布魯氏菌DnaJ蛋白的表達及功能分析
2022-04-27 09:27:06印雙紅張俊波李寅翠張孟琴蔡春連李志強陳創夫
中國獸醫雜志 2022年1期
關鍵詞:小鼠
印雙紅 , 張俊波 , 李寅翠 , 張孟琴 , 毛 宏 , 易 萌 , 蔡春連 , 張 紅 , 李志強 , 陳創夫
(1.銅仁學院大健康學院 , 貴州 銅仁 554300 ; 2.銅仁學院農林工程與規劃學院 , 貴州 銅仁 554300 ;3.銅仁學院貴州省梵凈山地區生物多樣性保護與利用重點實驗室 , 貴州 銅仁 554300 ; 4.銅仁學院材料與化學工程學院 , 貴州 銅仁 554300 ; 5.商丘師范學院生物與食品學院 , 河南 商丘 476000 ; 6.石河子大學動物科技學院 , 新疆 石河子 832000)
布魯氏菌病是一種人獸共患傳染病,布魯氏菌疫苗仍存在安全性差、免疫期短、重復免疫等一些缺陷[1],現有的亞單位疫苗還未達到減毒活疫苗的保護水平[2],與減毒活疫苗相比,亞單位疫苗具有避免發生免疫副反應并有助于血清學監測的優點[3],用于布魯氏菌病亞單位疫苗研制的蛋白主要包括分子伴侶DnaK、外膜蛋白和AsnC等[4-5],但這些融合蛋白疫苗不能持續誘導有效的細胞免疫反應。
DnaJ是一種具有分子伴侶特性的蛋白質,DnaJ蛋白在生命體細胞中具有活性,功能為糾正或清除細胞內錯誤折疊、損傷失去活性的蛋白質和指導生成正確且具有生物活性的蛋白質,從而保護細胞免受應激破壞[6]。本試驗通過對DnaJ蛋白進行表達與純化,并研究其在細胞和小鼠體內產生的細胞和體液免疫反應,旨在為該蛋白的功能和疫苗開發研究奠定基礎。
1 材料與方法
1.1 主要試劑及實驗動物TaqDNA聚合酶、T4 DNA連接酶、蛋白Marker和DNA Marker,均購自寶生物工程(大連)有限公司;質粒pET-30a,購自Novagen公司;小鼠IFN-γ、IL-4、IgG1和IgG2a ELISA試劑盒,均購自美國GBD公司;瓊脂糖凝膠回收試劑盒和質粒提取試劑盒,均購自天根生化科技(北京)有限公司。
1.2 菌株和細胞 布魯氏菌M5-90、DE3感受態細胞、DH5α感受態細胞和RAW264.7細胞為貴州省梵凈山地區生物多樣性保護與利用重點實驗室保存。
1.3 實驗動物 6周齡體重約17 g的BALB/c小鼠,購自新疆醫科大學動物實驗中心。
1.4 試驗方法
1.4.1 DnaJ蛋白的生物信息學分析 利用Predicting Antigenic Peptides在線預測DnaJ蛋白的抗原決定簇。使用SOPMA在線軟件分析預測DnaJ蛋白的二級結構。
1.4.2DnaJ基因的合成及重組表達質粒pET-30a-DnaJ的構建 在布魯氏菌M5-90株的DnaJ基因序列(BMEI1513)上接上NdeI/Hind III酶切位點,然后送通用生物系統(安徽)有限公司合成基因序列。對pET-30a表達載體進行NdeI/Hind III雙酶切,然后將DnaJ(NdeI/Hind III)目的片段連接至pET-30a載體,連接體系為20 μL:pET-30a 4.0 μL,10×Ligation Buffer 4.0 μL,T4 DNA Ligase 0.3 μL,ddH2O 9.7 μL。22 ℃連接16 h。用試劑盒提取pET-30a-DnaJ質粒,然后對該質粒進行酶切驗證,酶切體系為30 μL:NdeI內切酶1.0 μL,Hind III內切酶1.0 μL,質粒pET-30a-DnaJ 4.0 μL,10×Ligation Buffer 3.0 μL,ddH2O 21.0 μL,然后將質粒送深圳華大基因科技有限公司測序,然后進行DnaJ蛋白表達。
1.4.3 DnaJ蛋白的表達及純化 將PET-30a-DnaJ質粒轉化至DE3感受態細胞中,37 ℃、220 r/min振搖至菌體OD600值為0.6~0.8,向細菌培養物中加入誘導劑IPTG(1 mmol/L),在37 ℃、220 r/min條件下培養2、4 h和6 h以誘導DnaJ蛋白的表達,用20 μL的PBS重懸菌體并加入20 μL的5×Loading Buffer,利用12%的SDS-PAGE凝膠電泳檢測,凝膠經染色和脫色后檢測DnaJ蛋白的表達情況。將培養好的菌液12 000 r/min離心6 min,加入裂解液Liysis 4 ℃裂解10 h,在液氮和37 ℃水浴鍋中反復凍融3次后,對菌體進行超聲破碎,12 000 r/min離心25 min后收集沉淀,加入8 mol/L尿素,用0.45 μm濾膜過濾,用AKTAxpress智能多維純化系統對DnaJ蛋白純化,收集洗脫液,SDS-PAGE電泳檢測DnaJ蛋白的純化效果。
1.4.4 小鼠巨噬細胞RAW 264.7中的IFN-γ和IL-4檢測 用含10%胎牛血清的培養基在37 ℃、5%CO2條件下培養RAW264.7細胞至對數生長期,試驗組每孔加入25 mg的DnaJ蛋白,對照組添加PBS,在4、12 h和24 h收集細胞上清液,用ELISA試劑盒測定IFN-γ和IL-4的水平,操作方法見試劑盒說明書。
1.4.5 小鼠脾細胞中的IFN-γ和IL-4檢測 選取6周齡的BALB/c小鼠,將正常小鼠斷頸椎處死(每組5只),無菌取出脾臟,通過勻漿法獲得單細胞懸液,利用ACK紅細胞裂解液除去紅細胞。脾細胞在96孔微量滴定板中以4×105個細胞/孔進行培養;試驗組每孔加入25 mg的DnaJ蛋白,對照組添加PBS。在37 ℃、5%CO2條件下與細胞共同孵育12 h和24 h,收集細胞上清液,用ELISA試劑盒檢測脾細胞分泌的IFN-γ和IL-4水平,操作步驟見試劑盒使用說明書。
1.4.6 小鼠免疫及體液免疫水平檢測 將6周齡的BALB/c小鼠隨機分為試驗組和對照組,20只/組。試驗組小鼠腹腔注射DnaJ蛋白(50 μg/只),對照組小鼠注射PBS(200 μL/只),共免疫2次,免疫間隔21 d,首次免疫后的第7、21天和第35天對小鼠斷尾采血分離血清,用ELISA試劑盒測定血清中IgG1和IgG2a水平,操作步驟見試劑盒使用說明書。
1.4.7 DnaJ蛋白免疫小鼠后誘導IFN-γ和IL-4水平分析 取1.4.6中免疫35 d的小鼠,將小鼠斷頸椎處死(每組5只),無菌取出脾臟,通過勻漿法獲得單細胞懸液,利用ACK紅細胞裂解液除去紅細胞。脾細胞在96孔微量滴定板中以4×105個/孔進行培養;每孔加入25 mg的DnaJ蛋白或熱滅活的布魯氏菌16M裂解液,陽性對照組添加0.5 mg的ConA,陰性對照組添加RPMI 1640細胞培養基,在37 ℃、5%CO2條件下與細胞共同孵育72 h,收集細胞上清液,用ELISA試劑盒測定細胞中IFN-γ和IL-4的水平,操作方法見試劑盒說明書。

2 結果
2.1 DnaJ重組蛋白的生物信息學分析 Predicting Antigenic Peptides預測DnaJ蛋白有11個抗原決定簇,分別在第4~12、21~30、46~53、71~78、149~172、178~188、209~217、227~245、254~262、265~271、285~293位氨基酸處(表1)。SOPMA預測DnaJ蛋白中有68個氨基酸參與形成α-螺旋,占總氨基酸的比例為21.73%;54個氨基酸參與延伸鏈的形成,占總氨基酸的比例為17.25%;21個氨基酸參與β-折疊的形成,占總氨基酸的比例為6.71%;170個氨基酸參與無規卷曲結構的形成,占總氨基酸的比例為54.31%(圖1)。

表1 DnaJ蛋白抗原決定簇的預測Table 1 Prediction of antigen clusters in DnaJ protein

圖1 DnaJ蛋白二級結構的預測Fig.1 Prediction of secondary structure in DnaJ protein
2.2 重組表達質粒pET-30a-DnaJ的構建及鑒定 對重組載體pET-30a-DnaJ進行NdeI/Hind III雙酶切,得到942 bp的條帶,與理論值相符(圖2),同時測序結果與GenBank中一致,表明重組質粒pET-30a-DnaJ構建成功。
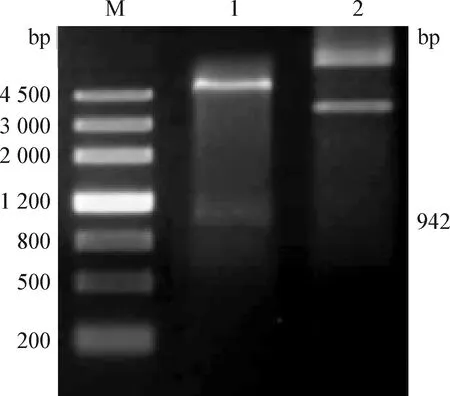
圖2 重組質粒pET-30a-DnaJ的構建及鑒定Fig. 2 Construction and verification of recombinant plasmid pET-30a-DnaJM:DNA marker 4 500; 1:pET-30a-DnaJ的酶切產物; 2:pET-30a-DnaJ質粒M:DNA marker 4 500; 1:pET-30a-DnaJ digestion product;2:pET-30a-DnaJ plasmid
2.3 DnaJ蛋白的表達及純化 表達結果顯示,得到與預期蛋白質分子質量大小一致的37.7 kDa DnaJ蛋白(圖3A)。純化結果顯示,DnaJ蛋白的純化效果較好(圖3B)。
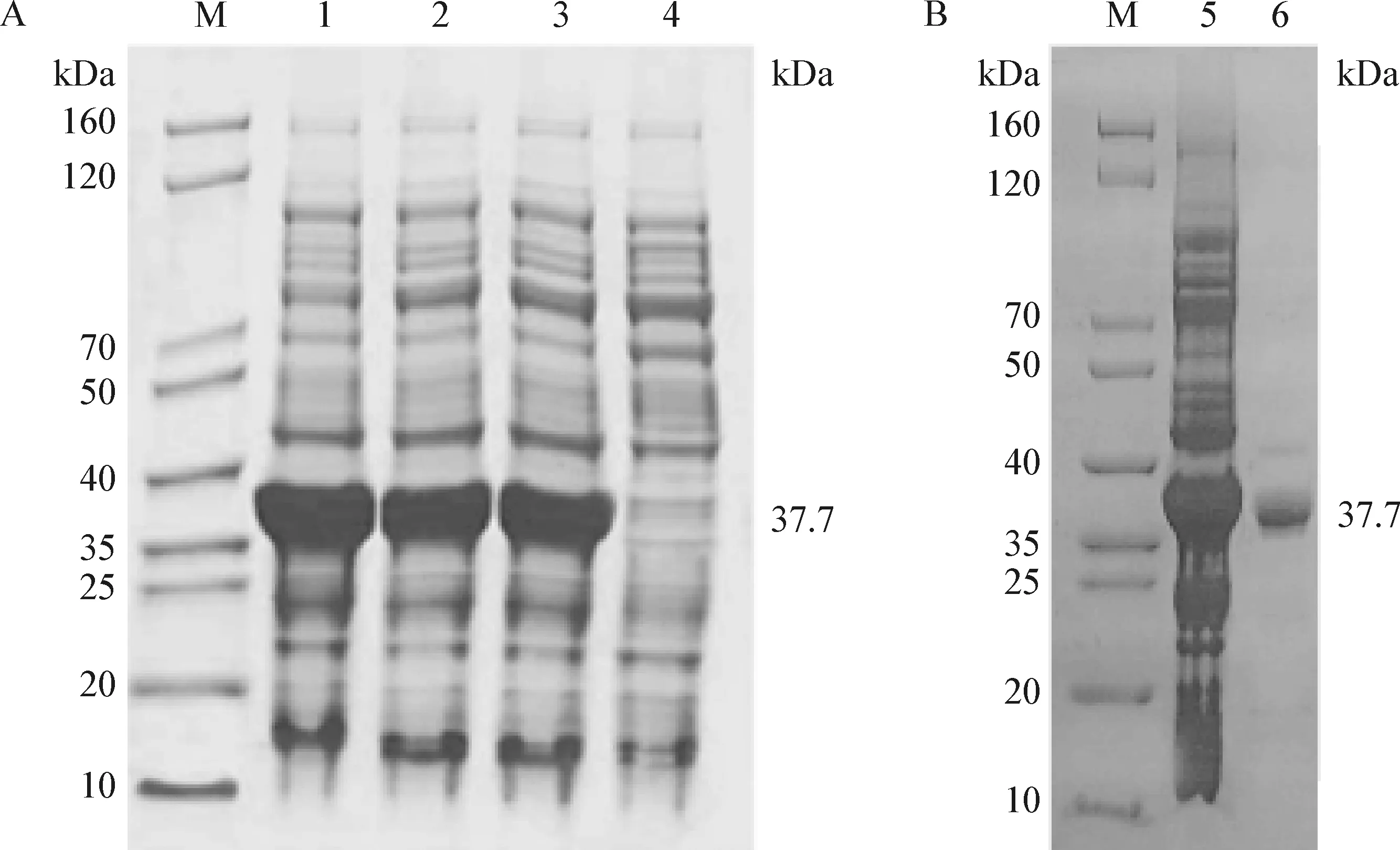
圖3 融合蛋白DnaJ的表達鑒定(A)及純化(B)Fig. 3 Expression,identification (A) and purification (B) of fusion protein DnaJM:蛋白分子marker; 1:IPTG誘導2 h的pET-30a-DnaJ; 2:IPTG誘導4 h的pET-30a-DnaJ; 3:IPTG誘導6 h的pET-30a-DnaJ; 4:DE3; 5:未純化蛋白; 6:純化蛋白M:Protein molecule marker; 1:Induced pET-30a-DnaJ for 2 h by IPTG; 2:Induced pET-30a-DnaJ for 4 h by IPTG; 3:Induced pET-30a-DnaJ for 6 h by IPTG; 4:DE3; 5:Unpurified protein; 6:Purified protein
2.4 DnaJ蛋白誘導RAW 264.7細胞產生IFN-γ和IL-4 結果如圖4所示,在4 h時,DnaJ蛋白刺激RAW 264.7細胞中的IFN-γ水平顯著高于PBS對照組(P<0.05),而IL-4水平無明顯變化(P>0.05);在12 h和24 h時,DnaJ蛋白刺激RAW 264.7細胞中的IFN-γ和IL-4水平均極顯著高于PBS對照組(P<0.01)。結果表明,DnaJ蛋白可誘導RAW 264.7細胞產生IFN-γ和IL-4。
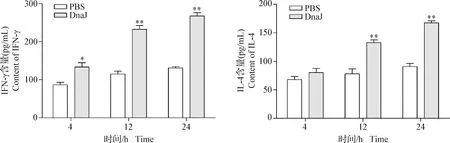
圖4 DnaJ融合蛋白誘導RAW 264.7細胞產生的IFN-γ和IL-4水平變化Fig.4 The levels of IFN-γ and IL-4 cytokine production in RAW 264.7 cells after induction with DnaJ fusion protein與PBS對照組相比,*:P<0.05,**:P<0.01; 下圖同Compare to group PBS, *:P<0.05,**:P<0.01. The same as below
2.5 DnaJ蛋白誘導小鼠脾細胞產生IFN-γ和IL-4 結果如圖5所示,在12 h和24 h時,DnaJ蛋白刺激小鼠脾細胞產生的IFN-γ和IL-4水平均極顯著高于對照組(P<0.01)。結果表明,DnaJ蛋白可誘導小鼠脾細胞產生Th1和Th2型免疫反應。
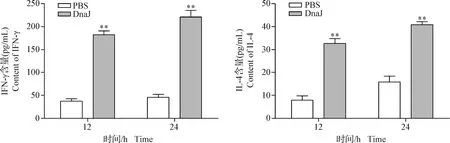
圖5 DnaJ融合蛋白誘導小鼠脾細胞產生的IFN-γ和IL-4水平變化Fig.5 The levels of IFN-γ and IL-4 production in mouse splenocytes after induction with DnaJ fusion protein
2.6 DnaJ蛋白可誘導細胞免疫反應 結果如圖6所示,免疫DnaJ蛋白的小鼠脾細胞產生的IFN-γ和IL-4水平均極顯著高于免疫PBS的小鼠(P<0.01);ConA可誘導小鼠脾細胞產生高水平的IFN-γ和IL-4;而RPMI 1640和免疫PBS不能誘導小鼠脾細胞產生IFN-γ和IL-4。結果表明,DnaJ蛋白可誘導小鼠產生Th1和Th2型免疫反應。
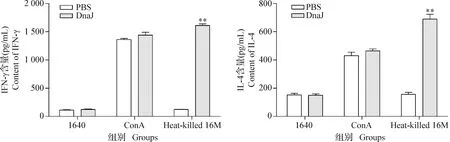
圖6 DnaJ融合蛋白誘導的細胞免疫反應變化Fig. 6 Changes in cellular immune response induction with DnaJ fusion protein
2.7 DnaJ蛋白可誘導體液免疫反應 結果如圖7所示,在7 d時,免疫DnaJ蛋白的小鼠產生的IgG1和IgG2a與PBS對照組相比無明顯變化(P>0.05);在21 d和35 d時,免疫DnaJ蛋白的小鼠產生的IgG1和IgG2a水平極顯著高于PBS對照組(P<0.01)。結果表明,DnaJ蛋白免疫小鼠可誘導機體產生IgG1和IgG2a的體液免疫反應。
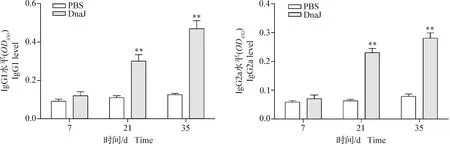
圖7 DnaJ融合蛋白誘導的體液免疫反應變化Fig. 7 Changes in humoral immune response induction with DnaJ fusion protein
3 討論
Th1型免疫反應的主要特性是產生IFN-γ,以抵抗布魯氏菌的感染[7]。巨噬細胞在布魯氏菌感染過程中可分泌大量IFN-γ以殺死胞內的細菌[8]。IL-4由Th2型細胞產生,可介導細胞免疫應答,誘導Th2細胞的生長和分化[9]。IgG1抗體的產生依賴于Th2型細胞因子IL-4的分泌,而IgG2a抗體的產生依賴于Th1型細胞因子IFN-γ的分泌[10]。本試驗發現,DnaJ融合蛋白可刺激小鼠巨噬細胞、脾細胞及小鼠體內均產生高水平的IFN-γ和IL-4,且在小鼠體內誘導高水平的IgG1和IgG2a抗體,表明DnaJ蛋白可誘導小鼠體外細胞和體內產生Th1和Th2型免疫反應,且Th1和Th2型免疫反應進一步促進了IgG1和IgG2a抗體產生,增加了小鼠的體液免疫。
DnaJ是刺激DnaK的ATP酶和蛋白折疊活性的輔助分子伴侶[11]。DnaJ作為毒力因子參與一些細菌的致病過程,影響細菌感染宿主細胞[12-13]。黏膜免疫的DnaJ蛋白可誘導細胞產生抗體、IL-10、IFN-γ和IL-17A,通過腹膜內接種DnaJ后誘導小鼠IgG滴度和淋巴細胞增殖;黏膜免疫接種DnaJ蛋白能夠降低鼻或肺定植的肺炎球菌,并抵御不同種血清型肺炎球菌的感染,腹腔內接種DnaJ蛋白也能夠抵御不同種血清型肺炎球菌的感染,DnaJ是肺炎球菌具潛力的候選蛋白疫苗[14]。本試驗發現,布魯氏菌DnaJ蛋白可誘導較好的細胞和體液免疫,因此,該蛋白可作為候選的亞單位疫苗。
熱休克蛋白的誘導合成對于致病菌在宿主體內存活至關重要[15]。肺炎鏈球菌DnaJ蛋白具有良好的免疫原性,且與其感染致病力相關[16]。研究表明,DnaJ在遲緩愛德華氏菌的致病中發揮重要作用,且具有免疫保護作用,因此DnaJ可用于控制水產養殖中的遲緩愛德華氏菌感染[17]。布魯氏菌的應激反應蛋白毒力因子包括Hfq、DnaK、DnaJ、HtrA和Lon。研究表明,Hfq在布魯氏菌致病中可調控大量靶標基因的表達水平,影響布魯氏菌在宿主體內的慢性持續性感染[18-19]。然而,目前只初步證明Lon、htrA、DnaJ和DnaK影響布魯氏菌的毒力[20-23],它們在布魯氏菌致病中的作用機制尚不清楚。本課題組前期已經獲得了可作為候選疫苗株的布魯氏菌Hfq和DnaK基因缺失株,因此后期也將進一步構建DnaJ基因缺失株,獲得DnaJ基因調控布魯氏菌的毒力機制。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
