、圖1i)。

表6 #R52 體胚的萌發(fā)與移栽Table 6 Germination and transplanting on somatic embryos of #R52
4 討論
4.1 胚性愈傷組織誘導(dǎo)
基因型是影響體胚誘導(dǎo)的重要因素之一,已有研究表明,在體胚發(fā)生起始時(shí)存在很強(qiáng)的遺傳累積效應(yīng)和母體效應(yīng),故在取材時(shí)選擇優(yōu)良基因型對(duì)體胚成功誘導(dǎo)至關(guān)重要[10-11]。不同基因型之間胚性愈傷組織誘導(dǎo)率存在顯著差異,Tang 等[12]和MacKay等[13]對(duì)火炬松不同家系在相同培養(yǎng)條件下進(jìn)行胚性愈傷組織誘導(dǎo),均得出相似的結(jié)論。Pullman等[5]對(duì)10 個(gè)開放授粉的火炬松無(wú)性系進(jìn)行了體胚發(fā)生研究,胚性愈傷組織誘導(dǎo)率最高的無(wú)性系可達(dá)33%,部分無(wú)性系誘導(dǎo)困難,不能誘導(dǎo)出胚性愈傷組織。本研究中,火炬松6 個(gè)基因型在誘導(dǎo)胚性愈傷組織效率上也存在明顯差異,其中,榮山52 和湖南39 兩個(gè)無(wú)性系種子的誘導(dǎo)率均超過(guò)50%,榮山52 的無(wú)性系種子的誘導(dǎo)率高達(dá)63.33%,高于以往的報(bào)道[8-13]。在其他針葉樹種的研究中,如馬尾松(Pinus massonianaLamb.)[14]和云南松(Pinus yunnanensisFranch.)[15],不同基因型之間胚性愈傷組織誘導(dǎo)率也呈顯著差異,說(shuō)明基因型是影響胚性愈傷組織誘導(dǎo)率的關(guān)鍵因子。
研究發(fā)現(xiàn),利用未成熟合子胚為材料更有利于胚性愈傷組織誘導(dǎo)[11,16-17],且大多研究表明,當(dāng)合子胚發(fā)育處于裂生多胚期時(shí)其胚性愈傷組織誘導(dǎo)率顯著高于其他發(fā)育階段[18-19]。國(guó)內(nèi)外對(duì)火炬松球果采樣期發(fā)現(xiàn),其最佳采樣期在7 月中下旬,該時(shí)期合子胚發(fā)育階段大多處于裂生多胚期[3,8],此時(shí)胚性愈傷組織誘導(dǎo)率最高。本研究結(jié)果與以往研究結(jié)果一致。
培養(yǎng)基成分是誘導(dǎo)體胚成功的關(guān)鍵因子之一,包括基本培養(yǎng)基、PGR 及碳源。基本培養(yǎng)基中某些成分(尤其是氮源和一些金屬離子)的增加或減少能有效促進(jìn)胚性愈傷組織誘導(dǎo)與生長(zhǎng)[20],Pullman等[21]發(fā)現(xiàn)火炬松未成熟種子在改良的1/2 P6 基本培養(yǎng)基上胚性愈傷組織誘導(dǎo)率最高,隨后,研究表明在基本培養(yǎng)基1/2 P6 基礎(chǔ)上添加一定濃度的MES、生物素和葉酸,可以顯著提高胚性愈傷組織的誘導(dǎo)率和生長(zhǎng)速度[18]。本研究發(fā)現(xiàn),DCR 培養(yǎng)基是誘導(dǎo)胚性愈傷組織的最佳基本培養(yǎng)基,DCR 培養(yǎng)基中NH4+的含量介于MLP 與MLV 培養(yǎng)基之間,NO3?的含量低于MLP 與MLV,DCR 培養(yǎng)基中NH4+:NO3?約為0.38,MLP 約為0.17,MLV 約為0.44,這說(shuō)明適當(dāng)降低氮含量以及調(diào)節(jié)NH4+與NO3?的比例是誘導(dǎo)胚性愈傷組織的關(guān)鍵因素。在增殖培養(yǎng)與成熟分化培養(yǎng)中,本研究發(fā)現(xiàn),火炬松胚性愈傷組織適合在MLP 中培養(yǎng),這可能同樣和氮素的含量與比例有關(guān)。在Pullman 等[5]、Tang 等[12]的研究中也有相似的發(fā)現(xiàn)。PGR 是培養(yǎng)基中誘導(dǎo)體胚的決定性因素,火炬松胚性愈傷組織誘導(dǎo)中,使用最多的PGR 組合是2,4-D + 6-BA,2,4-D + 6-BA +KT,NAA + 6-BA + KT 等[8,22-23],且有研究表明向培養(yǎng)基中添加 ABA,火炬松胚性愈傷組織誘導(dǎo)作用增強(qiáng)[5,24]。Pullman 等[24]的研究中,在添加1.1 mg·L?12,4-D + 0.45 mg·L?16-BA + 0.43 mg·L?1KT的培養(yǎng)基中,火炬松誘導(dǎo)率最高可達(dá)30.1%。添加1 mg·L?1ABA 可以顯著提高誘導(dǎo)率,32 個(gè)火炬松半同胞家系的平均誘導(dǎo)率可達(dá)17.90%[25]。本研究利用4 種PGR(2,4-D、6-BA、NAA 和ABA)組合顯著提升了火炬松胚性愈傷組織誘導(dǎo)效率,誘導(dǎo)率最高可達(dá)50%,明顯優(yōu)于以往的研究結(jié)果。4 種PGR 中6-BA、NAA、ABA 對(duì)胚性愈傷組織誘導(dǎo)的影響是顯著的,說(shuō)明這3 種PGR 在火炬松胚性愈傷組織誘導(dǎo)中發(fā)揮著重要作用,而高濃度2,4-D 會(huì)導(dǎo)致胚性愈傷組織過(guò)甲基化,抑制體胚的成熟,所以在后續(xù)的培養(yǎng)中應(yīng)及時(shí)調(diào)整,降低2,4-D 的濃度或用NAA 替代。因此,作者認(rèn)為胚性愈傷組織的誘導(dǎo)率的提高更多的是多種PGR 的聯(lián)合效應(yīng),尤其是6-BA、NAA、ABA,若要進(jìn)一步提高胚性愈傷組織的誘導(dǎo)率,可在這3 種PGR 濃度的基礎(chǔ)上深入開展試驗(yàn),但3 種PGR 對(duì)胚性愈傷組織誘導(dǎo)的聯(lián)合效應(yīng)的機(jī)制尚不明確,還需要更加深入的研究。
4.2 體胚成熟分化培養(yǎng)
體胚成熟分化培養(yǎng)是體胚發(fā)生技術(shù)研究中的難點(diǎn)之一,也是限制體胚技術(shù)商業(yè)化生產(chǎn)應(yīng)用的重要因素,在體胚誘導(dǎo)過(guò)程中,體細(xì)胞胚畸形或發(fā)育不完全嚴(yán)重影響后期體胚的萌發(fā)率[26]。眾多研究表明,ABA、PEG 8 000 以及麥芽糖在針葉樹種體胚成熟分化培養(yǎng)中發(fā)揮著重要作用[13,18,27-29]。
適當(dāng)濃度的ABA 促進(jìn)體細(xì)胞胚胎的成熟[30],針葉樹種中ABA 濃度對(duì)體胚成熟分化培養(yǎng)的作用效果差異顯著。馬尾松[17]在成熟培養(yǎng)基中添加2 mg·L?1ABA,對(duì)細(xì)胞系體胚的成熟效果較好。20 mg·L?1是紅皮云杉(Picea koraiensisNakai)細(xì)胞系體胚成熟的ABA 最佳濃度,能獲得較高的成熟體胚數(shù)量[31],并且畸形率較低。在對(duì)火炬松體胚的研究中,眾多學(xué)者也對(duì)ABA 在其體胚成熟的作用展開了研究。Kapik 等[32]對(duì)火炬松合子胚的發(fā)育過(guò)程的ABA 估算研究時(shí),發(fā)現(xiàn)火炬松合子胚ABA 濃度的峰值出現(xiàn)在合子胚胚胎發(fā)育的早期和晚期。唐巍[33]在對(duì)火炬松懸浮細(xì)胞進(jìn)行成熟培養(yǎng)過(guò)程中,添加5 mg·L?1ABA,能夠觀察到裂生多胚以及子葉原基的分化。Pullman 等[6]在火炬松的體胚成熟分化培養(yǎng)的研究中認(rèn)為,ABA 濃度為9.0 mg·L?1時(shí),獲得的成熟體胚的數(shù)量較多;而本研究中,火炬松體胚成熟分化培養(yǎng)的最適ABA 濃度為8.0 mg·L?1,稍低于Pullman 的研究結(jié)果,說(shuō)明火炬松不同基因型甚至不同細(xì)胞系之間體胚成熟分化培養(yǎng)所需ABA 濃度可能略有差異。
PEG 在培養(yǎng)基中作為滲透劑的作用可能與誘導(dǎo)水分脅迫、脫水耐受性和一些貯藏儲(chǔ)備的積累有關(guān),在針葉樹種的體細(xì)胞胚胎發(fā)生研究中也常被用于促進(jìn)體胚成熟[34-35]。在日本黑松(Pinus thunbergiiParl.)[34]體胚成熟培養(yǎng)的培養(yǎng)基中添加0~150 g·L?1PEG 8 000,3~4 周后可見子葉胚,培養(yǎng)8 周左右體胚基本成熟。李亦軒等[36]在對(duì)油松(Pinus tabuliformisCarr.)體胚的成熟培養(yǎng)中亦發(fā)現(xiàn),成熟胚的數(shù)量隨著PEG4000 濃度(25~100 g·L?1)的增加而增加。Pullman 等[6]的研究中,在成熟分化培養(yǎng)基中添加130 g·L?1PEG 8000,火炬松體胚的成熟度較好,可獲得成熟胚150 個(gè)·g?1。本研究在添加140 g·L?1PEG 8000 的成熟分化培養(yǎng)基中,獲得411 個(gè)·g?1成熟胚,成熟分化培養(yǎng)效率得到顯著提升。PEG 對(duì)體細(xì)胞胚胎發(fā)生的作用,不僅體現(xiàn)在PEG 種類及濃度的差異上,更多的是PEG 和培養(yǎng)基的其他添加劑對(duì)體胚發(fā)生產(chǎn)生的組合效應(yīng),這也表明體胚成熟過(guò)程的生理生化反應(yīng)及其調(diào)控機(jī)制是極其復(fù)雜的。
糖類物質(zhì)是體胚發(fā)生過(guò)程中必不可少的添加劑之一,不僅給體細(xì)胞胚胎生存和發(fā)育提供營(yíng)養(yǎng),還能調(diào)控培養(yǎng)基的滲透勢(shì)[37]。針葉樹種體胚成熟培養(yǎng)中常用的糖類多為麥芽糖和蔗糖,糖的濃度也對(duì)體胚成熟培養(yǎng)有顯著影響。Xia 等[17]發(fā)現(xiàn),馬尾松使用麥芽糖進(jìn)行體胚成熟培養(yǎng)效果較好,且增加麥芽糖的濃度提高了產(chǎn)生體細(xì)胞胚胎的能力,但在高濃度(30.0~40.0 g·L?1)之間沒有顯著差異。Peng 等[38]則認(rèn)為,蔗糖有利于紅松(Pinus koraiensisSieb. et Zucc.)體胚成熟培養(yǎng),并且蔗糖濃度對(duì)紅松體胚的成熟有極顯著影響,68.5 g·L?1蔗糖可獲得成熟胚134 個(gè)·g?1。在火炬松中,麥芽糖比蔗糖更能促進(jìn)火炬松體胚的成熟[6-8],吳麗君[39]在添加蔗糖的成熟培養(yǎng)基中未獲得成熟胚,Pullman 等[6]通過(guò)添加60 g·L?1麥芽糖,能獲得150 個(gè)·g?1成熟胚。本研究發(fā)現(xiàn),用40.0 g·L?1麥芽糖效果最佳,且體胚成熟效率較以往研究有顯著提升。
在對(duì)火炬松體胚的研究中發(fā)現(xiàn),ABA、PEG 8000 以及麥芽糖的濃度均對(duì)其體胚成熟分化培養(yǎng)具有極顯著影響,其中,麥芽糖濃度的影響最顯著。ABA、PEG 和麥芽糖對(duì)火炬松體胚成熟分化的影響都不是獨(dú)立產(chǎn)生作用,也不是簡(jiǎn)單的累加效應(yīng),而是三者聯(lián)合時(shí)共同對(duì)火炬松體胚成熟分化產(chǎn)生的促進(jìn)作用。
5 結(jié)論
通過(guò)對(duì)火炬松體細(xì)胞胚胎發(fā)生各培養(yǎng)環(huán)節(jié)進(jìn)行優(yōu)化研究,總結(jié)出一套技術(shù)較為成熟的火炬松體細(xì)胞胚胎發(fā)生技術(shù)體系。火炬松胚性愈傷組織誘導(dǎo)的最佳條件為:以處于裂生多胚期(約7 月20 日左右)的榮山52 無(wú)性系的合子胚為最佳起始材料,其胚性愈傷組織誘導(dǎo)率可達(dá)63.33%;誘導(dǎo)培養(yǎng)最適PGR 組合是0.5 mg·L?16-BA+ 1.0 mg·L?12,4-D+2.0 mg·L?1NAA+ 5.0 mg·L?1ABA;在成熟分化培養(yǎng)基MLP + 8.0 mg·L?1ABA + 140.0 g·L?1PEG 8000 +40.0 g·L?1麥芽糖 + 7.5 g·L?1卡拉膠中,可獲得411個(gè)·g?1成熟胚;在添加活性炭的WPM 培養(yǎng)基上,體胚萌發(fā)率可達(dá)93.33%。
致謝:感謝美國(guó)艾博金林木種苗公司(ArborGen Inc.)和南京林業(yè)大學(xué)陳金慧教授在研究中給予的幫助和指導(dǎo)。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國(guó)際商務(wù)財(cái)會(huì)(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
