豆大薊馬鈉離子通道基因克隆及分析
2022-05-24 06:49:36潘雪蓮袁琳琳陳龍威吳少英
熱帶生物學報 2022年3期
潘雪蓮,楊 磊,袁琳琳,陳龍威,吳少英
(1.海南大學 三亞南繁研究院,海南 三亞 572024; 2.海南大學 植物保護學院,海口 570228;3.崖州灣科技城,海南 三亞 572024)
海南島地處熱帶,屬熱帶季風氣候,除中部山區外,年均氣溫為23~26 ℃,是天然溫室。同時,海南大部分地區年降雨量都超過1 700 mm,良好的水熱條件,非常適合各類農作物的種植[1]。海南是我國最早實現“南菜北運”的省份,豇豆(Vigna unguiculata)就是其中尤為重要的一種[2?3],作為海南農民增產提收最重要的蔬菜品種,從2009—2020年,豇豆的種植面積不斷增加,并達到了2.25萬hm2,總產量為58萬t,約占海南省蔬菜總產量的1/10[4?5]。但隨著豇豆種植面積的逐年增加,病蟲害的發生也逐年加重。豆大薊馬(Megalurothrips usitatus)是豆科作物的一種毀滅性害蟲,嚴重危害豇豆品質和產量[6?7]。該蟲又名豆花薊馬或普通大薊馬,屬纓翅目薊馬科,廣泛分布于熱帶和亞熱帶地區,其寄主植物主要為豆科類作物[6]。豆大薊馬生長周期短、繁殖力強、隱匿性強,世代重疊嚴重,因此,極易在短時間內大面積爆發[8]。豆大薊馬為銼吸式口器,其在取食過程刺穿植物組織并吸取植物汁液,造成豇豆葉片皺縮和畸形,嚴重時會造成生長點萎縮,生長緩慢甚至停止,莢果受害果實表面會變得粗糙或者瘢痕,降低豇豆的果實質量[9?10]。此外,該蟲還可以傳播植物病毒病,從而對豇豆造成間接危害。目前,其防治仍然以化學防治為主[11]。擬除蟲菊酯為一類作用于昆蟲鈉離子通道(Nav)的廣譜性殺蟲劑,具有易降解等優點,因此在田間已經大面積應用。其通過延長Nav開放時間使動作電位升高,導致神經細胞重復放電,從而實現殺蟲的目的[12]。近年來,隨著田間用藥量的成倍增加,豆大薊馬對擬除蟲菊酯類藥劑的抗藥性不斷增強[12]。其產生抗藥性的主要原因之一是Nav發生抗性突變,從而影響藥劑與Nav的結合能力,導致敏感性降低進而產生抗藥性[13?14]。昆蟲Nav由α亞基和β亞基組成,α亞基由4個同源結構域組成,每個同源結構域含有6個跨膜片段,其中,S1~S4稱為電壓傳感模塊,而S5和S6區域稱為P區,β亞基輔助α亞基調控 Nav 表達[14?16]。
本研究組前期的研究表明,海南省豆大薊馬田間種群對擬除蟲菊酯類殺蟲劑已經產生了抗性,但對于豆大薊馬鈉離子通道結構與功能仍不清楚。因此,本研究對豆大薊馬鈉離子通道基因(MuNav)進行克隆和序列分析,旨在為后續解析豆大薊馬對擬除蟲菊酯抗性機制奠定基礎。
1 材料與方法
1.1 供試昆蟲豆大薊馬于2018年12月采自海南省海口市美蘭區海南大學海甸校區基地,并在實驗室內長期飼養,期間未施用任何藥劑。飼養方法如下:將豆大薊馬成蟲及若蟲轉移至240 mL養蟲罐中,將新鮮豇豆洗凈,切成長度約5 cm的豆莢放入20%蜂蜜水中浸泡1 min,晾干,放入養蟲罐中,置于人工氣候箱(PYX-400Q-A,廣東韶關科力實驗儀器有限公司)中飼養。成若蟲的飼養溫度為26 ℃,濕度為60 %~65 %,光周期16 L∶8 D。
1.2 PCR擴增取豆大薊馬成蟲30~40頭,通過Trizol法提取RNA,并用1 %的瓊脂糖進行凝膠電泳。隨后進行反轉錄,方法參照Prime ScriptTM 1st Strand cDNA Synthesis Kit(東洋紡生物科技有限公司,上海)說明書。根據NCBI西花薊馬(Frankliniella occidentalis)(XM_026422864.1)和棕櫚薊馬(Thrips palmi)(XM_034385472.1)Nav序列,使用Primer 5軟件對MuNav進行通用引物設計,正向引物為:ATGCCGAGTGTCCGGGAGTCG,反向引物為:TCAGACATCCGCAAGGGCCGGAG。以上述cDNA為模板,進行PCR擴增,反應體系如下:cDNA 1 μL、dNTP Mix 1 μL、上下游引物各2 μL、ddH2O 18 μL、2×Phanta Max Buffer (Mg2+plus)25 μL、Phanta Max Super-Fidelity DNA Ploymerase 1 μL。PCR 反應條件如下:95 ℃ 預變性 3 min;95 ℃ 15 s,56 ℃ 15 s,72 ℃ 延伸 6.5 min,共 35 個循環;72 ℃反應后延伸8 min。
1.3 PCR產物回收及連接轉化用Cycle-Pure Kit(OMEGA, America)將上述PCR產物進行純化回收,用NanoDrop 2000分光光度計(賽默飛世爾科技(中國)有限公司,上海,中國)對回收產物進行濃度測定,并置于?20 ℃備用。分別用HindⅢ-HF和X maⅠ對PGH 19進行雙酶切,反應時間為2 h,溫度為37 ℃,將純化回收產物和PGH 19鏈接,反應條件為:37 ℃、45 min,取出后在冰上冷卻5 min,將連接好的產物導入感受態細胞stbl 2中并涂板,方法參照stbl 2感受態細胞(吐露港生物科技有限公司,上海)說明書,將平板倒置在22 ℃的保溫箱中培養。在96孔板每孔中加入200 μL LA培養基,并挑取單克隆置于其中,密封后180 r·min?1,30 ℃ 搖菌 12 h。取 50 μL 菌液加入 5 mL LA 培養基中于 30 ℃、180 r·min?12 次搖菌 16 h,使用 Plasmid mini Kit(OMEGA, America)試劑盒進行質粒提取,并進行測序。
1.4 序列分析及進化樹構建克隆得到MuNav全長,并上傳至NCBI(GenBank accession number:MZ043856),用Simple Modular Architecture Research Tool預測其保守性結構域。同時,從NCBI上下載得到其他58種昆蟲的Nav全長(表1)。隨后,用Clustal Omega軟件(https://www.ebi.ac.uk/Tools/msa/clustalo/)進行多序列比對,并利用Mega 7(DNASTAR, America)軟件,采用最大似然法構建系統發育樹,自舉值設置為1 000。
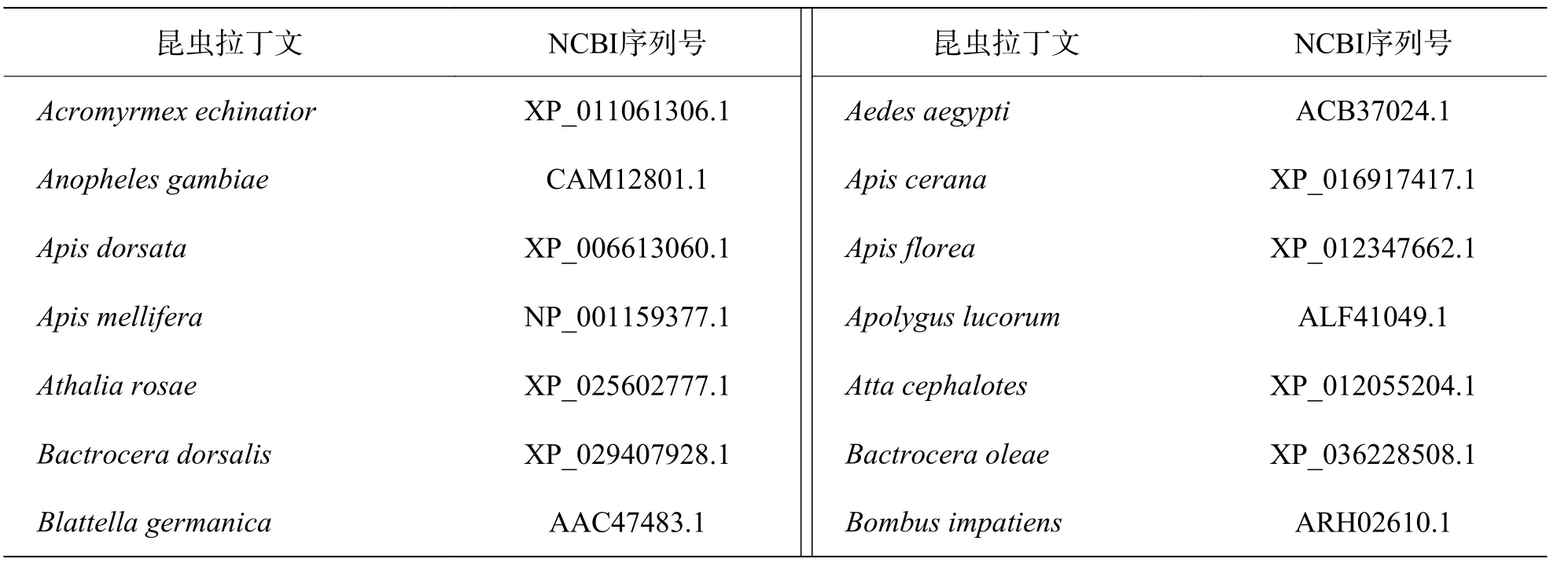
表1 進化樹分析 Nav NCBI 序列號

續表1
2 結果與分析
2.1 豆大薊馬電壓門控鈉離子通道基因克隆及序列分析克隆得到MuNav,其全長為6 279 bp,編碼 2 093 aa (GenBank:MZ043856) (圖1),含有4個同源結構域,結構域 Ⅰ~Ⅳ,每個同源結構域含有6個跨膜片段,第3和第4結構域之間含有“MFM”疏水性殘基,符合昆蟲Nav的典型特征(圖2)。
將MuNav的氨基酸序列與西花薊馬、棕櫚薊馬、綠盲蝽(Apolygus lucorum)、美洲大蠊(Periplaneta americana)Nav進行同源性比對,發現其與棕櫚薊馬Nav序列的相似度高達94.88%,與西花薊馬Nav序列的相似度同樣高達94.68%,而與美洲大蠊和綠盲蝽Nav的相似度分別僅為80.74%和80.19%,表明豆大薊馬與薊馬類昆蟲的親緣關系較近,而與其他目昆蟲則發生了分化(圖3)。
2.2 豆大薊馬鈉離子通道系統發育分析進化樹的結果表明(圖4),MuNav與其他昆蟲均存在一定的親緣關系。其中,豆大薊馬與棕櫚薊馬(T.palmi)、煙薊馬(Thrips tabaci) 和西花薊馬聚為一類,顯示薊馬類昆蟲Nav符合進化過程,即同一類昆蟲聚類在相近位置。反之,豆大薊馬與其他目的昆蟲,如黑腹果蠅(Drosophila melanogaster)、德國小蠊(Blattella germanica)、綠盲蝽等則發生了較大程度的分化,親緣關系相對較遠。
3 討 論
大部分昆蟲僅編碼1條或2條Nav,昆蟲Nav由1個分子量約為260 kD的α亞基和5個輔助小亞基組成,α亞基具有Nav活性,而輔助小亞基則起到調節Nav表達的作用。本研究通過PCR的方法克隆得到了MuNav全長序列,發現其與其他昆蟲Nav結構類似,有4個同源性極高的跨膜結構域,即結構域I、結構域II、結構域III和結構域IV組成,且每個結構域均包含6個疏水性跨膜螺旋體(S1~S6)。在MuNav的P-LOOP環上同樣鑒定到與其他昆蟲類似的4個保守性氨基酸殘基D、 E、 K、A和1個關鍵失活閥門MFM,這些結果均表明,MuNav具有昆蟲Nav特有的全部結構特征,這為后續MuNav功能研究奠定了基礎。
本研究的序列比對結果表明,豆大薊馬與其他目昆蟲,如蜚蠊目美洲大蠊和半翅目綠盲蝽Nav的序列相似性均高達80%以上,這為MuNav功能研究提供了重要參考。值得一提的是,豆大薊馬與其他薊馬類昆蟲序列的相似性極高,如西花薊馬和棕櫚薊馬的序列相似性高達94%以上,表明纓翅目薊馬類昆蟲Nav的結構與功能極為保守。另外,基于NCBI上獲取得到的59種昆蟲Nav序列,構建了系統發育樹。與上述序列比對結果類似,進化樹結果表明雖然昆蟲Nav具有一定的保守性,但豆大薊馬與其他薊馬類昆蟲聚為一類,表明且它們在進化過程中極為保守,即發生的變異較小,而與其他昆蟲則形成了不同的分支,表明它們發生了一定程度的分化。
昆蟲Nav可通過轉錄后修飾,即選擇性剪切和RNA編輯實現其功能的多樣性[17?18]。RNA編輯是mRNA在轉錄水平上通過堿基插入、缺失或替換,從而引起氨基酸改變,擴大了生物的遺傳信息,增加Nav結構與功能的多樣性,從而幫助其迅速適應外界環境的變化,其研究在過去的幾年里逐漸得到重視[17,19?20]。昆蟲Nav RNA編輯存在2種形式:一種是腺苷A去氨基轉變為次黃嘌呤I,即A至I編輯,另外一種是U至C編輯。其中A至I的RNA編輯在昆蟲中尤為常見,可導致蛋白質的結構和功能發生改變。如在黑腹果蠅、德國小蠊和綠盲蝽中均被鑒定到,并可引起昆蟲神經性興奮[21?23]。昆蟲RNA編輯存在組織特異性,A-I RNA編輯主要發生在編碼離子通道、神經遞質受體或G蛋白偶聯受體的神經系統基因轉錄本中[24?25]。例如,在黑腹果蠅鈉通道轉錄本中發現了11個A-I編輯[26],在擬果蠅中發現了3個A-I的RNA編輯,分別是Q52R、C189Y和N1260D[27],而在德國小蠊和綠盲蝽Nav中也分別發現了2個和1個A-I RNA編輯[21,23]。此外,昆蟲Nav還存在U-C編輯,該類型的RNA編輯一般存在卵巢和腸道中,如黑腹果蠅和德國小蠊體內F1919S位點的U-C編輯可產生持續電流[20]。
近年來,隨著田間用藥量的增加,昆蟲對擬除蟲菊酯的抗藥性不斷增加,Nav突變也越來越頻繁,在很多高抗藥性種群中發現多個突變位點。大部分昆蟲中最常見的突變發生在 1 014位點,該位點在家蠅中最早被發現,由L突變為F,使家蠅對氯菊酯的敏感性降低了至少10倍,并且增加了擬除蟲菊酯誘導的鈉尾電流的衰減率[28]。除了家蠅外,在岡比亞按蚊(Anopheles gambiae)、人蚤(Pulex irritans)、中華按蚊(Anopheles sinensis)、致倦庫蚊Culex quinquefasciatus)等昆蟲中也發現了L1014 突變[29?32]。除了 1 014 位點外,918 位點氨基酸發生改變也會影響擬除蟲菊酯藥劑的敏感性。1999年,LEE等[33]對家蠅鈉通道918位點進行定點修飾,用蘇氨酸代替甲硫氨酸,通過雙電極電壓鉗進行驗證,發現其對高濃度的氯氰菊酯完全不敏感。在白紋伊蚊體內發現了I1532T和F1534S/L,當這2個突變單獨出現時顯著降低了對I型擬除蟲菊酯類藥劑氯菊酯和聯苯菊酯的敏感性,但是對Ⅱ型擬除蟲菊酯類藥劑敏感性沒有顯著影響[34]。本研究未在MuNav中發現918和1 014位點突變,但在對序列分析時發現了T929I突變,其在對擬除蟲菊酯殺蟲劑產生抗性過程中的具體作用還需進一步實驗探究。
在高抗種群中,突變點往往都不是單個出現的,它們會同時出現2個突變點,大部分情況下該類型突往往比單點突變更高的抗藥性。在蚊子中,Nav中單獨的N1575Y突變不會改變昆蟲對擬除蟲菊的敏感性,但N1575Y和L1014F雙突變的種群對氯氰菊酯的抗性是野生型的80倍,對溴氰菊酯的抗性是野生型的53倍,相較于單突變分別增加了3.4倍和9.8倍[35]。在煙薊馬中,T929I的單突變會導致煙薊馬對氯氰菊酯產生中等水平抗性,當T929I和K1774N雙突變共同出現會導致煙薊馬對氯氰菊酯產生高等水平抗性(RR=2 700)[36]。反之,某些Nav雙突變時,并不會降低鈉通道的抗性,例如在埃及伊蚊體內,無論是V1023G/S996P還是V1023G/D1794Y對擬除蟲菊酯的抗性都沒有單一突變體高[37]。此外,有些位點單獨突變不會改變鈉通道的敏感性,但會顯著降低其他位點對擬除蟲菊酯殺蟲劑的敏感性。例如,蟑螂體內E434K和C764R單獨出現不會改變鈉通道對溴氰菊酯的敏感性,但是E434K或C764R與L993F雙突變時鈉通道對溴氰菊酯的敏感性會降低100倍[38]。除單突變,雙突變外,昆蟲鈉通道有時還會出現三突變,例如在蟑螂Nav中引入V409M突變,可使其對溴氰菊酯的敏感性降低了10倍,而當V409M、E434K、C764R 3個突變同時出現時,蟑螂對溴氰菊酯的敏感性降低了100倍[39]。除上述突變類型,昆蟲體內Nav突變還會出現連鎖反應,在致倦庫蚊中,M943V和I973T就存在著明顯的連鎖反應,但是其生理意義還需進一步探索[40]。雖然在MuNav中未發現918和1 014位點突變,但豆大薊馬鈉離子通道出現眾多潛在的抗性位點(未發表),仍需用雙電極電壓鉗技術進一步驗證。
本研究克隆了豆大薊馬鈉離子通道全長序列,不僅擴展了昆蟲鈉離子通道的遺傳信息,也為后續豆大薊馬抗性位點的鑒定,kdr抗性突變位點分子診斷技術的研發和探明化學殺蟲劑與MuNav的結合能力奠定了基礎,同時為延緩豆大薊馬抗藥性的發展及抗藥性治理指明了方向。
