退化高寒草甸狼毒群落分類特征及其環境影響因子
2022-04-15 12:06:50劉詠梅董幸枝龍永清朱志梅王雷蓋星華趙樊李京忠
草業學報 2022年4期
劉詠梅,董幸枝,龍永清,2,朱志梅,2,王雷,2,蓋星華,趙樊,李京忠
(1.西北大學城市與環境學院,陜西 西安 710127;2.陜西省地表系統與環境承載力重點實驗室,陜西 西安 710127;3.中科星圖空間技術有限公司,陜西 西安 710100;4.許昌學院城鄉規劃與園林學院,河南 許昌 461000)
高寒草甸約占青藏高原高寒草地面積的近1/2[1-2],是最具代表性的高寒草地生態系統之一,發揮著調節氣候、涵養水源和維護生態平衡等多項重要的生態功能[3]。在氣候變化和人類活動的影響下,高寒草甸的生物多樣性減少、生產力下降、毒雜草化趨勢明顯[4-5]。隨著草地退化加劇,高寒草甸的物種構成發生改變,優良牧草相對豐度下降,毒雜草種類和數量快速增長[5]。狼毒(Stellera chamaejasme)是青藏高原危害最嚴重的有毒植物之一,狼毒的出現是草地植物群落長期逆向演替的結果,已成為草地重度退化階段的標志性植物[6-7]。狼毒從20世紀60年代開始在青藏高原高寒草地入侵擴散,密度逐年提高。2011年調查顯示,青海省海北州祁連縣僅狼毒入侵嚴重的峨堡鎮、默勒鎮、阿柔鄉及野牛溝鄉等地,受影響草甸面積達到9.033×104hm2[8],近年來其分布區域仍在迅速擴大。狼毒在青藏高原草地核心區域呈大面積快速蔓延的趨勢,對高寒草甸生態系統演變的深刻影響和當地畜牧業健康發展的嚴重制約日益顯現。
狼毒是瑞香科(Thymelaeaceae)狼毒屬(Stellera)多年生草本植物,全株有毒,廣泛分布于海拔2300~4200 m、年均溫約0℃的高山及亞高山草地。狼毒為草地群落伴生種,在群落物種競爭中逐漸替代原生嵩草屬種類而成為中重度退化草甸的主要優勢種[6]。近年來,多位學者在狼毒種群空間分布格局及控制機理[9-10]、種內種間競爭與空間關聯[11]、葉、枝生長對地形因子的響應[12-13]、狼毒種子對其他植物種子及幼苗的化感影響[14],以及狼毒入侵草地土壤水分特征及空間變異[8]等方面開展了深入的研究工作。結果顯示,狼毒具有較強的高寒環境適應能力,通過優化資源利用、減弱種內競爭及影響其他植物生長等在種間競爭過程中占據明顯優勢,種群分布迅速增長。現有研究為深入認識狼毒的擴散機制提供了科學參考,然而對于退化高寒草甸狼毒群落的分類特征及其對環境因素的響應研究未見報道。數量分類采用數學方法研究植物群落分布的間斷性,排序則研究植被的連續變化,兩者結合為科學揭示植被-環境的生態關系提供了重要的分析手段[15-16]。植物群落的環境影響因子與空間尺度密切關聯[17],大尺度研究反映了植被空間格局與氣候因素之間的關系,小尺度研究則能更加精細深入地揭示土壤、地形及水文條件對植物群落空間分布的影響[18]。鑒于此,本研究基于祁連山中段典型狼毒入侵草甸的樣方調查、土壤采樣和地形測量數據,運用雙向指示種分析(two-way indicator species analysis,TWINSPAN)、除趨勢對應分析(detrended correspondence analysis,DCA)和冗余分析(redundancy analysis,RDA)在樣地尺度上分析退化高寒草甸狼毒群落的物種構成和群叢類型,探討狼毒群落的環境梯度變化規律和主要影響因素,旨在揭示狼毒入侵草甸的退化演替趨勢,闡明微觀環境中狼毒群落分布的驅動機制,為祁連山地區狼毒型退化草甸的恢復重建、毒雜草的綜合治理和畜牧業的可持續發展提供科學參考和依據。
1 材料與方法
1.1 研究區概況
研究區位于祁連山中段的祁連縣境內,屬于大陸性高寒山區氣候,年平均日照時間為2852.45 h[19],氣溫日較差大,年平均氣溫1℃,年平均降水量約412 mm[8],土壤類型為高山草甸土,草地類型主要為高寒草甸。典型樣地選在八寶鎮青陽溝內,地理位置100°21′38.207″-100°21′51.610″E、38°9′31.760″-38°9′39.721″N,面積約4.05×104m2,地勢開闊略有起伏,平均海拔3073 m,平均坡度6.3°。區域內為典型的狼毒入侵退化草甸,禾本科植物以紫花針茅(Stipa purpurea)、早熟禾(Poa annua)為優勢種,莎草科牧草以線葉嵩草(Kobresia capillifolia)和矮生嵩草(Kobresia humilis)為主,豆科植物主要有花苜蓿(Medicago ruthenica)和異葉米口袋(Gueldenstaedtia diversifolia);毒雜草種類較多,主要有狼毒、草玉梅(Anemone rivularis)、披針葉黃華(Thermopsis lanceolata)、乳白 香 青(Anaphalis lactea)、青 海 刺 參(Morina kokonorica)、高 山唐松 草(Thalictrum alpinum)、高 原 毛 茛(Ranunculus tanguticus)、秦 艽(Gentiana macrophylla)、麻 花 艽(Gentiana straminea)和 蒲 公 英(Taraxacum mongolicum)等,約占群落物種總數的1/3。群落蓋度26.0%~63.0%,狼毒平均蓋度15.4%,平均斑塊密度34株·m-2,呈斑塊狀聚集的空間分布特征。
1.2 群落調查
群落調查于2019年7月上中旬狼毒盛花期完成,綜合考慮狼毒群落覆蓋及區域地形特征,在樣地內布設3條樣線,間隔45 m,沿樣線布設1 m×1 m樣方15個,間隔30 m,共計45個樣方(圖1)。記錄每個樣方內出現的植物種類、測量各物種的高度和密度,采用數碼相機對樣方垂直向下拍照獲取蓋度照片,在ArcGIS軟件中對照片進行裁剪和校正,解譯樣方總蓋度和物種分蓋度,同時利用GNSS RTK測定樣方的經緯度坐標和高程。在地面調查的同時采用大疆精靈4 RTK無人機對研究樣地進行低空航拍(航高75 m),獲取2 cm數字正射影像圖(digital orthophoto map,DOM)與數字表面模型(digital surface model,DSM)[20],通過人工編輯消除植被影響,重采樣生成50 cm數字高程模型(digital elevation model,DEM)。在ArcGIS軟件中基于DEM生成坡度和坡向兩個專題層,提取每個樣方的坡度與坡向值。其中,坡向定義為朝東為起點,按順時針方向旋轉每45°劃分為一個等級,以1~8依次代表北、東北、西北、東、西、東南、西南及南8個坡向,數字越大越向陽干熱[21]。

圖1 調查樣方分布Fig.1 Quadrats distribution
在每個樣方中心用土鉆采集土壤樣品,采樣深度60 cm,采樣間隔10 cm,用鋁盒裝入樣品并稱取濕重用于測定每層的土壤含水量,等量混合6層樣品并裝入自封袋中用于測定土壤養分和pH值。土壤水分樣品帶回實驗室后采用烘干法測定土壤含水量(soil water content,SWC),混合樣品經過自然晾干研磨后分別采用1 mm和0.15 mm土壤篩進行處理。采用0.15 mm樣品測定土壤有機質(soil organic matter,SOM)、全氮(total nitrogen,TN)、全磷(total phosphorus,TP)及全鉀(total potassium,TK)含量,采用1 mm樣品測定土壤pH值。其中,采用重鉻酸鉀氧化-外加熱法測定土壤有機質含量,采用凱氏定氮法測定全氮含量,采用堿熔-鉬銻抗比色法測定全磷含量,采用堿熔-原子吸收分光光度法測定全鉀含量,采用電位測定法[V(水)∶m(土)=2.5∶1]測定土壤pH值[22]。
1.3 數據處理
采用重要值(important value,IV)作為物種的優勢度指標,評價各物種在群落中的功能和作用,其計算公式如下[15]:
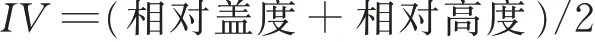
群落調查記錄研究區內植物35種,剔除出現頻次小于2的植物種后得到30種,建立45個樣方和30種植物重要值組成的植被數據矩陣(45×30),研究區內物種的重要值見表1。依據野外測量和實驗室測定數據,選取地形因子3個:海拔(altitude,AL)、坡度(slope,SL)、坡向(aspect,AS);土壤因子6個:SOM、TN、TP、TK、p H和SWC,建立45個樣方和9個環境因子組成的環境數據矩陣(45×9),環境因子數據見表2。

表1 退化草甸狼毒群落物種重要值Table 1 Important values of the species of vegetation communities in S.chamaejasme invaded alpine meadow
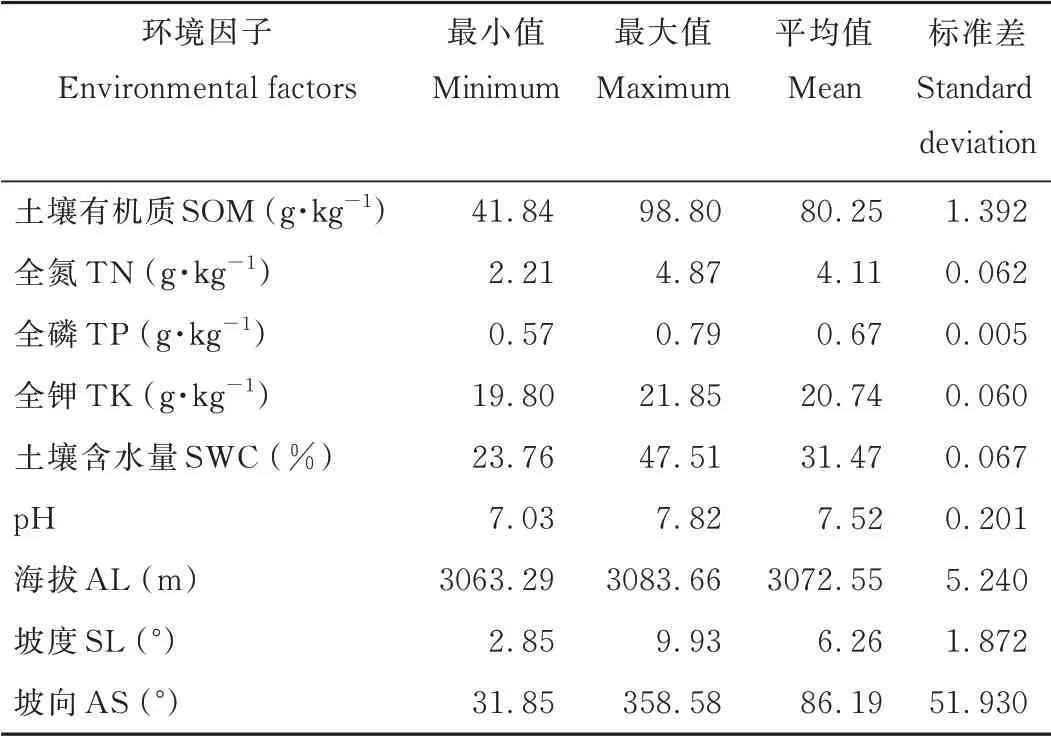
表2 研究區環境影響因子參數Table 2 Gener al sur vey of environmental par ameter s in the study area
應用雙向指示種分析(TWINSPAN)進行退化草甸狼毒群落分類。TWINSPAN采用二歧式分割法劃分群叢類型,在PC-ORD 5.0中運行程序時以樣方-物種重要值矩陣為基礎,將切割水平定為0、0.02、0.05、0.1和0.2,將研究區植物重要值劃分為3個等級。應用Canoco 5.0軟件對樣方-物種重要值矩陣進行除趨勢對應分析,根據DCA排序軸的梯度長度選擇適宜的排序方法[23]。研究區45個樣方-物種重要值矩陣的DCA排序顯示,4個排序軸梯度長度均小于3,最大值為1.49,因此選擇基于線性模型的冗余分析(RDA)進行樣方和環境變量的排序,應用蒙特卡洛置換檢驗(Monte Carlo permutation test,模擬999次)和前向選擇法分析環境因子對物種分布影響的顯著性。依據蒙特卡羅檢驗結果可以接受所選環境因子整體對高寒草甸狼毒群落分布的解釋量(P=0.002)。在此基礎上利用Canoco 5.0軟件的自動前向選擇程序進一步檢測單個環境因子影響物種分布的顯著性,從9個環境因子中選出對狼毒群落物種分布影響最重要的5個環境因子:TP、TK、SOM、AL和AS(P<0.05),作為RDA分析的環境變量。
2 結果與分析
2.1 狼毒群落TWINSPAN分類
對研究區45個樣方進行TWINSPAN分類,結合實地調查結果,采用第2級分類結果將退化草甸狼毒群落的45個樣方劃分為3個群叢(圖2)。
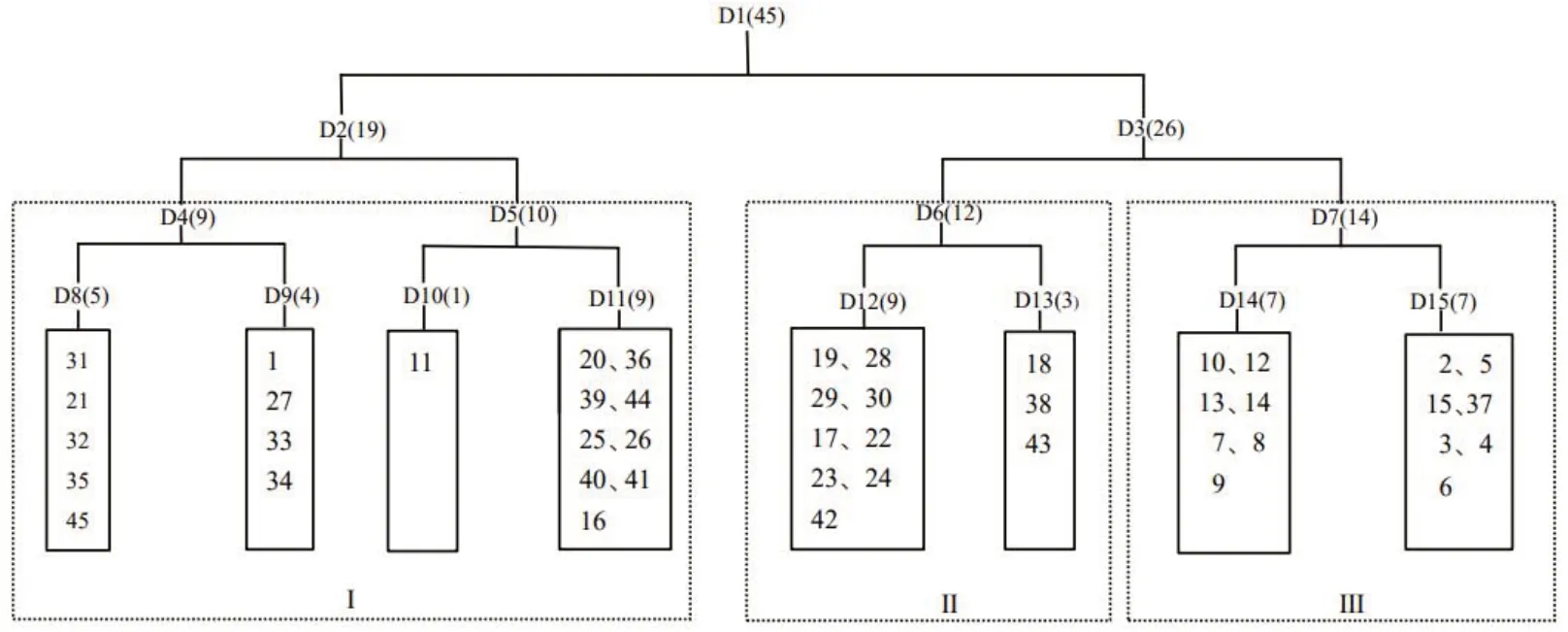
圖2 退化草甸狼毒群落TWINSPAN分類樹狀圖Fig.2 Dendrogram of the TWINSPAN classification of 45 plots in S.chamaejasme invaded alpine meadow
群叢Ⅰ:狼毒+紫花針茅+線葉嵩草+披針葉黃華,伴生種有青海刺參、早熟禾、委陵菜、薄荷、柔軟紫菀、高原毛茛等,包括1、11、16、20、21、25~27、31~36、39~41、44、45,共19個樣方。群落主要分布于西南、東南和南坡,海拔3066~3084 m,坡度3.5°~8.6°,土壤含水量24.4%~44.5%,群落總蓋度40.4%~63.0%,狼毒平均蓋度29.7%。禾本科植物重要值2.10,豆科0.25,莎草科2.19;毒雜草類10.08,狼毒5.64。
群叢Ⅱ:狼毒+紫花針茅+委陵菜+美麗風毛菊,伴生種有早熟禾、乳白香青、披針葉黃華、矮生嵩草、花苜蓿、青海刺參等,包括17~19、22~24、28~30、38、42~43,共12個樣方。群落主要分布于東南、西南和南坡,海拔3066~3079 m,坡度3.2°~8.8°,土壤含水量23.8%~44.6%。群落總蓋度36.6%~59.4%,狼毒平均蓋度11.0%。禾本科植物重要值3.12,豆科0.57,莎草科1.18;毒雜草類5.14,狼毒1.82。
群叢Ⅲ:紫花針茅+草玉梅+狼毒+早熟禾,伴生種有委陵菜、乳白香青、美麗風毛菊、線葉嵩草、矮生嵩草、高山唐松草等,包括2~10、12~15、37,共14個樣方。群落生長范圍主要涉及東南和南坡,海拔3063~3077 m,坡度2.9°~10.0°,土壤含水量25.4%~47.5%。群落總蓋度26.0%~48.6%,狼毒平均蓋度5.5%。禾本科植物重要值4.10,豆科0.28,莎草科0.23;毒雜草類6.10,狼毒1.70。
2.2 DCA排序分析
采用DCA對研究區內45個樣方的物種重要值數據進行排序分析,前4個排序軸的特征值分別為0.1560、0.0825、0.0437、0.0310,其中前兩個排序軸的特征值較大,所含信息量占4個排序軸總信息量的76.1%,生態意義顯著,因此采用前兩軸對樣方進行排序分析(圖3)。結果顯示,TWINSPAN劃分的3個群叢類型在二維排序圖上呈聚集分布,每個群叢的界限明確。DCA排序結果與TWINSPAN分類結果基本一致,準確反映了退化草甸狼毒群落的環境梯度變化,同時進一步驗證了TWINSPAN分類結果的合理性。
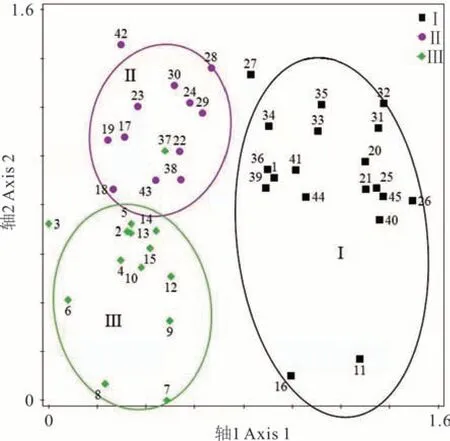
圖3 退化草甸狼毒群落樣方DCA排序Fig.3 DCA two-dimensional or dination for plots in S.chamaejasme invaded alpine meadow
2.3 群落分布-環境因子關系
對研究區45個樣方和選出的5個環境因子進行RDA排序,探討狼毒群落分布與環境因子之間的關系。RDA前4個排序軸的特征值分別為0.3952、0.0612、0.0328、0.0203,前兩軸包含了絕大部分信息,占總信息量的89.6%。第1軸和第2軸分別解釋了39.52%和6.12%的物種變異,與環境因子的相關系數分別達到0.88和0.74。RDA前4軸對物種-環境關系的累積解釋量為93.71%,其中前兩軸的累積解釋量達到83.94%,表明排序效果良好(表3)。第1軸和第2軸組成的雙序圖直觀表達了退化高寒草甸狼毒群落的主要環境特征,圖中箭頭表示環境因子,箭頭所處象限表示環境因子與排序軸間的正負相關性,箭頭連線長度表示環境因子與群落分布和物種分布之間相關性的大小,連線越長,相關性越大;箭頭連線與排序軸夾角則表示環境因子與排序軸相關性的大小,夾角越小,相關性越大(圖4)。
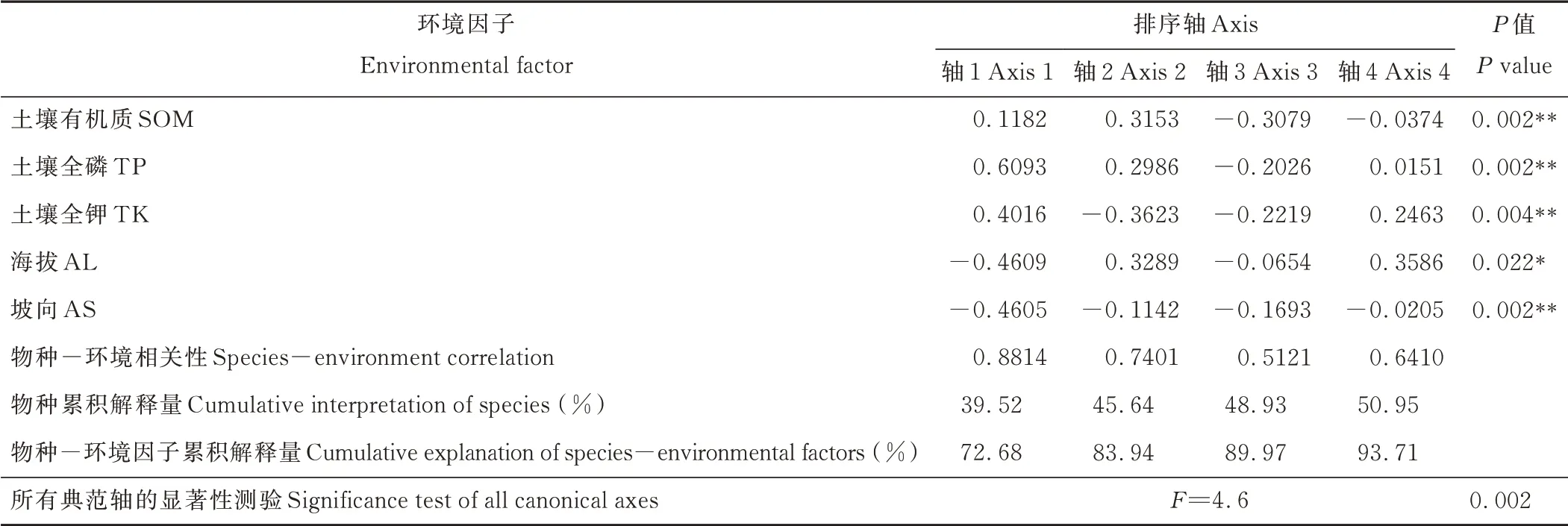
表3 退化草甸狼毒群落樣方-環境因子的RDA排序Table 3 RDA analysis of plots and envir onmental factor s in S.chamaejasme invaded alpine meadow
RDA第1排序軸與土壤全磷、全鉀及有機質含量呈正相關(表3和圖4),與海拔和坡向呈負相關,相關系數絕對值大小依次為全磷>海拔>坡向>全鉀>土壤有機質,坡向與第1軸夾角最小,但全磷的連線長度明顯大于坡向的長度,考慮相關系數、夾角和連線長度可以看出,第1軸主要反映了狼毒群落分布隨全磷和坡向的梯度變化,從左到右全磷含量增加,坡向趨于背陰。RDA第2排序軸分別與土壤全鉀含量、海拔及土壤有機質含量的負、正相關性最高,土壤有機質與第2軸的夾角最小,但全鉀的長度明顯大于土壤有機質的長度,因此第2軸主要體現了全鉀和土壤有機質的梯度變化,從下到上全鉀含量降低、土壤有機質逐漸增加。綜合來看,土壤全磷和全鉀含量是祁連山中段退化草甸狼毒群落分布最重要的影響因子,其次是坡向、土壤有機質和海拔。結合群落TWINSPAN分類結果(圖2)可知,群叢Ⅰ(狼毒平均蓋度29.7%)主要分布在土壤全磷、全鉀較低、向陽干燥的區域,位于排序圖的左方;群叢Ⅱ(狼毒平均蓋度11.0%)主要分布在全磷較低、全鉀低、有機質含量高、趨于向陽且海拔較高的區域,主要位于排序圖左上方;群叢Ⅲ(狼毒平均蓋度5.5%)生長在全磷、全鉀高、有機質含量低、相對背陰且海拔較低的環境中,主要位于排序圖右下方。RDA排序和TWINSPAN分類結果具有一致性,較好地反映了不同狼毒群叢類型的環境影響因子差異。

圖4 樣方RDA二維排序Fig.4 RDA two-dimensional ordination for plots in S.chamaejasme invaded alpine meadow
將環境因子對狼毒群叢分布格局的解釋能力進行定量分解(圖5)。土壤和地形兩類因子共同解釋了研究區狼毒群叢空間分布的47.5%,未能解釋的部分占52.5%。土壤因子的解釋能力為36.4%,其中24.5%由純土壤因子引起;地形因子的解釋能力為23.0%,其中11.1%由純地形因子引起。土壤和地形因子對狼毒群叢變化的耦合解釋能力為11.9%,分別占兩者解釋能力的32.7%和51.7%,顯示出土壤與地形因子對退化草甸狼毒群落分布具有較強的交互影響。
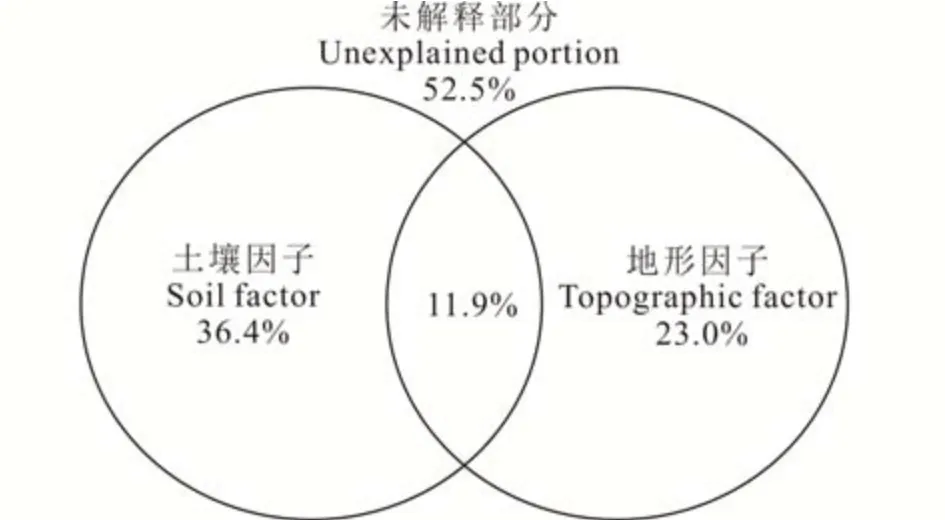
圖5 土壤和地形因子對狼毒群落分布的解釋能力Fig.5 Variation in S.chamaejasme community explained by soil and topographic factor s
2.4 物種分布-環境因子關系
對研究區30個物種進行RDA排序,進一步分析物種分布與環境因子的關系,物種在雙序圖內的位置體現了該物種的綜合生境特征。與樣方RDA排序相同,第1軸和第2軸主要反映了物種分布的土壤養分情況和地形條件,土壤全磷、全鉀及坡向是影響物種分布的主要環境變量。沿RDA第1軸從左向右,土壤全磷、全鉀含量逐漸增加,坡向趨于背陰,狼毒位于排序圖最左側,耐旱的生境特征明顯(圖6);紫花針茅、早熟禾分布在排序軸的偏中部及中部,顯示出其在該區域較強的生境適應能力,是狼毒群落的主要優勢種和伴生種,分布較為廣泛;草玉梅、圓穗蓼的生境相對濕潤,分布在排序軸右側。這些主要物種的分布趨勢總體上與其在不同狼毒群叢中的優勢度具有一致性,反映出小尺度上物種分布格局與群落分布格局之間具有一定的關聯性(圖6)。

圖6 物種RDA二維排序Fig.6 RDA two-dimensional ordination for species in S.chamaejasme invaded alpine meadow
3 討論
3.1 退化草甸狼毒群落分類特征
近年來在青藏高原高寒草地狼毒大面積出現且快速擴散,形成典型的狼毒型退化草地景觀[7]。本研究采用TWINSPAN分類法結合DCA分析將祁連山中段退化草甸狼毒群落劃分為3種群叢類型(圖2,3),群叢Ⅲ、Ⅱ和Ⅰ的植物種類基本相同,但隨著狼毒入侵加重不同物種的優勢度發生明顯的更替變化:狼毒重要值增加(1.70、1.82、5.64),禾本科植物的重要值減小(4.10、3.12、2.10),豆科植物的重要值先增后減(0.28、0.57、0.25),莎草科植物的重要值增加(0.23、1.18、2.19),各種毒雜草的重要值總體呈明顯上升趨勢(6.10、5.14、10.08),狼毒平均蓋度呈規律地梯度增加。大尺度的研究顯示高寒草甸的退化演替主要表現為草地總蓋度下降、禾本科及莎草科牧草種類減少,毒雜草類種類及數量明顯上升[3,24]。隨著草地退化加劇,禾草類減少而莎草類牧草逐漸增加、莎草類植物具有更強的物種競爭優勢[25]。環青海湖及祁連山地區高寒草甸群落中毒草逐漸發展為優勢種,雜類草的重要性明顯上升,可食性牧草產量大幅度下降[26-27]。本研究與上述成果具有一致性,在小尺度上反映了狼毒型退化草甸逆向演替過程中的群落變化特征。隨著狼毒入侵加重豆科植物的重要值呈波動變化,相對于禾本科和莎草科植物,狼毒分布對豆科植物的影響規律不明顯。同時由于狼毒生長株叢密集,草地群落總蓋度隨著狼毒覆蓋度的增加呈增加趨勢。當覆蓋度達到10%左右時狼毒即替代禾本科的紫花針茅成為群落最主要的優勢種,對原生植被群落構成產生明顯影響。
3.2 狼毒群落分布的環境影響因子
植被分布的環境影響因素隨著空間尺度而變化,降水、溫度等氣候條件是大尺度植被分布格局的主導因素,土壤、地形等環境因子則主導著小尺度的植被群落分布[17]。本研究顯示,在樣地尺度上影響狼毒群叢分布格局的主要環境因子是土壤全磷、全鉀含量,其次是坡向,覆蓋度高的狼毒群叢主要分布于土壤全磷、全鉀含量低,干燥向陽的區域(表3和圖4)。土壤營養元素對群落物種構成有直接影響[18],而坡向控制了局部生境的光、熱、水條件,從而間接反映了土壤養分的空間差異[28],因此局部環境中的土壤養分狀況和微地形條件對于狼毒群叢空間格局的最終形成發揮了重要作用。環境因素對植被群落分布的解釋程度取決于群落自身的復雜性,群落越復雜則環境解釋度越低[29]。本研究中土壤和地形因子對狼毒群落分布變化的解釋率為47.5%,其綜合解釋度與同類相關研究,如宋同清等[30]關于喀斯特峰叢洼地森林群落分布的環境解釋(52.34%),以及余敏等[28]對靈空山小蛇溝林下草本層植物群落梯度的環境影響分析(42.9%)相接近。其中,土壤因子的解釋率明顯大于地形因子,表明土壤因子是影響退化草甸狼毒群落分布的主導因子(圖5)。
斑塊尺度的研究表明狼毒斑塊內土壤有機質含量顯著高于狼毒斑塊間[7,31]。本研究樣方RDA排序結果顯示,狼毒入侵優勢明顯的群落(狼毒蓋度10%~30%)與優勢較弱的群落(狼毒蓋度5%)相比,其生長區域土壤有機質含量相對較高(圖4)。狼毒易于形成肥力島來提高其在高寒草甸的擴散能力,比其他植物明顯更高的生物量是其增加土壤碳庫的有效機制之一[31]。相關研究顯示狼毒斑塊內和斑塊間土壤有機質、全氮含量差異不顯著[32]、狼毒斑塊內全氮含量明顯高于斑塊間[7,31]、斑塊內和斑塊間土壤全磷、全鉀含量差異不顯著[7],本研究在樣地尺度的分析則反映出狼毒群落對低磷、低鉀的土壤環境更敏感,這些研究結果的不一致與研究尺度、采樣深度及群落覆蓋的差異密切相關。狼毒可憑借極發達的根系從深層土壤中汲取氮、磷等營養成分并在表層土壤進行再分配,繼而改變這些養分在群落中的流動及分布格局[31],這可能是不同尺度上研究結果存在差異的主要原因。
3.3 退化草地恢復和狼毒治理措施
狼毒型退化草地治理已經成為祁連山區高寒草地生態保持和生產力恢復亟待解決的重要問題。禁牧封育、劃區輪牧,實施植被自然恢復[33-35],同時結合化學防除、施肥補播等輔助手段是狼毒入侵退化草甸治理的根本措施[36-38],因此,應根據植被群落及其生境對狼毒入侵的響應制定合理的草地治理方案。本研究發現,隨著覆蓋度增加狼毒生長的局部環境內土壤全磷、全鉀含量趨于貧乏,禾本科和豆科植物優勢度有所降低,毒雜草類植物優勢度明顯增高,在植物群落和土壤養分兩個方面呈一定的退化趨勢。當狼毒入侵程度較輕、蓋度達到約10%時草甸群落結構發生明顯變化。在該階段應及時實施劃區輪牧制度,合理配置放牧強度和放牧頻度,使退化草地群落結構向演替恢復方向發展。當入侵加劇、狼毒蓋度達到30%~40%甚至更高時,應采用“狼毒凈”等除草劑,實施化學滅除與短期禁牧封育相結合的綜合措施,同時根據狼毒入侵草地的土壤養分特征采取合理施肥方案,使優良牧草競爭力增強、毒雜草優勢度下降,退化草地生態系統能夠逐步得到恢復。
4 結論
1)祁連山中段退化草甸狼毒群落可劃分為3個群叢類型,Ⅰ:狼毒+紫花針茅+線葉嵩草+披針葉黃華群叢;Ⅱ:狼毒+紫花針茅+委陵菜+美麗風毛菊群叢;Ⅲ:紫花針茅+草玉梅+狼毒+早熟禾群叢。2)狼毒群叢分布格局的主要環境影響因子是土壤全磷、全鉀含量,其次是坡向,土壤養分在狼毒群叢梯度分布中發揮著主導作用,覆蓋度高的狼毒群叢類型對低磷、低鉀的土壤環境更敏感。3)隨著入侵程度加重,狼毒群叢Ⅲ、Ⅱ和Ⅰ的物種優勢度發生明顯更替,狼毒逐漸成為群落中最重要的建群種,禾本科及豆科植物重要值減小,莎草科植物重要值增加,毒雜草重要值及覆蓋度總體呈明顯上升趨勢。
本研究在微觀尺度上探討了狼毒型退化草甸植物群落逆向演替過程中各植物種的生態適應特征及主要環境影響因子,后續將進一步開展區域尺度上狼毒空間格局對氣候、地形等因素的響應關系研究,以期為退化高寒草地修復和狼毒綜合治理提供科學基礎。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
兒童時代·幸福寶寶(2022年12期)2022-12-09 11:24:14
中學生數理化·七年級數學人教版(2022年11期)2022-02-14 07:14:12
科普童話·學霸日記(2020年1期)2020-05-08 16:45:11
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
小天使·一年級語數英綜合(2019年2期)2019-01-10 11:57:30
汽車觀察(2018年10期)2018-11-06 07:05:26
兒童繪本(2018年5期)2018-04-12 16:45:32
