內蒙古苜蓿銹菌越冬的分子定量檢測
2022-05-19 12:49:04馬麗杰武占敏張俊杰楊政偉卓瑪曲措胡小平
西北農業學報 2022年4期
馬麗杰,武占敏,張俊杰,呂 文,張 偉,楊政偉,卓瑪曲措,胡小平
(1.鄂爾多斯生態環境職業學院,內蒙古鄂爾多斯 017010;2.西北農林科技大學 植物保護學院,農業農村部黃土高原作物有害生物綜合治理重點實驗室,陜西楊凌 712100;3.鄂爾多斯市植保植檢站,內蒙古鄂爾多斯 017010;4.鄂爾多斯市農產品質量安全中心,內蒙古鄂爾多斯 017010;5.西藏農牧學院 植物科學學院,西藏林芝 860000)
苜蓿是世界上種植面積最大的豆科牧草,也是中國近年來大面積推廣和種植的優良牧草[1]。內蒙古是國內五大牧草種植區之一,苜蓿商品草種植面積和產量居全國第二[2]。由苜蓿條紋單孢銹菌(Uromycesstriatus)引起的苜蓿銹病是中國北方苜蓿種植區普遍發生的重要流行性葉部病害,發生嚴重時可導致苜蓿減產10%~30%,種子減產50%[3-4],病葉的鮮質量比健康葉片減少44%,粗蛋白含量下降18%,粗纖維含量增加15%[5],苜蓿莖葉枯黃不能飼用[3],大幅度降低苜蓿產量和品質。內蒙古中部地區苜蓿最后一次刈割一般在10月下旬結束,殘存的苜蓿銹菌很可能在苜蓿或其轉主寄主上潛伏侵染并越冬,成功越冬的病菌可成為翌年病害流行的初侵染源。關于苜蓿銹菌的越冬部位及越冬形態,目前有四種觀點:第一,苜蓿銹菌可在轉主寄主乳漿大戟等的地下器官內以菌絲體形態越冬[6-8];第二,苜蓿銹菌可以冬孢子形態在病殘體上越冬[9]。第二年侵染其轉主寄主大戟并在其上完成有性世代,形成銹孢子后,由銹孢子轉染到苜蓿上,使之發病[10];第三,苜蓿銹菌可以直接在苜蓿根莖內越冬[3];第四,在冬季較溫暖的地區如伊拉克[11],苜蓿銹菌也能以夏孢子形態越冬[10,12]。
上述關于苜蓿銹菌越冬部位的研究,大多根據田間調查經驗推斷所得,缺乏直接試驗證據。目前,內蒙古中部地區苜蓿銹菌越冬的菌量和越冬部位還不清楚。因此,本研究建立一種基于RNA水平的real-time qPCR技術,對內蒙古中部5個苜蓿主產區返青后植株內的銹菌量進行檢測,明確苜蓿銹菌越冬菌量和越冬部位,為苜蓿銹病的綜合防控提供理論指導。
1 材料與方法
1.1 調查點選擇
2018-2020年,選擇內蒙古中部5個苜蓿主產區作為調查點,自東向西分別為烏蘭察布市、呼和浩特市、包頭市、鄂爾多斯市和巴彥淖爾市 (圖1)。
圖1 內蒙古苜蓿銹病調查點
1.2 試驗方法
1.2.1 植株標記 2018-2020年每年11月中下旬,對每個調查點采用5點取樣法標記50株冬前發病的植株,翌年4月初,從冬前標記現已返青的植株上從上向下剪取5 cm長度的植株材料裝于含有RNAlaterTMSoIn保存溶液(Invitrogen, AM7021)的10 mL離心管中,備用。
1.2.2 田間樣品處理 將田間樣品從離心管中取出,用吸水紙吸掉殘留溶液,用蒸餾水反復沖洗干凈并吸干水分,剪碎混勻,稱取100 mg,備用。
1.3 建立RNA水平的Real-time qPCR體系
1.3.1 RNA提取、純化并反轉錄成cDNA 稱取發病的苜蓿銹病葉片100 mg(約5片),液氮研磨成粉末,用SV Total RNA Isolation System(Promega)試劑盒提取總RNA。用Nanodrop 2000(Thermo Fisher)儀器測RNA總濃度。最終獲得總RNA,其濃度為145 ng/μL,OD260/280為2.1,OD260/230為2.2,RNA完整性好。根據Thermo scientific ReverAid First Stand cDNA Synthesis kit試劑盒進行RNA純化和反轉錄。
1.3.2 苜蓿銹菌特異性引物設計與合成 根據文獻[13-14]報道,Eric Kemen設計特異性引物將苜蓿銹菌(Uromycesstriatus)和同源性較高的蠶豆銹菌(Uromycesfabae)進行區分,引物合成見表1的Us-RTP1-NLS1。采用Us-RTP1-NLS1的下游引物,并在Us-RTP1基因組上重新設計上游引物,合成新引物Us-PTP1-NLS2(表1)。通過序列比對軟件BioEdit比對2對引物,結果顯示2對引物只有2個堿基的差異(圖2)。將2對引物分別在NCBI上進行比對,發現其與苜蓿上常見的其他4種葉部病害苜蓿褐斑病(苜蓿假盤菌Pseudopezizamedicaginis),苜蓿葉腫病(小光殼屬Leptosphaerulinabriosiana),苜蓿灰斑病(尾孢菌屬Cercosporamedicaginis)和苜蓿炭疽病(炭疽屬Colletotrichumspp.)均無匹配。
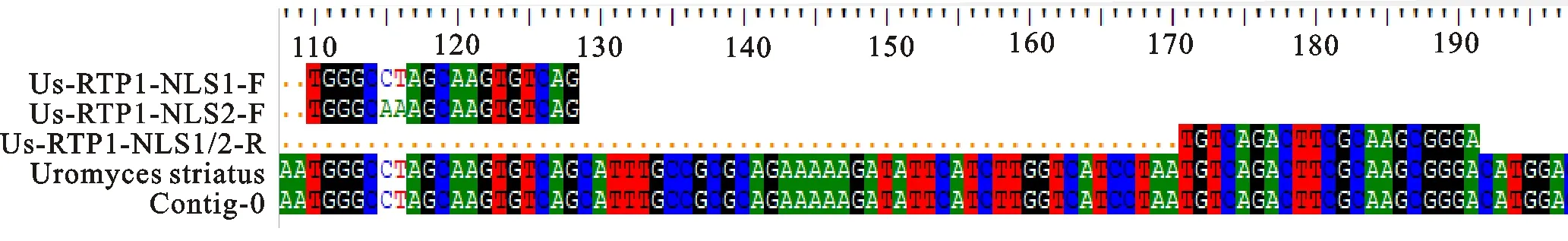
圖2 兩對引物在苜蓿銹菌基因組上的比對
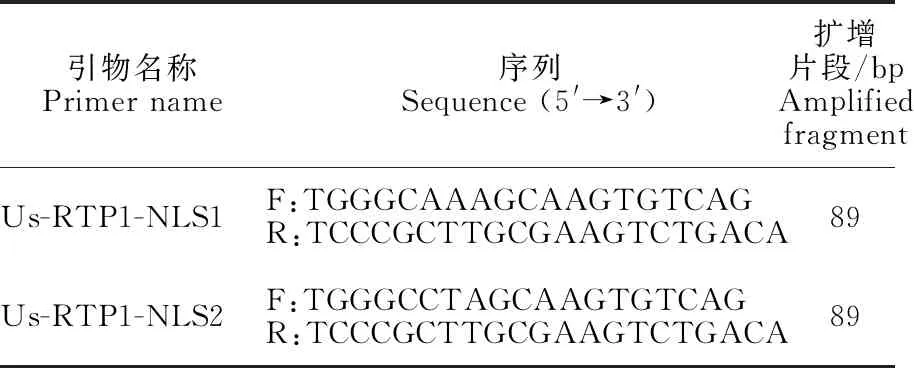
表1 苜蓿銹菌特異性引物
1.3.3 苜蓿組織特異性引物合成 選用文獻[15]中擴增苜蓿葉片組織基因PDF2、UBC和Ubiquitin的3對引物進行合成并擴增(表2)。
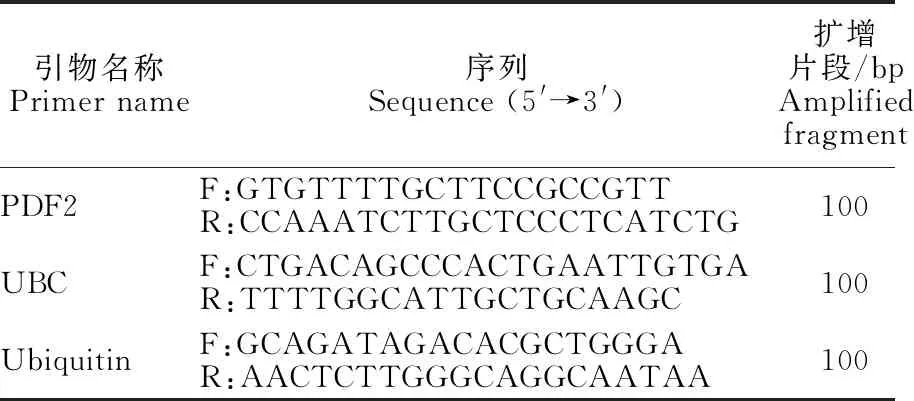
表2 苜蓿葉片組織特異性引物
1.3.4 標準曲線的構建 根據UltraSYBR Mixture(CWBIO)試劑盒說明書,進行標準曲線的構建。以獲取的初始cDNA為模板進行10倍梯度稀釋,最終獲得2.03、2.03×10-1、 2.03×10-2、2.03×10-3、2.03×10-4、2.03×10-5、 2.03×10-6、2.03×10-7ng/μL 8個濃度梯度cDNA,分別利用苜蓿銹菌特異性引物和苜蓿葉片組織特異性引物進行qPCR擴增,每個梯度樣品重復3次。
1.4 指標測定方法
每個cDNA樣品由苜蓿銹菌cDNA和苜蓿葉片組織cDNA組成。經標準曲線換算,以分子病情指數(molecular-detected disease index, MDI)進行定義并測算[16]:
MDI=苜蓿銹菌cDNA量(pg)/苜蓿葉片組織cDNA量(ng)×100%
1.5 返青后苜蓿植株發病率調查
在6月中旬,利用5點取樣法對各監測點苜蓿銹病發病情況進行調查。每點調查20株,每個監測點共調查100株,記錄植株發病率。
植株發病率(%)=發病的植株數/調查的總株數×100%
1.6 數據分析
采用SPSS 17.0軟件對所測數據進行統計分析,用“平均值±標準誤”的形式表示測定結果,分別對同一年份不同地區和同一地區不同年份的分子病情指數進行單因素方差分析,用S-N-K法對各測定數據進行多重比較,對不同地區不同年份進行單變量方差分析,采用Excel 2010制圖。
2 結果與分析
2.1 基于RNA水平的real-time qPCR體系構建
2.1.1 引物特異性驗證 利用普通PCR對2對苜蓿銹菌引物的特異性進行驗證,2對引物均可以擴出目標條帶且位置大小一致(圖3-A),后續以Us-RTP1-NLS2引物進行銹菌qPCR定量研究。利用苜蓿葉片特異性引物進行普通PCR擴增,PDF2引物擴增條帶亮(圖3-B),后續以PDF2引物進行苜蓿葉片組織qPCR定量研究。

A.苜蓿銹菌 Us-RTP1基因的擴增片段條帶,其中,泳道1是陰性對照,泳道2和3是相同引物Us-RTP1-NLS1擴增條帶,泳道4和5是相同引物Us-RTP1-NLS2擴增條帶,泳道6是Marker
2.1.2 標準曲線構建 以2.03、2.03×10-1、 2.03×10-2、 2.03×10-3、2.03×10-4ng/μL cDNA為模板進行qPCR,構建發病葉片中苜蓿銹菌cDNA標準曲線(圖4-A)。以2.03×10-3、2.03×10-4、2.03×10-5、2.03×10-6、2.03×10-7ng/μL cDNA為模板進行qPCR,構建發病葉片組織cDNA標準曲線(圖4-B)。樣品中苜蓿銹菌的量遠低于苜蓿葉片組織的量。

A.苜蓿銹菌cDNA標準曲線;B.苜蓿葉片組織cDNA標準曲線
2.2 苜蓿銹菌越冬菌量的測定
2.2.1 不同地區間越冬菌量比較 2018-2020年,利用建立的RNA水平real-time qPCR體系對內蒙古5個苜蓿主產區樣品進行定量檢測。烏蘭察布市、呼和浩特市、包頭市、鄂爾多斯市、巴彥淖爾市3 a平均分子病情指數MDI分別為 0.139、0.284、0.196、0.569、0.280。其中,鄂爾多斯市3 a平均MDI顯著高于其他4個地區的平均MDI(P=0.005)(圖5)。
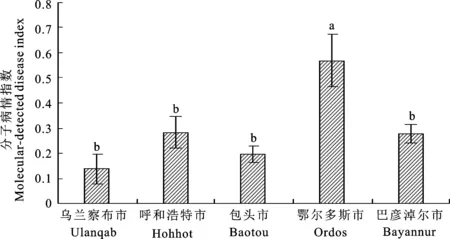
不同小寫字母代表在0.05水平差異顯著
2.2.2 不同年份間越冬菌量比較 2019年,檢測到5個地區苜蓿銹菌平均MDI為0.43,顯著高于2018年的MDI 0.24(P<0.05)和2020年的MDI 0.13(P<0.001)。2018年,5個地區的平均MDI無顯著差異(表3)。2019年,鄂爾多斯市的平均MDI為1.02,顯著高于其他4個地區(P< 0.001)。2020年,鄂爾多斯市的平均MDI為0.20,與巴彥淖爾市和呼和浩特市無顯著差異,但顯著高于包頭市(P=0.004)和烏蘭察布市(P=0.001)。
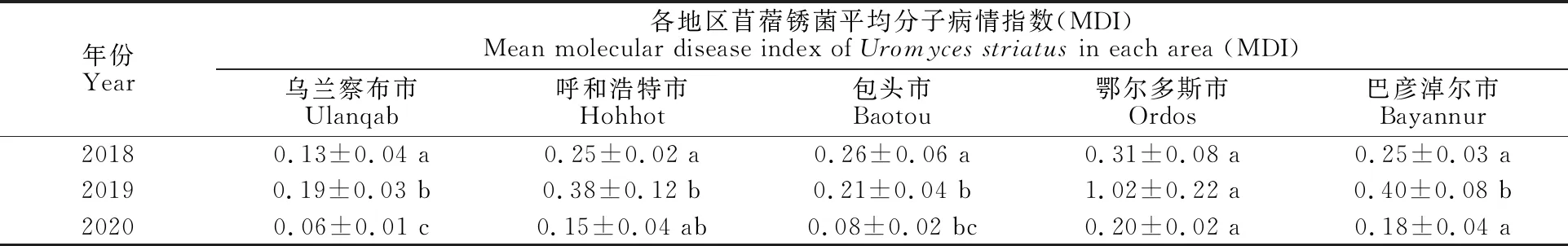
表3 各地區在不同年份間苜蓿銹菌分子病情指數MDI的方差分析
以苜蓿銹菌平均分子病情指數MDI作為因變量,對3 a(2018-2020年)、5個地區(烏蘭察布市、呼和浩特市、包頭市、鄂爾多斯市、巴彥淖爾市)分子病情指數MDI進行方差分析(表4),結果顯示,年份和地區內均存在極顯著差異(P< 0.001),年份*地區之間存在顯著差異(P< 0.05)。

表4 不同處理不同年份不同地區苜蓿銹菌分子病情指數MDI的方差分析
2.3 苜蓿銹菌越冬菌量與返青后植株發病率的關系
2018-2020年,對返青后苜蓿植株發病率進行田間調查統計,并與當年苜蓿銹菌越冬菌量進行比較,各監測點在相同年份下的分子病情指數和返青后植株發病率趨勢基本一致(圖6)。對2018-2020年的苜蓿銹菌越冬菌量和返青后植株發病率進行雙變量相關性檢測,結果顯示,苜蓿銹菌越冬菌量和返青后苜蓿植株發病率呈正相關,皮爾遜相關性為r=0.806,P=0.000< 0.001。
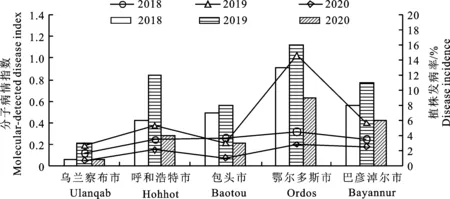
圖6 苜蓿銹菌越冬銹菌量與返青后植株發病率比較
3 結論與討論
Real-time qPCR技術是一種具有高度靈敏性和準確性的核酸定量監測技術。它是利用已知濃度的標準樣品構建標準曲線,定量獲得未知樣品的Ct值,再通過標準曲線上計算出未知樣品的起始量[17]。該方法被廣泛應用于動植物真菌、病毒、細菌等微生物的RNA和DNA檢測。Ma等[18]利用RNA水平的real-time qPCR技術建立檢測田間越冬小麥條銹菌初始菌量的體系。鄭亞明[19]和閆佳會等[16]分別基于DNA水平的qPCR技術對田間潛伏侵染的小麥白粉菌量和條銹菌量進行檢測。Dhar等[20]利用qPCR對空氣中霜霉病菌的孢子量進行實時監測,確定了病害防控閾值(8.55個/m3)。本研究建立的RNA水平的qPCR技術可用于定量檢測苜蓿銹菌越冬的初始菌量,能在田間苜蓿見病前2個月檢測到活體苜蓿菌量,結合本研究結果,苜蓿銹菌越冬菌量與返青后苜蓿植株發病率呈正相關,說明建立苜蓿銹菌越冬菌量qPCR檢測技術的必要性,為確定病害發生的重點區域及科學防控方法提供理論依據。
苜蓿銹菌是嚴格的專性寄生真菌,苜蓿銹病的分布與苜蓿種植區基本一致。1995年,侯天爵等[7]初步探究內蒙古苜蓿銹病的發生和分布范圍,其中,呼和浩特市和赤峰市(北緯約40°~41°)發病較為嚴重,錫林浩特市(北緯約44°)發病較輕,并提出苜蓿銹病的發生隨緯度升高而病害減輕的觀點。本研究的5個調查點中,烏蘭察布市的緯度(北緯約41.56°)最高,鄂爾多斯市的緯度(北緯約39.82°)最低,結合Real-time qPCR定量結果,烏蘭察布市的越冬苜蓿銹菌量最低,鄂爾多斯市的最高,結果與前人表述一致。但包頭市、巴彥淖爾市、呼和浩特市的越冬苜蓿銹菌量分布與地理緯度關系不明顯。病原菌的越冬菌量與品種抗凍性、溫度和濕度等相關氣象因子、秋季發病程度、越冬部位、種植環境等因素密切相關,單從地理緯度區分意義不大,仍需要通過熒光定量PCR技術才能獲得最準確可靠的越冬菌量。
本研究在返青后苜蓿植株上部5 cm材料中檢測到苜蓿銹菌,說明苜蓿銹菌可以在苜蓿地下根莖組織中順利越冬。這與侯天爵提出的苜蓿銹菌可以直接在苜蓿莖內越冬的觀點一致[3]。
溫度是影響苜蓿銹菌越冬的關鍵氣象因子。2019年的越冬苜蓿銹菌量顯著高于2018和2020年的。結合氣象資料分析,除包頭市外的4個調查點在2019年11月中旬到翌年1月中旬的日平均溫度分別為烏蘭察布市(-11.37 ℃)、呼和浩特市(-8.63 ℃)、鄂爾多斯市(-6.91 ℃)、巴彥淖爾市(-7.47 ℃),均高于同一地區在2018年和2020年該監測段的日平均溫度。包頭市在2018年的越冬銹菌量達到3 a中最大值,包頭市2018年該監測段的日平均溫度為2.55 ℃,高于2019年(-10.02 ℃)和2020年(-15.61 ℃)的日平均溫度。本研究顯示11月中旬到翌年1月中旬的日平均溫度與越冬苜蓿銹菌量呈正相關,溫度對銹菌越冬的影響仍需進一步研究證明。
關于苜蓿銹菌越冬的具體溫度研究,目前未見報道。但與其同屬于銹菌目柄銹菌科的小麥條銹菌(Pucciniastriiformisf.sp.tritici)的越冬溫度報道較多,如當翌年1月的平均氣溫低至 -6 ℃~-7 ℃時,小麥條銹菌越冬困難[21-25],但當有積雪覆蓋時,空氣溫度下降至-10 ℃小麥條銹菌菌絲體仍能順利越冬[21, 26-27]。Ma等[18]的研究表明,田間小麥條銹菌越冬存活率隨著溫度的降低而降低。室內試驗證明,小麥條銹菌在溫度 -20 ℃~-15 ℃的條件下處理12 h后仍存活,但存活率低于-10 ℃及以上溫度的銹菌存活率[28]。本研究統計了各調查點2020年11月中旬到翌年1月中旬的日平均溫度≤-10 ℃的積累時間,烏蘭察布市(71 d)、呼和浩特市(51 d)、包頭市(70 d)、鄂爾多斯市(37 d)、烏蘭察布市(58 d)。結合real-time qPCR定量結果分析,2020年烏蘭察布市和包頭市的越冬苜蓿銹菌量顯著低于其他地區。本研究顯示,低溫積累時間也是影響苜蓿銹菌越冬的重要因子,低溫積累時間越長,越冬銹菌量越少,但該結論還需要通過試驗進一步研究證明。
