基于55K SNP芯片分析的優質小麥的遺傳多樣性
2022-05-20 01:37:28彭義峰李亞青張士昌李孟軍何明琦
麥類作物學報 2022年3期
張 楠,彭義峰,李亞青,張士昌,李孟軍,何明琦
(石家莊市農林科學研究院,河北省小麥工程技術研究中心,河北石家莊 050041)
隨著人民生活水平的不斷提高,市場對優質小麥的需求量不斷增長。當前,優質小麥生產面臨著大面積推廣應用品種少、品種抗逆性不佳、品質不穩定等問題。在優質小麥育種實踐中,可選用的育種親本數目有限,僅師欒02-1、藁優2018、濟南17、新麥26、西農979等優質強筋小麥可作為育種的親本。利用分子標記(如SSR、SNP芯片)分析優質小麥品種的親緣關系和遺傳相似度,對優質小麥種質創新和育種具有重要意義,但目前有關優質小麥遺傳多樣性的研究報道較少。利用SNP芯片對中國主要優質小麥品種的遺傳多樣性進行系統研究尚未見報道。
小麥基因組測序工作的不斷深入,極大促進了SNP標記的研究與開發。SNP芯片具有標記數量多、通量高等優點。目前,已開發的小麥SNP芯片主要有9K iSelect、90K iSelect、820K Axiom array、35K Axiom array、50K TraitBreed、15K、660K Axiom array、55K等。研究表明,小麥SNP芯片是研究小麥遺傳多樣性、群體結構和QTL定位的有效技術手段。與小麥其他SNP芯片比較,55K SNP芯片的標記有效性更高,已用于QTL定位、農藝性狀關聯分析、遺傳多樣性分析等研究。本研究選取中國主要優質小麥品種,利用55K SNP芯片揭示其遺傳多樣性和親緣關系,以期為優質小麥育種親本選配、種質資源創制和分子標記輔助選擇提供參考依據。
1 材料與方法
1.1 供試材料
45個優質小麥品種來自國家產業體系歷年種質資源征集品種;5個優質高代品系(品鑒01、品鑒02、品鑒03、品鑒04和品鑒05)由石家莊市農林科學研究院小麥研究所提供。所有材料現保存于石家莊市農林科學研究院分子育種實驗室,品種(系)的具體信息見表1。
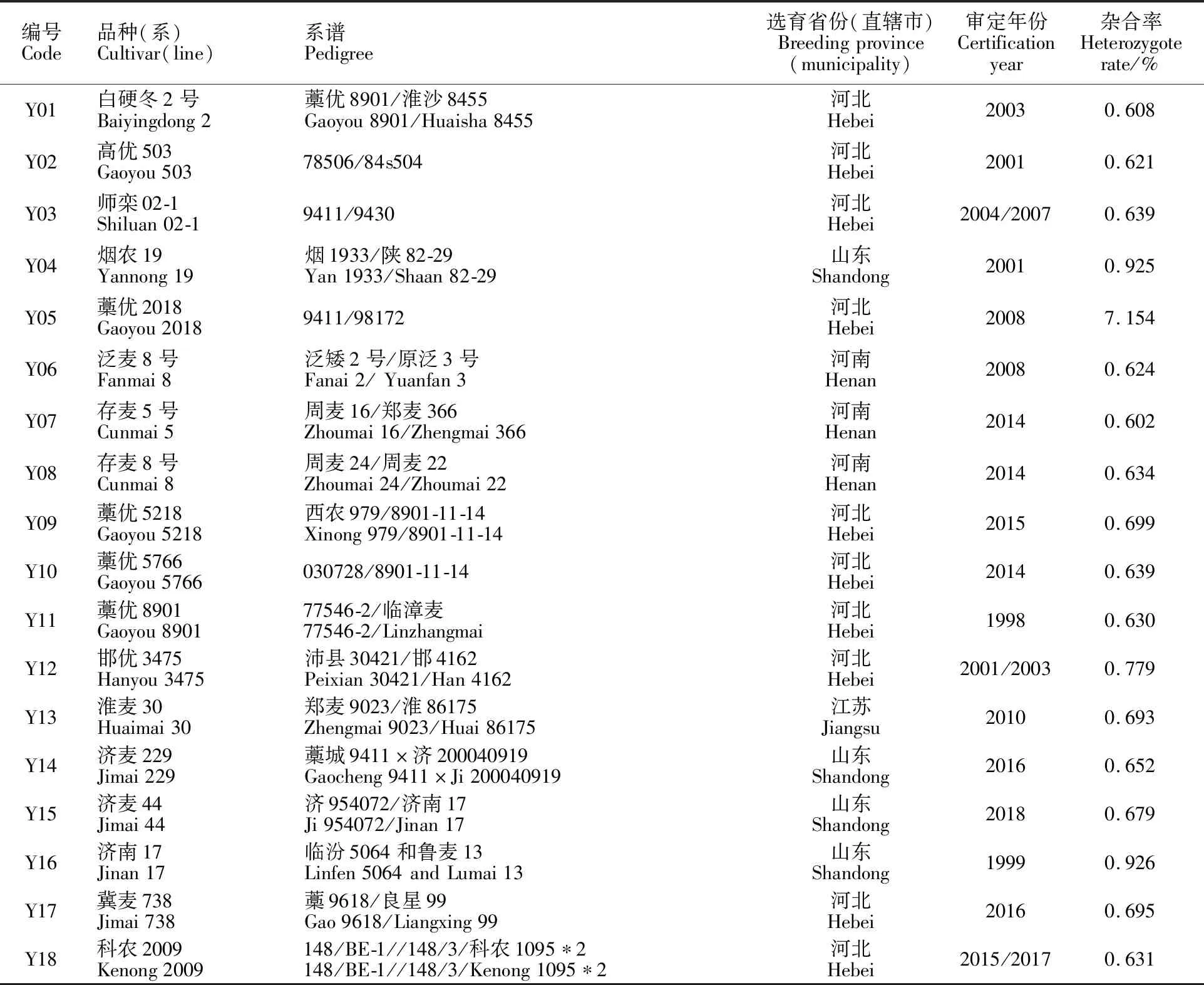
表1 供試小麥品種(系)的來源及其雜合率Table 1 Origin and heterozygote rate of wheat cultivars(lines)used in the study

(續表1 Continued table 1)
1.2 基因組DNA提取
用于SNP芯片分析的50個小麥品種(品系)為經過連續2代田間選擇的單株。試驗材料在植物生長室中培養,待長至二葉一心時,取小麥葉片,用CTAB法提取基因組DNA。
1.3 SNP標記分析
1.4 統計分析
對Dish QC(DQC)>0.82和標記檢出率(CR)>95% 的樣品進行SNP位點質控。以缺失率(MISS)<0.1和最小等位基因頻率(MAF)>0.05為標準對SNP標記進行過濾。以雜合標記數除以總標記數計算品種的雜合率。以基因型一致的標記數除以成功分型的標記數計算品種的遺傳相似度。使用Structure軟件進行群體結構分析。使用TreeBest軟件的NJ-tree模型構建進化樹。
2 結果與分析
2.1 染色體低多態性區段
染色體低多態性區段是指SNP多態性低的染色體區段,由MAF值較低的一段SNP位點組成。55K SNP芯片共有53 007個探針位點,通過對DQC>0.82 和CR>95%的樣本進行SNP位點質控,共獲得36 691個有效標記,去除無染色體和物理位置的標記,最終獲得33 876個有效標記,可用于染色體低多態性區段分析。其中,多態性標記有33 202個,占有效標記數的98.01%。B基因組多態性位點最多,為13 207個,占比 39.78%,其中4B染色體最多;A基因組上多態性位點有12 873個,占比38.77%,其中5A染色體上最多;D基因組上多態性位點最少,為7 122個,占比21.45%,4D和5D染色體上分別僅有648和674個。多態性SNP位點在部分同源群上的分布頻率大小依次為7號染色體>4號染色體>3號染色體>2號染色體>5號染色體>1號染色體>6號染色體。
在33 876個有效標記中,MAF在0.00~ 0.20之間的SNP標記占比31.45%(圖1A),因此,進一步分析閾值為0.10和0.05兩種情況(圖1B和1C)。低多態性區段在優質小麥品種(系)染色體上的分布表明,A基因組和D基因組染色體上的低多態性區段均多于B基因組。6A、4D和6D染色體上的低多態性區段多于其他染色體 (圖1)。
2.2 品種的雜合率
雜合率反映優質小麥品種(系)在基因組水平上的雜合程度。一般而言,自交代數越高的材料,其雜合率越低。利用55K SNP芯片對50個小麥品種(系)進行分析,結果(表1)發現,在45個優質小麥品種中,除藁優2018的雜合率為7.154%外,其余44個優質小麥品種的雜合率變化范圍為0.496%~1.457%,平均值為0.849%,表明這些優質小麥品種的純合程度整體較高。5個優質高代品系中,品鑒2為F代株系,雜合率高達 8.399%,但其群體農藝性狀表現一致。
2.3 基于SNP標記的品種間遺傳相似度
遺傳相似度分析能夠為優質小麥親本組配(相似度低的材料)和群體改良(相似度高的材料)提供數據支撐。45個優質小麥品種間的遺傳相似度系數(genetic similarity,GS)變化范圍為 0.488~0.928,平均值為0.603;有1 944個GS變化范圍為0.500~0.800,占98.18%,其中 0.500~0.600、0.600~0.700和0.700~0.800分別占54.34%、38.18%和5.66%。GS小于 0.500的有20個(1.01%),大于0.800的有16個(0.81%)。因此,中國優質小麥品種間具有相對較低的遺傳多樣性。
河北省15個優質小麥品種間的GS變化范圍為0.467~0.928,平均值為0.618;山東省8個優質品種間的GS變化范圍為0.557~0.740,平均值為0.638;河南省9個優質品種間的GS變化范圍為0.564~0.736,平均值為0.630。可以看出,河北省優質小麥品種間的GS低于山東省和河南省。系譜分析顯示,這種現象與河北省優質小麥的育種親本較為廣泛有關。
品種間GS低于0.500的優質小麥品種為:(1)師欒02-1和藁優2018;(2)藁優2018和藁優5818;(3)高優503和龍麥40;(4)藁優8901和西農889;(5)藁優8901和藁優2018;(6)白硬冬和龍麥40;(7)邯優3475和鎮麥168;(8)泛麥8號和藁優8901;(9)藁優2018和科農2009;(10)冀麥738和運旱618,這些優質小麥品種分別來自河北、河南、黑龍江、江蘇、陜西和山西,親緣關系較遠。品種間GS高于0.800的優質小麥品種為:(1)西農20和存麥5號;(2)石4366和石優17;(3)石優20和濟南17;(4)中麥578和濟麥44;(5)西農979、西農9718、藁優5218和藁優5818;(6)龍麥26和龍麥40;(7)藁優5766和藁優5218。其中龍麥26和龍麥40、藁優5766和藁優5218的GS均高于0.900。這些品種屬于近似品種,在優質小麥種質創新和育種中應予以注意。

a、b和c分別表示染色體上0.00≤MAF≤0.20、0.00≤MAF≤0.10和0.00≤MAF≤0.05的低多態性區段。藍色表示小于等于MAF閾值的多態性區段,紅色表示大于閾值的多態性區段。a,b and c indicate low-diversity chromosomal regions with 0.00≤MAF≤0.20,0.00≤MAF≤0.10 and 0.00≤MAF≤0.05,respectively.Red indicates the MAFs of chromosomal regions are higher than threshold value,and blue indicates the other regions.圖1 低多態性區段在優質小麥品種(系)染色體上的分布Fig.1 Distribution of low-diversity regions on high-quality wheat chromosomes
2.4 群體結構
群體結構分析是按照一定標準將一個群體分為若干亞群,處于同一亞群內的個體親緣關系較高。群體結構分析可了解自然群體的遺傳多樣性和遺傳分化情況。在群體結構分析中,最低的交叉驗證誤差(cross-validation error,CV error)對應的值(群數)是最理想的選擇。在50個小麥品種(系)中,=2 時,CV errors最低(1.15),因此,選擇=2進行進一步分析。當=2時,有12個優質小麥品種(系)血統混雜,在不同的群結構分析中可能發生變化,但不影響整體分群評估(圖2A)。
2.5 進化樹
使用TreeBest軟件的NJ-tree模型將50個小麥品種(系)分為2個類群,這與群體結構分析結果一致。其中,類群1包含6個亞群(亞群Ⅰ~Ⅵ),有39個品種(系);類群2僅包含亞群Ⅶ,有11個品種(系)。類群1中,亞群I包含6個品種(系),其中品鑒01與中麥578親緣關系較近,聚類在一個分支上。亞群Ⅱ包含5個品種,其中,石優17是石4366的父本,聚類在一個分支上。亞群Ⅲ包含3個品種,濟南17是石優20和新麥26的父本。亞群Ⅳ包含6個品種(系),其中,鄭麥7698是品鑒03、品鑒04和品鑒05的父本;鄭麥366是西農20和存麥5號的共同親本。亞群Ⅴ包含10個品種,其中鄭麥98、煙農21、西農889、泛麥8號、鄭麥366血統混雜,該亞群聚類結果與系譜關系不明;亞群Ⅵ包含9個品種,有2個分支,其中,1個分支是西農2611或西農979的后代;另1個分支包含4個品種,其中3個為龍麥系列品種,龍麥26是龍麥35的父本。類群2中,濟麥229、邯麥18、冀麥738、龍麥20和邯優3475血統混雜。類群2有2個分支,其中1個分支為臨漳麥或藁優8901的后代,另1個分支中,邯麥18、師欒02-1和冀麥738均具有藁優8901血統,龍麥20和邯優3475的聚類結果與系譜關系不明(圖2B)。
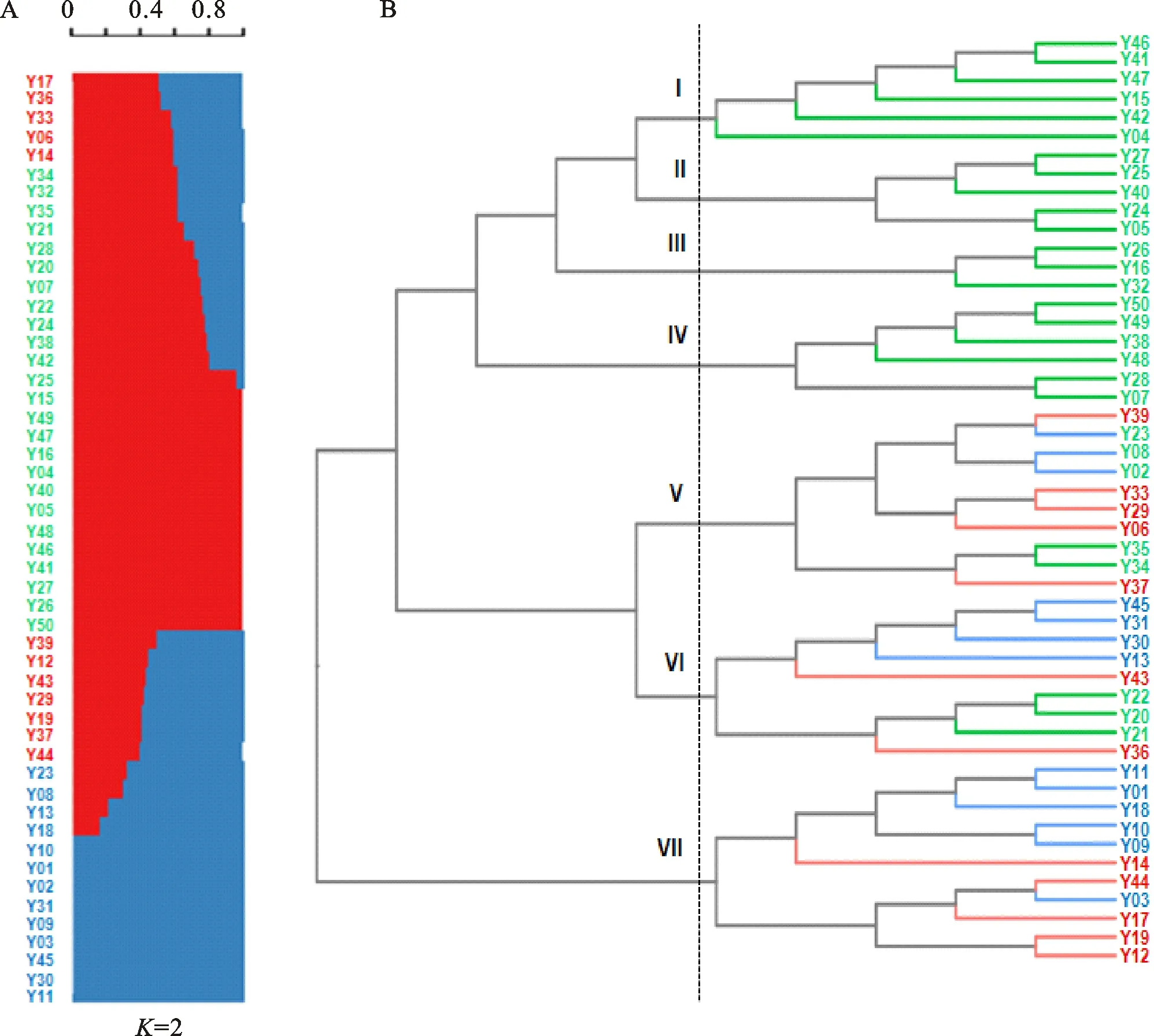
A:群體結構分析;B:進化樹分析。品種(系)編號對應的品種(系)見表1。綠色和藍色字體表示不同類群,圖B中分枝線條顏色與圖A不同類群對應;紅色字體表示血統混雜品種(系)。A:Population structure analysis;B:Phylogeny tree analysis.Cultivars(lines)correspond to numbers showed in table 1.Green and red fonts represent different population,and the colors of branch lines in figure B correspond to different groups in figure A;Red font represents mixed ancestry cultivars(lines).圖2 供試小麥品種(系)的群體遺傳結構和進化樹分析Fig.2 Population structure and phylogeny tree analysis of the tested wheat cultivars(lines)
3 討 論
本研究發現,55K SNP芯片中有效標記和多態性標記數量分別占總標記數量的63.91%和62.64%。有效標記中多態性標記占比高達 98.01%。D基因組中有效標記和多態性標記數量顯著低于A基因組和B 基因組,但多態性比例無顯著差異。7個部分同源群上的有效標記數量為3 686~5 051個,相應多態性標記占比 97.38%~98.90%。且55K SNP芯片中有效標記的多態性以及在不同基因組和7個部分同源群上的分布均優于90K SNP芯片。因此,55K SNP 芯片適用于對中國優質小麥開展遺傳多樣性研究。
GS反映不同小麥品種(系)間在基因組水平上的相似程度。鄭永勝等基于SSR標記,建議在DUS測試中與申請品種遺傳相似度系數高于0.800的已知品種作為近似品種進行田間種植和特異性評價。王立新等利用Genomic-SSR、EST-SSR、AFLP-SCAR標記篩查547個國家冬小麥區域試驗品系及134個國審品種,發現絕大多數相似品種(系)間的GS 均大于0.900。曹廷杰等利用90K SNP芯片對96個河南省小麥品種進行遺傳多樣性分析,發現品種間GS變化范圍為0.552~0.998,平均值為0.719,94.3%的品種GS分布在0.652~0.812 之間;李珊珊等利用90K SNP芯片對143份河北省推廣小麥品種進行遺傳多樣性分析,發現品種間GS變化范圍為 0.438~0.997,平均值為0.602,89.23%的品種間GS分布在0.520~0.730之間。本研究表明,中國優質小麥GS變化范圍為0.488~0.928,平均值為0.603,98.18%的品種間GS分布在 0.500~0.800之間。這表明,中國優質小麥品種遺傳多樣性總體不高。隨著中國優質小麥品種審定標準的提高,優質小麥的遺傳多樣性有進一步降低的趨勢。在45個優質小麥品種中,0.81%的優質小麥品種間GS高于0.800,但其農藝性狀存在顯著差異,因此,這些品種均具有特異性,符合現行國家小麥品種審定標準。SNP芯片探針根據基因組SNP設計,很多標記與重要農藝性狀基因無關,可能是導致上述現象的原因之一。
本研究群體結構和進化樹分析結果表明,50個供試小麥品種(系)僅分成 2個類群7個亞群,同時,2個類群間存在較嚴重的混血現象,這種現象是遺傳多樣性較低的一種表現。50個小麥品種(系)有12個血統混雜,這些血統混雜品種在不同群結構分析中變化較大,但不影響整體分群評估。聚類結果與系譜比較分析表明,除亞群Ⅴ外,供試小麥品種(系)聚類分析與系譜分析結果基本吻合。因此,利用55K SNP芯片能夠較好地反映中國小麥品種(系)的遺傳多樣性和親緣關系。
在50個供試小麥品種(系)中,多態性SNP標記數量在B基因組中最多,A基因組次之,D基因組最少,與利用RFLP標記、SSR標記和90K SNP芯片對小麥遺傳多樣性的研究結果一致。A基因組和D基因組的染色體低多態性區段均多于B基因組,這與多態性標記的分布規律相符合。6A、4D和6D染色體上的低多態性區段多于其他染色體,這表明,在優質小麥育種過程中,6A和6D染色體這些區段受到高強度選擇。一般認為,六倍體小麥D基因組的遺傳多樣性較低,4D染色體上的多態性標記最少。本研究在50個供試優質小麥品種(系)中,4D染色體上的多態性標記也最少(674個),多態性標記在有效標記中的比例也最低 (88.77%)。供試小麥品種(系)染色體上的SNP標記MAF在 0.00~0.20之間的占比31.45%,且4D染色體上占比39.04%,這表明,4D染色體在優質小麥育種過程中也受到了高強度選擇。
