家禽重要性狀的基因組學研究與應用現狀
2022-03-30 03:19:54歐陽清淵胡深強王繼文
畜牧獸醫學報 2022年3期
歐陽清淵,胡深強,王繼文
(四川農業大學動物科技學院,成都 611130)
家禽的育種工作始于20世紀初,接近一個世紀的傳統育種使得家禽的多項生產性能逼近表型選育極限值。基因組學技術的發展成為進一步綜合提升家禽各項生產性能的新途徑。基因組學利用現代生物學技術與分析方法獲得物種基因組序列并鑒定基因的功能[1],家禽基因組學研究的重點在于那些與重要性狀相關的區域,并利用這些區域在遺傳圖譜和物理圖譜中的位置來改良家禽品種。
鑒定與重要性狀相關的區域其關鍵在于要結合表型和基因組上的變異信息。基因組的變異類型主要分為單核苷酸突變(single nucleotide polymorphism, SNP)、插入和缺失(insertion and deletion, Indel)和拷貝數變異(copy number variations, CNVs)3類。檢測全基因組范圍內變異信息的技術手段是多樣的,包括芯片技術、簡化基因組測序(reduced-representation genome sequencing, RRGS)[2]和全基因組重測序等技術。全基因組關聯分析(genome-wide association studies, GWAS)是重要性狀功能基因鑒定常用的分析方法[3],其基于測序和芯片技術在全基因組范圍內尋找與重要經濟性狀相關聯的遺傳變異。選擇消除分析也是挖掘人工馴化過程中和物種適應性進化中受選擇基因的普遍策略[4]。多種測序技術和分析手段在家禽基因組上的綜合運用,加快了我們對于家禽重要性狀的認識和理解,并篩選到一批能夠應用于家禽重要性狀的基因組選擇育種分子標記。這些研究和應用為進一步綜合全面提升家禽各項生產性能提供參考,本文對這些信息進行綜述。
1 家禽參考基因組序列
基因組學研究依賴于基因組的構建。2004年,雞的第一個基因組草圖構建完成[5],標志著家禽的基因組時代正式來臨。家禽參考基因組的大小(1 Gb左右)僅為哺乳動物的一半,但其基因數目僅略少于哺乳動物,其重復元素的收縮、118個大的共素塊的缺失以及相關基因的丟失被認為導致了其特有的緊湊基因組大小[6]。伴隨著高通量測序技術的更新換代,家禽基因組也得到了飛速的發展,雞、鴨、鵝等家禽的基因組已更新升級,紅色原雞的基因組已正式進入6.0版本(表1)。鴨基因組序列與雞基因組序列相比,某些免疫基因家族中檢測到特異性復制,如防御素和親丁酚類基因,這可能有助于其對許多流感毒株的自然抗性[7]。基因組組裝和注釋的完整性直接決定功能基因和位點篩選的準確性。近年來,家禽的基因組組裝也從scaffold逐漸向染色體層面過渡,反映其質量的Contig N50和Scaffold N50得到提高,連續性顯著提升,并且性染色體(ZW)也逐漸被組裝完整。家禽獨特的性染色體特征也得到了廣泛關注,研究報道顯示,雞的性染色體具有擬常染色體區域高度分化和W染色體完全退化的特點[8]。然而,在對包括雞和鴨在內的17種鳥類的性染色體進行比較基因組分析的時候,發現并不是所有鳥類的W染色體都和雞一樣完全退化[9]。由于性染色體的組裝難度和程度,家禽內部性染色體的比較主要發生在雞和鴨之間。與雞相比,鴨的性染色體分化程度較低,其W染色體保留了比雞多2.5倍的基因[10]。家禽性染色體參考基因組序列的成功組裝使得定位一些與性別連鎖的性狀基因的遺傳位點成為可能。

表1 家禽參考基因組基本情況
2 家禽重要性狀的功能基因鑒定和定位
2.1 家禽外貌性狀的功能基因鑒定和定位
外貌性狀作為消費者可以直觀辨別的典型特征受到育種學者的廣泛關注,對這些性狀的遺傳機制開展研究,可能會為家禽提供獨特的品種特征。應用基因組學技術,研究學者對家禽的顏色、羽毛發育和頭部特征等外貌性狀均展開了研究。
2.1.1 家禽色素相關功能基因的鑒定和定位 羽毛、皮膚和脛等不同部位多樣的色素沉著能夠滿足消費者對不同顏色家禽的需求。關于羽毛的色素沉積得到了最廣泛的關注,Park等[21]利用279只雞的60K SNP芯片數據結合其表型進行了GWAS分析,估計了雞毛色的遺傳力,表明雞羽色是一個多基因性狀,并且發現了MC1R、TYR、PMEL、MLPH、ASIP、SOX10和SLC34A2與羽毛黑色素沉著相關。Yang等[22]通過構建黑羽與花羽雞的資源群體定位到與雞羽毛黑色素沉積相關的NUAK1和SHH基因。不同鴨品種之間,羽色也存在較大的差異,Zhou等[13]構建了白羽的北京鴨和綠頭野鴨的參考資源群體,發現鴨基因組中MITF基因內含子的6.6 kb片段插入導致其剪接方式發生變化使鴨產生白羽性狀。Wang等[23]通過對不同羽色類型的建昌鴨進行重測序也證實了MITF基因與鴨白羽性狀相關,并且MC1R基因與麻羽表型相關。Xi等[24]對灰羽和三點花的鋼鵝進行重測序,比較分析發現EDNRB2基因中14 bp的缺失插入與鵝不同的羽色表型相關。鵪鶉通常都表現麻羽性狀,其深淺程度存在差異,Wu等[25]通過種群重測序和GWAS,在Z染色體上發現了包含CCDC171和TYRP1基因一個選擇性掃描區的單倍型與栗色和黃色羽毛密切相關。鴿子的NDP基因也被證實與4種不同的羽色表型相關[26]。黃羽性狀在肉雞消費市場中往往能夠獲得較高的收益,Huang等[27]利用黑色和黃色的雞進行重測序后篩選到可能與黃羽性狀相關的RALY、LGR4、SLC23A2和SLC2A14等候選基因。隨后,有學者基于阿納克紅雞的雜交商業品系和惠陽須雞構建的F2群體,利用全基因關聯技術定位到CABLES1、CHST11、BCL2L1和CHD22與三黃雞的黃羽性狀相關[28]。在五花黃雞與紅色原雞的比較基因組研究結果中也發現PMEL和TYRP1是控制黃羽性狀的候選基因[29]。除了羽色以外,家禽的膚色、脛色和眼瞼等也呈現出不同顏色的變異。家禽的膚色主要包括白膚、黃膚和烏皮3種類型,研究發現控制家禽黃膚和黑皮的功能基因是BCDO2[30]和EDN3[31-32]。脛色主要包括黃色、黑色、白色和豆綠色等,與雞和鴨脛色相關的基因和位點最近均進行了定位[33-35]。SLCO4C1和TP63等基因也被證明參與了家禽紅/白色耳垂的調控[36-37]。
2.1.2 家禽羽毛發育功能基因的鑒定和定位 在羽毛外貌特征方面,部分家禽具有無毛、絲羽和翻毛等性狀,具有這些羽毛特征的家禽可能在一些特定的生產性狀方面異于普通品種[38-39],這使得研究這些獨特羽毛變異的調控機制顯得尤其重要。Wells等[40]通過對有羽雞和無羽雞的血液DNA進行SNP芯片掃描,將控制無羽的基因定位到4號染色體上,并進一步精確到FGF20基因。Ng等[41]通過對一只卷曲雜合子基因型的公雞和5只有正常羽毛母雞的45只后代進行基因組芯片掃描,確定了KRT75保守區有一個69 bp的序列缺失與家禽的卷曲羽毛性狀相關。Dorshorst等[42]使用SNP芯片將與絲羽性狀相關的位點定位在3號染色體上的69.7 Mb區域。Feng等[43]通過對F2群體進行60K芯片掃描以及基因分型結合表型記錄分析,進一步將控制絲羽性狀的位點縮小在3號染色體的18.9 kb區間,并確定了一個位于PDSS2基因上游103 bp的與絲羽性狀完全相關的SNP。毛腿性狀作為某些品種家禽的特定性狀,在對有毛腿和無毛腿性狀的雞進行全基因組重測序后通過比較其Indels變異,篩選到24個可能調控毛腿性狀的基因[44]。
2.1.3 家禽頭部特征功能基因的鑒定和定位 大多數家禽的頭部具有典型特征,不同品種間差異的頭部特征也成為識別家禽品種的重要標識。Shapiro等[17]在構建原鴿基因組的同時,對不同鳳頭性狀的鴿子進行基因組重測序發現,EphB2與鴿的鳳頭性狀密切相關。Zhang等[45]利用鳳頭鴨全基因組重測技術發現,Hoxc8、EphA2、EphA3、EphB2可能是與鴨鳳頭性狀相關的候選基因。針對雞頭部冠型的差異,瑞典烏普薩拉大學研究團隊利用基因組學技術展開了系列研究,結果發現SOX5 基因第一個內含子拷貝數的變異形成豆冠[46],7 號染色體一段 7.4 Mb 序列反轉引起MNR2 同源結構域蛋白基因異位表達形成玫瑰冠[47],EOMES基因上游200 kb調控區20 kb片段串聯重復則形成雙冠[48]。喙畸形也是一種在雞群中比例在3%左右的頭部外貌特征,喙畸形家禽的采食量和飲水量均會降低從而導致生產性能的下降。Bai等[49-50]通過全基因組SNP和CNVs掃描,發現LRIG2可能是調控喙畸形的候選功能基因。
2.2 家禽生長性能的功能基因鑒定和定位
生長性能與生產效益直接相關,對于家禽育種十分重要。然而,由于生長性能受到多基因的調控,并且各學者開展試驗的技術手段和材料存在差異,使得各項篩選得到的位點有差異(表2)。為核實測序結果的準確性,一些研究對篩選位點的功能做了進一步的驗證,Liao等[51]通過GGRS檢測了252只白來杭雞的基因型,發現RBPJ基因與其體重密切相關,并進一步通過表達量檢測探究了其參與表型調控的機制。Zhou等[13]利用1 024只鴨的重測序數據發現一種可能的遠程調控突變導致IGF2BP1基因在出生后持續表達,這種突變可以使鴨的體尺增加15%,飼料效率提高6%。在表觀遺傳方面,Hu等[52]采用甲基化DNA免疫沉淀測序(methylated DNA immunoprecipitation sequencing, MeDIP-seq)方法,研究隱性白洛克和新華雞的全基因組DNA甲基化模式,發現包括與生長發育相關的IGF1R、FGF12、FGF14、FGF18、FGFR2和FGFR3等75個基因的甲基化模式發生了改變。骨發育也是育種學家關注的重點[53-54],興義矮腳雞被認為是骨發育異常的品種,將其與正常骨發育個體雜交構建的家系進行全基因組掃描后分析,發現IHH基因是雞匍匐性狀的功能基因[55]。除了直接與生長發育相關的體重、采食量和骨發育以外,也有學者對屠宰性狀(表3)控制區域進行了定位,利用簡化基因組測序結果證明具有較高經濟價值的鵝肥肝性能與ARAP2、GABRE和IL6等基因有關[56]。

表2 利用基因組學技術篩選與家禽生長發育相關候選基因試驗
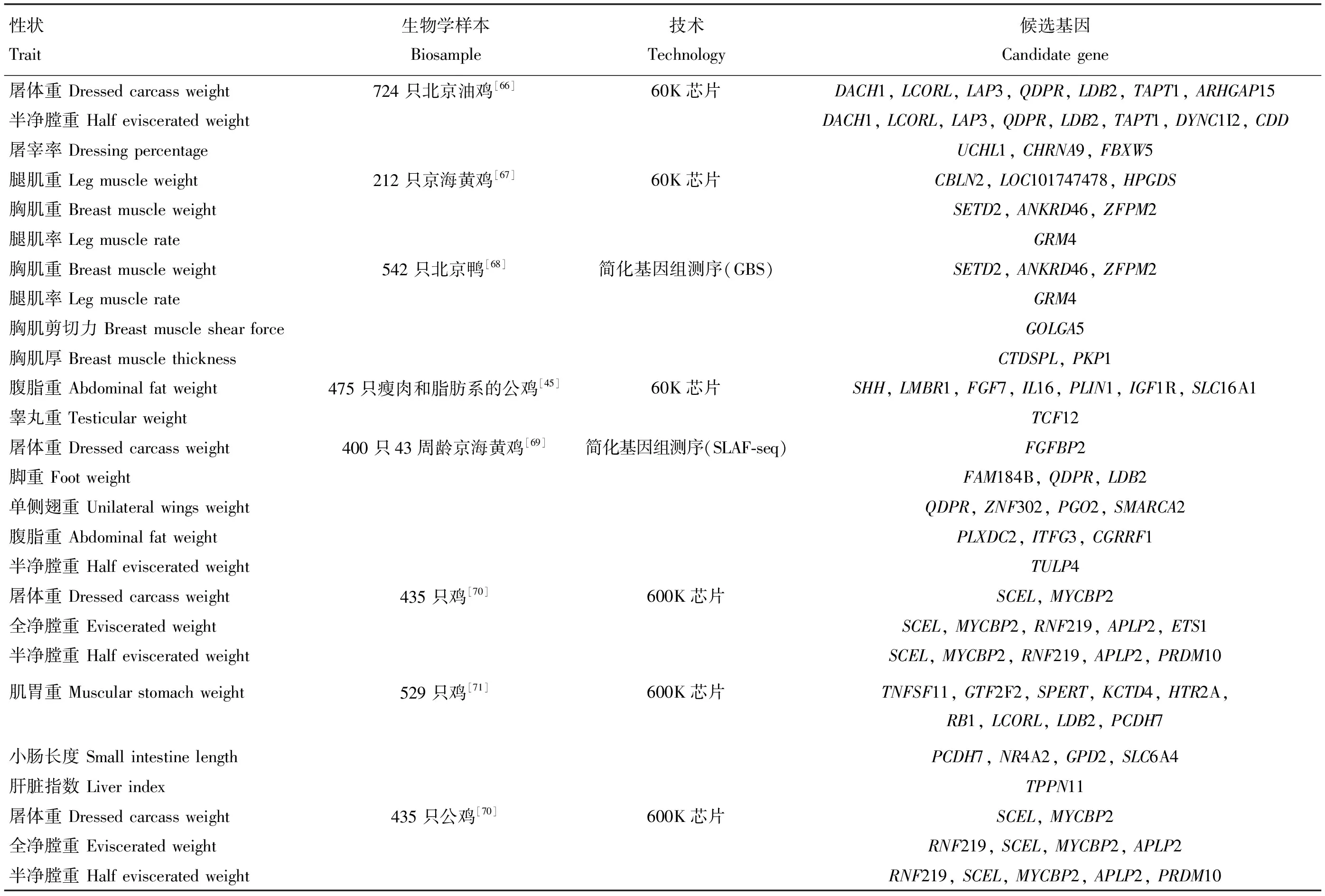
表3 利用基因組學技術篩選與家禽屠宰性狀相關候選基因試驗
2.3 家禽肉質性狀功能基因的鑒定和定位
隨著人們對肉質禽類產品需求的增加,家禽的育種目標也逐漸轉向提高肉質。然而,以往過分選育肉的產量導致禽類肌肉異常發生率也大大增加,如PSE肉、白條紋肉和木質肉等[72]。白條紋肉作為家禽尤其是雞中常見的肌肉疾病,對禽肉外觀以及消費者的購買產生負面影響。白條紋肉具有0.19~0.34左右的遺傳力[73],且含有較高的脂肪和較低的蛋白質[74-75],另外,白條紋肉的持水量和嫩度均轉低,它們還表現出較高的烹飪損失[76]。針對這種禽肉的病理現象,育種學家也采用基因組學的方法進行了研究,發現雞GGA1、GGA17和GGA18號染色體上的部分區域與白條紋肉有關,這些區域的候選基因與肌肉纖維再生和修復相關的肌肉結構和過程(MYH15、MYH1E、MYH1B、MYH1F、MYH13、MYOCD)、脂肪變性和纖維化(PDGFRα)、細胞外基質或肌膜組成(COL6A3、FN1、SGCB)、肌肉代謝(PNPLA7)和人類神經肌肉疾病(FN1、COL6A3、SGCB、LRSAM1)有關[77]。肉色也是影響消費者選擇禽肉的主要視覺因素。多項試驗針對禽類的肉色進行了GWAS分析[78-81],多個與肉色關聯的區域和基因被篩選出來,其中β-胡蘿卜素單加氧酶1(BCMO1)作為候選基因也在不同的試驗中證明其功能與禽類的肉色有關[82-83]。肌間脂肪(IMF)可以通過提高肉的風味、多汁度和柔嫩度等途徑提高肉的質量[84]。Liu等[85]通過59只雞的55K SNP芯片數據和IMF表型數據發現,ACSL1、PPARα、ACADL、FABP6、FABP7、PLPP3、PNLIPRP1、MBOAT1和ALDH3A2這9個與脂質代謝相關的基因可能與肌肉的IMF有關。對雞肌內脂肪和脂肪細胞的甲基化程度比較的試驗研究結果發現,COL6A1基因啟動子的甲基化水平可能與肌內前細胞增殖和分化有關[86]。
2.4 家禽繁殖性狀功能基因的鑒定和定位
針對家禽繁殖性狀的研究主要集中在產蛋量、蛋重和蛋殼顏色上(表4)。除了禽蛋的產量以外,禽蛋質量在近來也逐漸成為研究的熱點。蛋白是消費者判斷禽蛋新鮮程度的重要指標,遺傳背景可以部分解釋個體和品種之間蛋白質量的差異[87]。在眾多的關于禽蛋蛋白質量的基因組學研究中,7號[88-90]和Z[91-92]染色體被多次證明與其相關。禽類的蛋黃由于其較高的營養價值在食品工業中被廣泛的使用,Sun等[93]利用白來杭和東鄉雞的資源群體結合600K的SNP芯片數據證明蛋黃重量的遺傳力為0.25~0.38,并確定了ZAR1、STARD13、ACER1b、ACSBG2和DHRS12可能與蛋黃的重量有關。相比于大量圍繞母禽開展的研究,近年學者逐漸意識到公禽的繁殖性能可能也具有一定的遺傳能力。Wolc等[94]通過收集白來杭雞參考家系F9的1 239只和F6的1 575只公雞的精液質量數據以及F7代的822只公雞的600K SNP芯片數據,證實了公禽繁殖性能的遺傳成分,但未發現有較大影響的QTL。Azmal等[95]也通過190只京紅蛋雞的600K SNP芯片數據結合親本61~69周齡的受精率表型定位到與受精率相關的遺傳位點。公禽精液品質和種蛋的孵化率仍然非常重要,遺憾的是,目前并沒有相關的研究針對這兩個性狀進行基因組的定位。
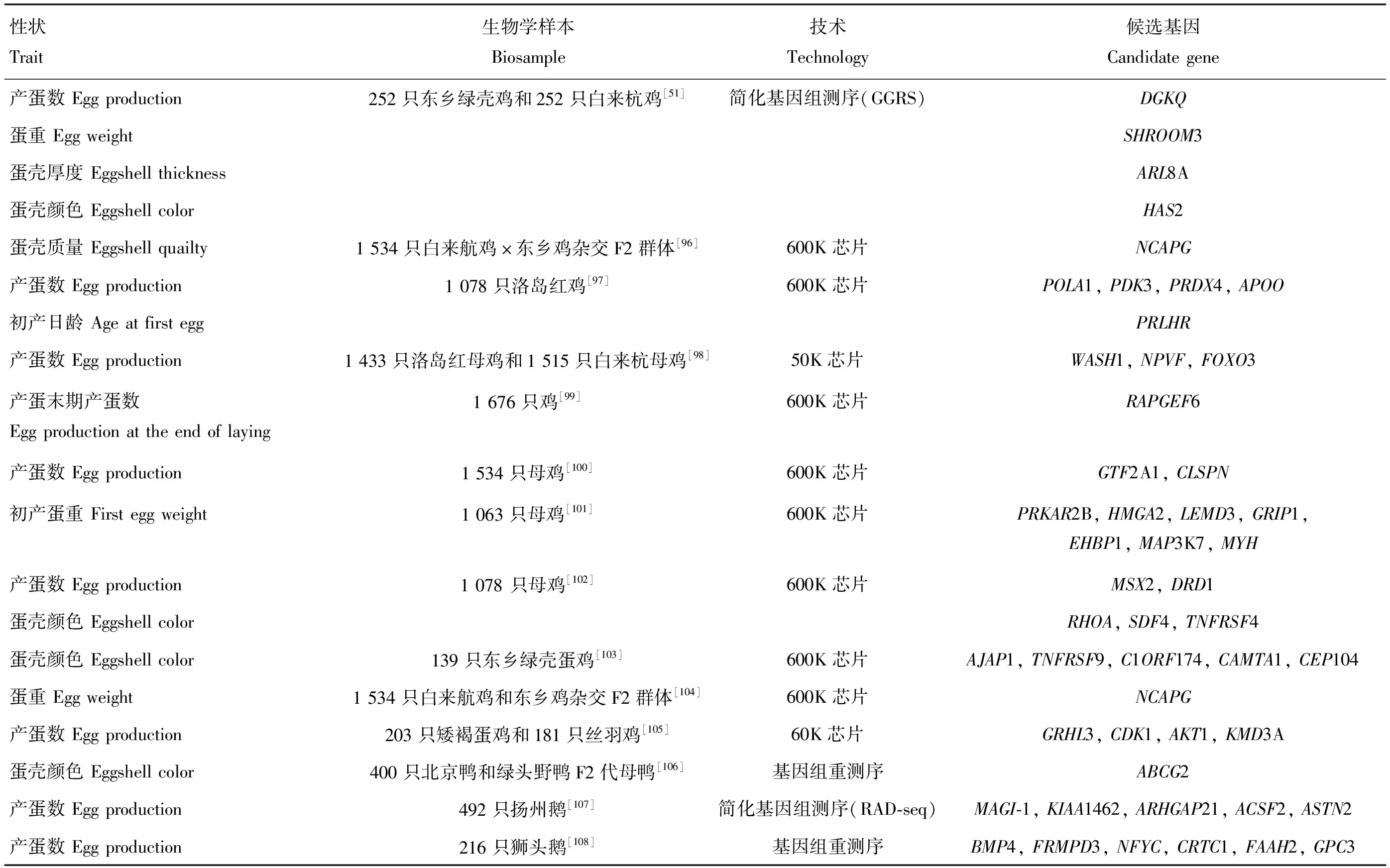
表4 利用基因組學技術篩選與母禽繁殖性狀相關候選基因試驗
2.5 家禽抗病性功能基因的鑒定和定位
近年來,家禽高密度的飼養方式和遺傳多樣性的減少都增強了疾病暴發的可能性。雖然疫苗的出現可以適當緩解一些問題,然而想要更好地解決目前的疾病和阻止可能出現的威脅仍然需要改進控制措施。基因組學領域為解決或改善這些問題提供了途徑。育種學家通過識別數量性狀基因座和控制抗病性狀的基因,利用現有生物多樣性的知識對具有優良抗病性狀的鳥類進行遺傳選擇。馬立克病毒(MDV)是目前應用全基因組學技術研究最多的病毒,Wolc等[109]首次利用貝葉斯變量選擇方法進行GWAS研究,確定與MDV感染蛋雞死亡率相關的基因組區域位于2、3、4、9、15、18和21號染色體。隨后,Li等[110]對57只易感馬立克病毒雞和10只抗性雞進行全基因組SNP掃描,關聯分析和qPCR試驗驗證發現,SMOC1基因可能與馬立克病毒的抗性有關。Yan等[111]對可能涉及MDV抗性的兩個雞品系基因組中的CNVs進行了初步篩選,發現了許多系特異的拷貝數變異,篩選出了一些與該疾病抗性有關的基因,并發現MAPK信號通路可能在宿主對馬立克病毒感染的反應中發揮重要作用。同樣,利用全基因組范圍內的CNVs掃描和分析,Bai等[112]在兩個MDV抗性有差異的自交系中發現IRF2基因與雞的MDV抗性有關。雛雞患病比例較高的沙門菌同樣也得到了關注,在對818只感染沙門菌后表現為死亡和攜帶者的雞進行600 K芯片分型后,通過GWAS分析確定FBXW7和LRBA是參與沙門菌抗藥性的候選基因[113]。新城疫病毒被證明具有中等遺傳力,并通過GWAS分析發現其與ROBO1和ROBO2等基因相關[114-115]。鴨的病毒性肝炎由于傳播速度快和致死性高嚴重危害產業發展,Xu等[116]通過GWAS分析發現,BCHE是一個測定鴨血漿膽堿酯酶(可區分肝病和非肝病)水平的功能基因。
2.6 家禽抗逆性功能基因的鑒定和定位
育種成功的一個關鍵要素是釋放動物的遺傳潛力,以使它們在各種環境條件下都能有最佳的表現。通常而言,雖然商品系家禽能夠達到較高的生產水平,但其對環境的適應能力遠低于地方家禽品種。熱應激是影響家禽生產性能和肉類品質的環境因素之一,它可以使熱休克因子和熱休克蛋白在家禽組織中過度表達從而影響家禽組織的生理機能進而影響生產能力。在過去的幾年中,一些全基因組選擇的研究也涉及熱應激反應的遺傳和分子機制[117-118]。對氣候的適應情況在不同家禽品種間也具有差異,844只臺灣雞和洛島紅雜交F2代60K SNP芯片數據和表型數據通過GWAS定位到與亞熱帶氣候相關的遺傳位點[119]。藏雞是我國唯一的低氧高海拔環境下的地方家禽品種,Wang等[11]對一只雌性藏雞進行de novo測序,并對5只紅色原雞和其他27只不同品種的家雞進行了全基因組重測序,從頭組裝了一個藏雞基因組,揭示了藏雞高海拔低氧適應性進化的遺傳機制。Jiang等[120]對54只藏雞死胚和82只存活藏雞進行60K芯片測序,發現FOXG1可能是藏雞缺氧適應的主要候選基因。Zhang等[121]也通過全基因甲基化研究發現,藏雞和低海拔地區雞的甲基化模式存在差異,這將有助于未來與適應高海拔條件相關的表觀遺傳學研究。
3 家禽重要性狀基因組學研究的應用
在遺傳資源的鑒定和保護方面,通過基因組學技術篩選特定品種的分子標記,可以為家禽遺傳資源保護提供有效保障,支持和引導家禽遺傳資源監測評估。研究者們利用基因組重測序技術檢測分析包括連城白鴨在內的12個鴨遺傳資源的基因組特異性,并開發出一種采用SNP分子標記技術鑒別連城白鴨的方法[122]。壽光雞和太湖雞特異性鑒定的分子標記也同樣得到了開發[123-124]。
對于較多的經濟性狀來說,育種學家通常整合基因組中與性狀相關的突變位點應用到實踐中。一般而言,基因組選擇需要在參考群體中通過已知的表型和候選標記位點的基因型計算標記的效應;隨后,根據估計的效應對已知表型和基因型的候選群體進行基因組育種值的估計;最后,根據基因組估計育種值對個體進行選留實現基因組的選擇育種。2012年,美國安偉捷育種公司宣布將基因組選擇技術應用于蛋雞和肉雞的商業育種流程,2017年開始在火雞中應用,提高了20%到40% 的準確性。在蛋雞繁殖性能方面,安偉捷公司同時對褐殼蛋雞進行為期3年的基因組選擇和表型選擇,發現基因組選擇可以縮小50%的世代間隔,并減少表型測定雞飼養量的3/4[125]。美國科寶公司在2008年就開始研究胸肌重及腿病選育中基因組選擇的應用,并于2015年將基因組選擇全面納入常規育種體系。近年來,中國育種工作者和企業在復雜性狀方面也進行了應用。研究者們利用整合全基因組范圍內與剩余采食量和產蛋數顯著SNP標記效應的一步法全基因組育種值(GEBV)估計方法進行白羽肉雞的選擇,發現與常規一步法GEBV估計結果相比,剩余采食量和產蛋數的選擇準確性分別可提高15.44%和8.1%,可縮短新品系育成的時間,節約選育的成本,提高育種效率,加快我國白羽肉雞自主品系培育進程[126-127]。研究者們結合重測序技術和極端群體表型信息,篩選得到了5個種母雞貯精能力性狀候選位點,從而有效提高了種母雞遺傳改良的效率[128]。而對于更多位點控制的性狀或需要同時對一些性狀進行選育的時候,育種工作者通常在對中外不同家禽品種進行基因組測序的基礎上,整合重要經濟性狀功能基因的顯著位點,研制家禽商業育種芯片進行基因組選擇育種。郝曉東等[129]在黃羽肉雞的育種過程中應用基因組選擇技術,發現其準確性相比BLUP更高,并且可以有效縮短世代間隔和表型測定數量,并且在多性狀選育時能獲得更佳的均衡性。Liu等[130]在724只北京油雞群體中使用60K芯片對雞的IMF進行了全基因組選擇的應用,發現基因組選擇F2代的IMF值比隨機交配F2代的IMF含量增加了9.62%。在白羽肉雞的育種工作中,對2 000只具有表型和全基因組55K芯片基因型的參考群體進行選擇,結果顯示胸肌率提高2.5%,腿肌率提高0.3%。
在品種培育方面,對于由單位點控制的性狀而言,基因組學篩選到的功能基因位點可以在家禽育種工作中得到簡單且有效的應用,目前在色素沉積方面的應用較為廣泛。育種工作者基于前期基因組學分析確定的功能基因和關鍵位點,設計操作簡單、準確率高的用于鑒別特定外貌基因型的PCR引物組合和分型方法,可加快選育具有特定性狀家禽品種的育種進程,縮短育種時間。研究者們利用基因組篩選到的變異位點開發出的分子標記,可快速對特定部位特定顏色個體的基因型進行鑒定,區分開同一表型純合和雜合個體,通過選留基因型純合個體,可保證該群體留種的后代表型一致[131-134]。李光奇等[135]采用全基因組重測序技術對白來航品系突變進行研究,成功鑒定到一個位于 11 號染色體的羽色基因突變點,攜帶該突變的純合型白來航母雞與洛島紅公雞雜交時,后代母雞表現為紅羽,根據該突變位點進行應用,培育出的蛋雞新配套系實現了商品代母雞紅羽比例在98% 以上。
4 展 望
綜上,家禽基因組參考序列組裝程度的不斷完善,為家禽重要性狀基因組區域的挖掘提供了重要保障。隨著高通量芯片和測序技術的發展,各國育種學家運用基因組學技術對家禽不同的性狀展開的研究篩選到了大量的分子標記。然而,這些研究的應用多集中在家禽的外觀、生產和繁殖性狀中,對于肉質、抗病和抗逆性的研究還比較缺乏。在消費多元化和生物安全防控升級的時代背景下,加強基因組選擇在家禽肉質、抗病和抗逆性等方面的研究和應用是后基因組時代家禽育種工作者應該關注的重點。另一方面,對于已有較多組學數據的性狀而言,如果能夠綜合利用各種信息,如比較基因組的信息、受選擇的信息、基因表達信息、調控信息和變異信息,將有助于更加精確地篩選與重要性狀相關的變異位點。因此,一個整合、分析和可視化家禽基因組數據庫的構建可能會加速這些分子標記應用到育種實踐中的進程。
