一株具有拮抗作用黃精內生菌HJ-3的鑒定及抗菌活性研究
2022-03-29 13:50:32呂彩云林永翔劉小壯相愛標鄭亞文柏曉輝
天然產物研究與開發 2022年3期
關鍵詞:植物
呂彩云,夏 娟,林永翔,劉小壯,相愛標,鄭亞文,柏曉輝
黃山學院生命與環境科學學院,黃山 245041
內生菌是一種棲息于健康植物組織中,在長期進化過程中,與植物宿主之間建立和諧共生關系。研究發現植物內生菌在植物病害生物防治方面[1,2]具有巨大應用潛能。也有研究表明,植物內生菌能夠產生與宿主相同或相似的代謝產物,尤其是藥用植物內生菌[3,4]產生的一些代謝產物對人類疾病有很好的療效,已被報道作為治療如癌癥,腫瘤以及細菌和真菌感染治療劑[5,6]。比如Bos等[7]報道用傳統的加拿大藥用植物Heracleummaximum分離的內生菌提取物具有抗分枝桿菌活性,可作為抗癌天然產物研究;Seo等[8]研究發現蘿卜中的內生菌對病原菌具有抗菌活性,可用作對抗人類和植物病原體的生物防治劑等等。
多花黃精(Polygonatumcyrtonema)為百合科黃精屬植物,主要成分為甾體皂苷類和黃精多糖,在增強免疫力、抗腫瘤、降血糖以及抑菌等方面具有非常重要的藥理作用[9]。作為藥食同源植物,現已廣泛應用于藥品、食品、化妝品等領域,但大部分產品處于初加工階段,還需對黃精藥效成分進行深入研究,開發更多的黃精精深加工產品。有關黃精內生菌的文獻報道并不多,如Li[10]從泰山黃精中分離到具有抑菌活性的內生真菌;Wang[11]從浙江多花黃精中分離到內生真菌進行抗菌代謝產物研究。而植物內生菌因其宿主植物地域不同,生長階段不同,組織部位不同,內生菌種類也不同。Bai、Zhai等[12,13]從安徽黃精中分別篩選到內生菌株芽孢桿菌和枯草芽孢桿菌進行了抑菌活性研究。本研究從安徽黃精中分離到另外一株具有拮抗作用的內生菌株HJ-3,對其進行分類鑒定、活性研究及代謝產物分析,以期為豐富黃精內生菌資源及開發利用奠定基礎。
1 材料與方法
1.1 實驗材料
1.1.1 植物樣品
樣品于2015年8月采集自安徽省牯降自然保護區,經黃山學院方建新老師和潘健博士鑒定為多花黃精,采集后于4 ℃保存備用,并在24 h內完成菌種分離。
1.1.2 供試菌株
供試植物病原真菌:盤長孢狀刺盤孢(CollettrichumgloeosporioidesPenz,Cg)、番茄尖鐮孢菌(Fusariumoxysporumf.sp.lycopersici,Fl)、尖孢鐮刀菌(Fusariumoxysporumf.sp.vasinfectum,Fv)、核盤菌(Sclerotiniasclerotiorumde Bary,S)、禾生腐霉(Pythiumgramineacola,P)、新月彎孢(Curvularialunata,Cl)、禾谷鐮刀菌(Fusariumgraminearum,Fg)、立枯絲核菌(RhizoctoniasolaniKühn,R)。
供試細菌:金黃色葡萄球菌(Staphylococcusaureus,S)、大腸桿菌(Escherichiacoli,E)、蘇云金桿菌(Bacillusthuringiensis,Bt)、枯草芽孢桿菌(Bacillussubtilis,Bs);以上菌株均購于中國典型培養物保藏中心并保存于黃山學院生命與環境科學學院微生物學實驗中心。
1.1.3 主要試劑
細菌基因組抽提試劑盒(Takara(北京)生物有限公司);二甲基亞砜(DMSO)、乙酸乙酯、丙酮和75%酒精等均為分析純試劑(上海國藥集團);葡萄糖、蛋白胨、酵母提取物、瓊脂等生化試劑(上海生工生物有限公司);甲醇、甲酸均為色譜純(美國 Fisher公司):純水為MliQ系統純化水(美國Millipore公司)。
1.1.4 主要儀器
DSX280B型高壓蒸汽滅菌鍋(上海申安醫療器械有限公司);MQD-B2R型振蕩培養箱(上海旻泉儀器有限公司);ZGP-2050型恒溫培養箱(上海智城分析儀器制造有限公司);OSB-2100型旋轉蒸發儀(上海愛朗儀器有限公司);FST-III-30超純水機(上海富詩特儀器設備有限公司);AERIS-GB096快速型PCR 儀(上海藝思高科技有限公司);BG-gdsAUTO320凝膠成像系統(北京百晶生物技術有限公司);S-3400N型掃描電鏡(日本日立公司);UPLC H-CLASS+QTOF G2-XS超高效液相色譜串聯四極桿飛行時間質譜儀(美國 Waters公司),配有電噴霧離子源(ESI),MassLynx V4.1軟件。
1.1.5 供試培養基
LB液體培養基:酵母提取物5 g/L,蛋白胨10 g/L,氯化鈉10 g/L,pH 7.0~7.2,用于內生菌的培養。
葡萄糖馬鈴薯固體培養基(PDA):馬鈴薯200 g/L,葡萄糖20 g/L、瓊脂20 g/L,自然pH。用于病原真菌的培養。
1.2 實驗方法
1.2.1 供試菌株活化培養
挑取保存于斜面培養基上的植物病原菌菌絲體接種于PDA培養基,于28 ℃培養,轉接2次,待菌絲長到培養皿2/3時,作為供試病原真菌,4 ℃保存備用。
挑取保存于斜面培養基上的細菌在固體LB培養基平板上劃線活化,接種環挑取單菌落轉接至液體LB培養基,37 ℃ 180 rpm培養12 h,作為供試細菌,4 ℃保存備用。
1.2.2 內生菌分離純化
參照文獻[14,15]方法,將實驗材料經水洗后,用5%(V/V)次氯酸鈉溶液處理5 min;ddH2O漂洗4次后用75%(V/V)乙醇溶液處理5 min,ddH2O漂洗4次并吸去殘留無菌水;再將其無菌切割成3 mm左右片段后分別種植于PDA和LB培養基;同時以隨機選取的根莖片段,將其兩端切口灼燒后種植于上述培養基作為對照。于28 ℃培養3~5天,待切口處長出菌落后,挑取菌體轉接至培養基進一步分離純化,直至純化為單一純種菌落;將純種菌轉接到斜面培養基上保存備用。
1.2.3 內生菌分類鑒定
依據微生物學常規實驗方法對內生菌進行菌落形態特征觀察,并用掃描電鏡進行菌體觀察;同時利用革蘭氏染色鏡檢,結合東秀珠等編寫的《常見細菌系統鑒定手冊》進行初步鑒定。
采用細菌基因組抽提試劑盒提取內生菌株全基因組DNA,利用細菌16S rRNA通用引物27F:5′-AGAGTTTGATCCTGGCTCAG-3′和1 492R:5′-TACGGCTACCTTGTTACGACTT-3′進行擴增[12],PCR反應條件為:95 ℃ 5 min;95 ℃ 30 s,55 ℃ 30 s,72 ℃ 40 s,35 個循環;72 ℃ 10 min。將擴增產物利用1.2%瓊脂糖凝膠電泳檢測,切膠純化后送至上海生工有限公司測序。將測得的序列經過拼接后,利用NCBI數據庫中的BLAST進行比對,檢索與其同源序列。選擇與待鑒定菌株同源性較高的菌株序列,利用Clustal Χ軟件進行比對,再結合MEGA 6.0軟件中Neighbor-Joining算法構建系統進化樹,確定該內生菌株的系統發育學地位。
1.2.4 內生菌對8種植物病原真菌拮抗活性觀察
參考文獻采用五點對峙培養法[16],觀察內生菌對8種植物病原真菌的拮抗作用。在PDA固體培養基平板正中央接入直徑為6 mm的供試病原真菌菌餅,同時在平板4個距供試真菌菌餅2.5 cm處接入相同大小的內生菌菌餅,以只在平板中央接入病原真菌作為空白對照組,于28 ℃恒溫培養,每隔24 h觀察1次,每組處理做三個平行,試驗重復兩次。觀察病原真菌的生長狀態,測量病原真菌菌落直徑,計算病原真菌菌株的抑制率,拍照記錄。
抑制率=(對照組菌落直徑-
處理組菌落直徑)/對照組菌落直徑×100%
1.2.5 內生菌發酵產物提取
將活化的單菌落接種于PD培養液,于28 ℃ 180 rpm培養12 h獲得種子液。按照1/40(V/V)比例將種子液進一步擴大培養,于28 ℃ 180 r/min培養7天,定時觀察培養液情況。
取發酵7天后的培養液,經離心過濾,按照1∶1.5(V/V)比例在發酵液中加入乙酸乙酯萃取,萃取3次;旋轉蒸發儀濃縮,再用丙酮溶解轉至干凈試劑瓶內,自然風干,獲得內生菌發酵產物。用滅菌的100%二甲基亞砜(DMSO)將獲得的發酵產物配成100 mg/mL貯存濃度,于4 ℃保存備用。
1.2.6 觀察內生菌發酵產物對4種供試菌的抑菌活性
用LB液體培養基將供試菌分別培養至OD600為0.2~0.3,取100 μL菌液分別均勻涂布到LB平板上制備菌平板;將平板4等分,在每一部分貼上直徑為6 mm的無菌濾紙片。將貯存的100 mg/mL發酵產物用滅菌的DMSO分別稀釋至60、40、20、10 mg/mL 4個濃度梯度,選擇0.22 μm針頭過濾器除菌后,分別取10 μL滴加至上述無菌濾紙片上,同時以滴加DMSO到濾紙片為對照,于37 ℃倒置培養12 h左右,重復3次,觀察結果。測定抑菌圈的大小,取均值,拍照并記錄。
1.2.7 內生菌發酵產物LC-MS檢測
采用色譜條件:色譜柱Waters BEH C18(2.1 mm ×50 mm,1.7 μm);流速0.3 mL/min;柱溫30 ℃;進樣量1 μL;流動相甲醇(A)和0.1%甲酸水溶液(B);梯度洗脫條件:0~1 min,5% B;1~10 min,5%→90% B;10~15 min,90% B。
質譜條件:電噴霧離子源(ESI),正負離子掃描模式;掃描范圍m/z50~1 500;毛細管電壓3 kV;錐孔電壓40 V;離子源溫度120 ℃;干燥氣溫度550 ℃;干燥氣流量900 L/h。
將按照“1.2.5”所制備的內生菌發酵產物過0.22 μm微孔濾膜,待LC-MS。
2 結果與分析
2.1 內生菌鑒定結果
2.1.1 形態學鑒定
經分離純化所獲得內生菌菌株在LB培養基上生長情況(見圖1)。可見,該菌株在LB培養基上形成的單菌落,均為圓形,菌落較大,表面粗糙隆起,不透明,中間有凹槽,邊緣不光滑,呈輻射狀。經革蘭氏染色呈紫色,疑似革蘭氏陽性菌,在油鏡下可見部分細菌具有明顯芽孢結構。經掃描電鏡觀察(見圖2),可見該菌體呈桿狀、圓端,菌體大小為1.5~1.9 μm ×0.5~0.6 μm。

圖1 菌株HJ-3 純化培養情況Fig.1 Purification and culture of strain HJ-3

圖2 內生菌株HJ-3掃描電鏡圖Fig.2 Scanning electron microscopy of endophytic strain HJ-3
2.1.2 分子鑒定
用16S rRNA通用引物對所獲得的內生菌基因組DNA進行PCR擴增,經1.2%瓊脂糖凝膠電泳結果(見圖3),可見獲得與預期相符的約1 500 bp目的片段,將PCR擴增產物經純化后送至生物公司測序后,所獲得序列進入NCBI 數據庫進行 BLAST同源性比對,利用ClustalΧ和MEGA軟件分析并繪制進化樹(見圖4),結果表明:該內生菌菌株與貝萊斯芽孢桿菌16S rDNA 序列相似度最高,在發育進化樹上與貝萊斯芽孢桿菌聚為一簇。因此將該內生菌菌株鑒定為貝萊斯芽孢桿菌(Bacillusvelezensis),該菌株命名為HJ-3。

圖3 16S rRNA通用引物PCR擴增產物電泳結果圖Fig.3 Electrophoresis results of PCR amplification products with 16S rRNA universal primers注:M:DNA Mark;A:陰性對照;B:菌株HJ-3;C:陽性對照。Note:M:DNA Mark;A:Negative control;B:Strain of HJ-3;C:Positive control.

圖4 菌株HJ-3的16S rRNA系統發育樹Fig.4 Phylogenetic tree of 16S rRNA of strain HJ-3
2.2 內生菌株HJ-3對病原真菌的拮抗活性
采用五點對峙培養法,測定內生菌株HJ-3對8種常見的植物病原真菌的對峙作用(見圖5),結果表明,菌株HJ-3對盤長孢狀刺盤孢、番茄尖鐮孢菌、尖孢鐮刀菌、核盤菌、禾生腐霉、新月彎孢、禾谷鐮刀菌及立枯絲核菌8種病原真菌均表現出較為明顯的抑制作用,通過計算抑制率分析繪圖(見圖6),可見在這8種病原真菌中,菌株HJ-3對立枯絲核菌的對峙作用非常明顯,抑制率高達81.90%;新月彎孢、禾生腐霉、尖孢鐮刀菌、禾谷鐮刀菌、核盤菌和番茄尖鐮孢菌這六種病原真菌抑制率分別為78.74%、73.87%、73.87%、72.16%、71.37%和70.37%,抑制率均超過70%,對峙作用明顯;而盤長孢狀刺盤孢抑菌制率為64.02%,在這8種病原真菌中抑制率最低。
2.3 內生菌發酵產物對供試細菌的抑菌活性
采用濾紙片擴散法檢測內生菌HJ-3不同濃度的發酵產物對4種供試細菌的抑菌活性。從圖7可見,內生菌發酵產物對金黃色葡萄球菌、大腸桿菌、蘇云金芽孢桿菌以及枯草芽孢桿菌4種供試菌均有抑菌作用。從圖8結果可知,不同濃度發酵產物作用于金黃色葡萄球時,發酵產物濃度為40 mg/mL時抑菌效果最明顯,抑菌圈直徑達16 mm左右;當濃度為10 mg/mL,抑菌圈直徑11 mm左右,差異最顯著。不同濃度發酵產物作用于蘇云金芽孢桿菌時,發酵產物濃度為60 mg/mL時抑菌效果最明顯,抑菌圈直徑達13 mm左右。而不同濃度發酵產物作用于大腸桿菌和枯草芽孢桿菌時,抑菌效果基本相似,隨著濃度升高,抑菌作用增強。從圖8明顯可以看出,發酵產物濃度不同,對各種供試菌的抑菌效果不同。四種供試菌中,內生菌HJ-3的發酵產物對金黃色葡萄球菌的抑菌效果要高于其他的三種供試菌。

圖5 內生菌株HJ-3對8種病原真菌的拮抗效果Fig.5 Antagonistic effect of endophytic strain HJ-3 on eight pathogenic fungi
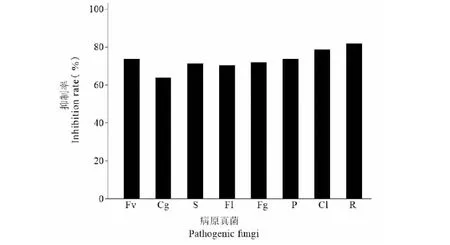
圖6 內生菌株HJ-3對8種病原真菌的抑制率Fig.6 Inhibition rate of endophytic strain HJ-3 against eight pathogenic fungi
2.4 內生菌發酵產物LC-MS檢測分析
取發酵液進LC-MS分析,正負模式總離子流見圖9,同培養液對比,發酵液中檢出了多種組分,其中1號信號峰m/z為339.232 6[M-H]-,經儀器軟件自帶插件Element composition計算其元素組成為C23H32O2,對該物質做二級質譜,采集信號如圖10所示,該化合物產生特征碎片m/z163.112 1為[M-H-C12H17O]-,從Chemspider及文獻[17]中查閱到該組分的分子式結構。由Chemdraw畫出該物質結構式經質譜儀器軟件自帶插件Massfragment識別出該物質碎裂過程,該物質可能從中間斷裂,脫落后形成m/z163.112 1碎片。與吳盼關于樺褐孔菌發酵液研究的文獻[14]報道的化合物相吻合,確定化合物為5,5′-methylenebis(2-tert-butyl-4-methylphenol),是一種酚類物質,裂解示意圖見圖10。經文獻[18]可知,樺褐孔菌發酵液在抗菌、抗腫瘤、抗氧化等方面具有生物活性功效。同時經正負模式及其加氫/脫氫和加鈉比較分析,發現2號峰m/z1 006.633 7[M-H]-、3號峰m/z1 020.652 3[M-H]-、4號峰m/z1 034.674 1[M-H]-分別相差-CH2,應為同系物,經儀器原件自帶插件推算分子式分別為C55H93NO15、C56H95NO15、C57H97NO15。分析這些化合物,發現都具有特征性的離子碎片m/z678.475 3[M-H]-、m/z452.285 2[M-H]-、m/z339.202 3[M-H]-,進一步說明2、3、4號化合物含有共同的基團,可能是同系列化合物,至于何種化合物有待下一步解析。

圖7 內生菌株HJ-3發酵產物對不同供試菌的抑菌效果Fig.7 Antibacterial effects of fermentation products of endophytic strain HJ-3 on different tested bacteria

圖8 內生菌株HJ-3發酵產物對不同供試菌的抑菌圈直徑Fig.8 Diameters of inhibitory zone of fermentation products of endophytic strain HJ-3 against different tested bacteria注:*P<0.05,**P<0.01,***P<0.001。

圖9 正負離子模式下發酵液總離子流圖Fig.9 Total ion flow diagram of zymotic fluid in positive and negative ion mode
3 討論與結論
內生菌在與宿主共生的過程中會產生許多具有藥用價值的生物活性物質,在抗癌、抗菌、抗氧化等人類疾病防治以及農業病蟲害防治領域具有潛在的價值。國內外針對內生菌相關的研究報道很多,比如Halim等[19]從水稻田的根葉中分離出BCA 3和BCA 12兩種內生菌,研究發現其對細菌性葉枯病(BLB)具有積極的拮抗活性;灰葡萄孢是最重要的收獲后真菌病原體之一,Chao等[20]從不同植物中篩選出內生細菌,發現對番茄中的灰葡萄孢具有生防作用;本課題獲得的內生菌HJ-3研究發現對植物病原真菌具有拮抗作用,而且表現出廣譜的拮抗活性。Zhang等[21]從小麥小穗中分離出一株具有廣譜拮抗細菌XS-2,發現對小麥赤霉病(FHB)的病原體禾谷鐮刀菌、棉花病原體立枯絲核菌等6種植物病原體具有抑制作用,抑菌圈在10~29.58 mm之間,使其成為寶貴的生物資源,用于開發生物防治劑。本課題所獲得的內生菌HJ-3對立枯絲核菌、禾谷鐮刀菌等植物病原真菌具有同樣抑制效果,所不同的是該內生菌不是從農作物分離,而是從藥用植物黃精分離。科學家研究表明,從植物尤其是一些藥用植物分離內生菌,可以抑制病原細菌、病原真菌生長,原因可能是內生菌與藥用植物共生過程中會產生代謝產物,這些代謝產物對病蟲害或病原菌產生抑制作用[22]。Bai等[12,13]從藥用植物黃精中分離出內生菌株HJ-1和HJ-2,發現其發酵產物對綠膿桿菌、金黃色葡萄球菌等病原細菌具有很好的抑制作用,本研究從藥用植物黃精分離內生菌HJ-3,也表現出對常見病原菌廣泛的抑菌作用,但本研究重點探討了內生菌對植物病原真菌的拮抗作用,進而豐富了黃精內生菌潛在應用價值。已有資料表明,從藥用植物篩選內生菌用于生產具有藥用價值的生物活性化合物,特別是從瀕臨滅絕的宿主植物內生菌中生產有用的生物活性化合物,成為新藥開發生物活性化合物的重要來源之一。

圖10 化合物C23H32O2化學結構圖和MS圖譜Fig.10 Chemical structure and MS spectra of compound C23H32O2
該課題是從安徽道地藥材黃精內篩選到一株內生菌株HJ-3,經形態初步鑒定以及16S rRNA分子生物學方法測序,將該菌株鑒定為貝萊斯芽孢桿菌(Bacillusvelezensis)。貝萊斯芽孢桿菌因其具有廣譜的抗菌活性、促進植物生長的作用以及具有作為生物表面活性劑的潛力被廣泛應用到工農業和環境保護領域,還可以作為飼用微生物菌劑,同時有望在醫療保健方面發揮更重要的作用。但同種不同株B.velezensis在功能上存在著差異,因此還需要進一步發掘自然界中B.velezensis的新菌株資源,以開發利用新功能菌株。Chi等[23]也從多花黃精內分離到貝萊斯芽胞桿菌株ZJU-3,研究了內生菌及其代謝產物對尖孢鐮刀菌拮抗活性。本研究所分離的B.velezensis菌株HJ-3對植物病原真菌除了尖孢鐮刀菌外,還有盤長孢狀刺盤孢、番茄尖鐮孢菌、核盤菌、禾生腐霉、新月彎孢、禾谷鐮刀菌及立枯絲核菌均有抑制作用,尤其是對立枯絲核菌具有非常明顯抑制作用,其次是新月彎孢、禾生腐霉、尖孢鐮刀菌、禾谷鐮刀菌、核盤菌和番茄尖鐮孢菌。由此可見,本研究所獲得的黃精內生菌HJ-3對常見植物病原真菌具有廣譜的拮抗活性,其研究結果對農業病蟲害生物防治具有一定指導意義。此外研究發現,內生菌株HJ-3的發酵產物對常見細菌表現出廣泛抑菌效果,尤其是對金黃色葡萄球菌,抑菌作用最為明顯,其中金黃色葡萄球菌是臨床常見的病原菌,其抗生素耐藥現象非常常見。因此,尋找有效的抗菌藥物,尤其是從天然產物中尋找新型的抗菌藥物對于細菌性疫病的防治具有非常重要的意義,也是解決細菌耐藥性問題的方法之一。
經以上研究表明,從中藥黃精中篩選到的HJ-3內生菌株對多種植物病原真菌具有明顯的拮抗效果,其發酵產物能抑制大腸桿菌等常見細菌的生長。通過UPLC-MS/MS,從內生菌發酵產物中檢測到5,5′-methylenebis(2-tert-butyl-4-methylphenol)酚類化合物,同時獲得三個同系列化合物。本研究獲得數據將為進一步挖掘黃精內生菌發酵產物中的活性成分奠定基礎,也為植物內生菌藥物廣泛應用提供依據。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
