陜北安塞不同林齡刺槐林下幼苗更新的動態變化特征
2022-03-15 05:20:20宗小天焦菊英吳多洋
水土保持研究 2022年2期
關鍵詞:物種
宗小天, 焦菊英,2, 吳多洋, 寇 萌, 徐 倩
(1.西北農林科技大學 水土保持研究所 黃土高原土壤侵蝕與旱地農業國家重點實驗室, 陜西 楊凌 712100;2.中國科學院 水利部 水土保持研究所 黃土高原土壤侵蝕與旱地農業國家重點實驗室, 陜西 楊凌 712100;3.陜西師范大學 西安 710000; 4.咸陽師范學院 資源環境與歷史文化學院, 陜西 咸陽 712000)
幼苗是植被恢復演替過程的基礎[1],也是植物生活史中對環境條件反映最敏感的時期[2]。幼苗通常會參與到植物群落更新的過程中,影響未來植被的種類組成、結構、動態變化和物種多樣性維持[3-5],從而影響著植被恢復。由此可見,幼苗更新是植被生態恢復和植物群落演替過程極其關鍵的環節[6],對植物種群的增殖、擴散、延續及穩定性維持具有極其重要的作用。因此,加強對幼苗的研究在一定程度上有助于了解植被恢復演替趨勢,為生態脆弱區的植被恢復重建提供理論依據。迄今為止,許多學者對黃土高原地區幼苗已有大量研究并取得了重要進展,研究內容主要涉及幼苗的更新特性[7-8]、幼苗生長影響因素[9-10]、幼苗與周圍植被的關系[1,5,11-12]、幼苗的種群結構與空間分布格局[13-14]。但前人的研究多是對幼苗進行定性描述或靜態定量比較,缺乏對幼苗更新的動態變化研究[15]。
陜北黃土丘陵溝壑區氣候變化劇烈、地形地貌復雜,以及人類在該區不合理的土地利用方式,使其已成為水土流失最嚴重的地區之一,同時也是生態環境極其脆弱的地區之一[16-17]。因此,中國于20世紀末在黃土高原大規模實施“退耕還林(草)”工程,用于防治黃土高原地區水土流失,改善脆弱的生態環境[18-19]。植被恢復是生態環境建設的核心和退耕還林還草的關鍵[20-21],可以有效地遏制土壤退化、促進退化生態系統恢復[22-23]。研究表明人工林可通過次生演替恢復退化生態系統[24]。其中,刺槐林是黃土高原地區植被恢復的主要人工林之一,因其較強的適應能力和抗逆能力得以在黃土高原廣泛種植[25]。但部分地區人工刺槐林卻出現了林分退化、生長減緩、生態效益低、林下植被稀疏、樹種單一等現象,導致黃土高原森林生態系統服務功能有下降的趨勢[26-27]。林下植被是人工林生態系統的重要組成部分,它能夠通過促進上層林木更新、改良土壤、增強林地水文效應、促進養分循環來維護人工林生態功能穩定性[28]。林下幼苗是林下植被生長發育的基礎,它的更新變化對人工林恢復和演替有著極其重要的作用。因此研究刺槐人工林恢復進程中林下幼苗更新的動態變化對營造一個健康刺槐人工林具有重要指導作用。目前關于刺槐林的研究大多集中在生理機能、土壤水分養分等方面[10],對于人工刺槐林下幼苗的研究較少。鑒于此,本研究對陜北安塞紙坊溝流域不同林齡刺槐林下的幼苗進行跟蹤觀測,分析幼苗更新的動態變化特征,以期為黃土高原地區人工林生態系統恢復與生態功能的提升提供一定的理論依據。
1 材料與方法
1.1 研究區概況
研究區位于陜西省延安市安塞區(36°30′45″—37°19′3″N,108°5′44″—109°26′18″E)。該區屬典型黃土丘陵溝壑區,地形地貌復雜多樣,境內溝壑縱橫,平均海拔約1 372 m。氣候類型屬暖溫帶半干旱大陸性氣候,年均溫8.8℃,年均降雨量505 mm,降雨年內分布不均,其中6—9月雨季降水量占年降水量的72.9%,年均無霜期160 d[29]。該區原始植被破壞嚴重,天然林較少,人工林以刺槐(Robiniapseudoacacia)、小葉楊(Populussimonii)、檸條錦雞兒(Caraganakorshinskii)、沙棘(Hippophaerhamnoides)為主,草本植物群落以細裂葉蓮蒿(Artemisiagmelinii)、白羊草(Bothriochloaischaemum)、長芒草(Stipabungeana)為主[30]。土壤類型以黃綿土為主,土質疏松,抗蝕抗沖能力較差[31]。
1.2 樣地選擇與調查
于2016年4月,在陜北安塞紙坊溝流域內選取4個不同恢復年限的刺槐林(10 a,20 a,30 a,40 a),每個林齡設置2個10 m×10 m的樣地,共計8個樣地。在每個樣地內設置4個1 m×1 m的固定小樣方,對樣方內的所有幼苗進行調查,并用竹簽對幼苗進行標記。于2016年7月份調查樣地的植被情況(表1),包括植被蓋度、郁閉度、地上植被物種組成等,同時記錄樣地的坡度、坡位等信息。
幼苗調查:即對一定面積的樣地中調查樣方內小于某一高度的所有植物幼苗進行調查。根據物種的生活型、高度等,將喬木物種高度小于50 cm的植物、灌木物種高度小于40 cm、草本物種高度小于10 cm的植物作為幼苗;同時考慮物種特性,如地錦屬于低矮草本植物,則將其高度小于5 cm的作為幼苗。于2016年4—10月,對小樣方內的幼苗逐月跟蹤調查,記錄小樣方內幼苗的物種組成、數量、生長狀況及死亡情況等。在跟蹤調查中,每月用不同顏色的竹簽對幼苗進行標記,以區分不同調查時間幼苗的存活與萌發數量。
地上植被調查:在每個樣地內設置2個2 m×2 m的小樣方,進行地上植被調查,記錄植被的蓋度、物種組成。
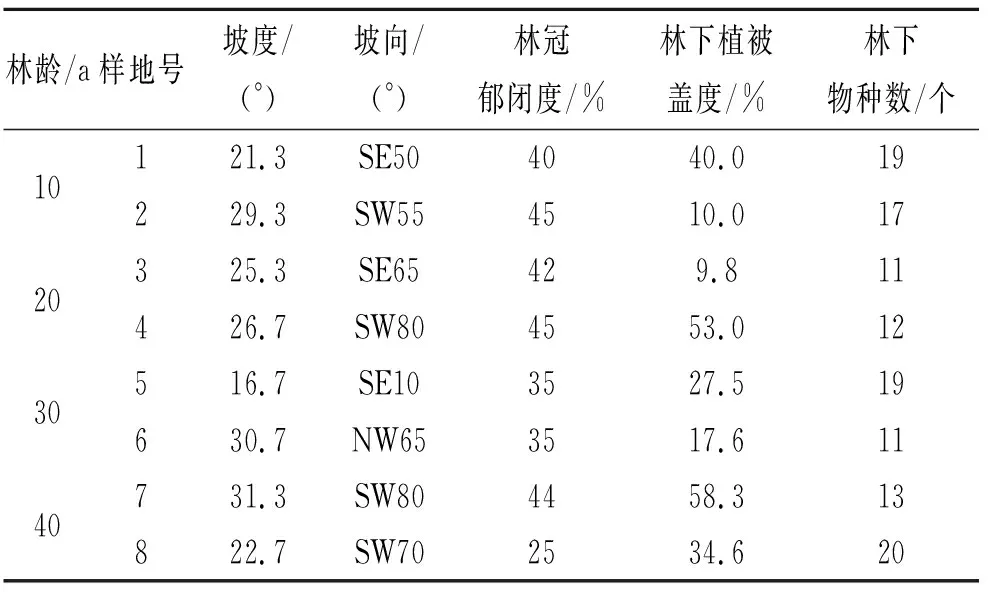
表1 樣地基本情況
1.3 數據處理
(1) 采用相對多度和相對頻度來計算不同幼苗物種的重要值,將重要值大于5%的物種作為主要物種[26],計算方法如下:
幼苗重要值=(相對多度+相對頻度)/2
(1)
式中:相對多度=(某物種的多度/所有物種的多度)×100%,相對頻度=(某物種的頻度/所有物種的頻度)×100%
(2) 采用Sorensen相似性系數逐月計算不同林齡刺槐林樣地內幼苗與地上物種間相似性,計算公式如下:
Sorensen相似性系數(CC)=2C/(A+B)
(2)
式中:C為共有物種數;A為幼苗物種數;B為地上植被物種數。
采用Excel 2019軟件整理調查數據,利用SPSS 22.0軟件進行差異顯著性分析(不同林齡幼苗的物種數、密度、幼苗與地上植被間相似性系數),顯著水平為p<0.05。
2 結果與分析
2.1 幼苗與地上植物的物種組成及相似性
在4—10月調查期間,共記錄幼苗48種(10 a林25種,20 a林28種,30 a林24種,40 a林23種)(表2),隸屬于20科39屬,其中菊科最多(17種),禾本科(5種)、豆科(4種)次之,三科幼苗物種數占總物種數的54.2%。多年生草本植物最多(25種),占總物種數的52.1%;一年生草本植物次之(12種),占24.9%;灌木(6種)占12.4%;喬木(3種)占6.3%;藤本(1種)、二年生草本植物(1種)各占2.1%。
地上植物共記錄51種(10 a林20種,20 a林19種,30 a林21種,40 a林26種)(表2),隸屬于16科40屬,菊科、豆科、禾本科物種數占總物種數的50%以上。多年生草本植物(27種)占總物種數的52.9%;一年生草本植物(10種)占19.6%;灌木(8種)占15.7%;喬木(4種)占7.8%;藤本(1種)、二年生草本植物(1種)各占2.0%。地上植物和幼苗的物種數類似,兩者都是以菊科、禾本科、豆科為主;就生活型而言,二者物種組成都是以多年生草本為主、一年生草本次之。
不同林齡林下自然更新和生存的物種種類差異明顯。10 a林下幼苗與地上植物都存在的物種有13種,其中出現月份較多的物種有阿爾泰狗娃花、苦苣菜、豬毛菜、豬毛蒿、田旋花;僅在幼苗中存在且出現月份較多的物種有刺槐、狗尾草、敗醬。20 a林下幼苗與地上植物都存在的物種有10種,其中出現月份較多的物種有臭草、牻牛兒苗、裂葉堇菜;僅在幼苗中存在且出現月份較多的物種有阿爾泰狗娃花、刺槐、狗尾草、翠雀。30 a林下幼苗與地上植物都存在的物種有7種,其中出現月份較多的物種有長芒草、裂葉堇菜;僅在幼苗中存在且出現月份較多的物種有刺槐、翠雀。40 a林下幼苗與地上植物都存在的物種有9種,其中出現月份較多的物種有臭草、長芒草、裂葉堇菜、刺槐;僅在幼苗中存在且出現月份較多的物種有翠雀。
不同月份,林下幼苗與地上植物間相似性系數變化明顯(表3),不同林齡相似性系數變化范圍分別為0.17~0.56,0.23~0.50,0.08~0.40,0.20~0.38,說明幼苗與地上植物間相似性較小。總體表現為10 a林下幼苗與地上植物間相似性系數最大(0.40),其次是20 a,40 a,30 a,分別為0.32,0.29,0.24,30 a顯著小于其他林齡(p<0.01)。從月變化趨勢來看,10 a,20 a,40 a林下幼苗與地上植被間相似性系數表現為下降(4—5月)—上升(5—6月)—下降(6—8月)—上升(8—9月)—下降(9—10月),30 a表現為下降(4—8月)—上升(8—10月)。

表2 幼苗與地上植物組成

續表2

表3 幼苗與地上植被間相似性系數
2.2 幼苗物種組成的月動態變化
由表4看出,在10 a刺槐林下,4月幼苗主要有豬毛菜、苦苣菜、刺兒菜、鬼針草、興安胡枝子等;5月調查時,豬毛蒿、刺兒菜、興安胡枝子幼苗大量死亡,狗尾草大量萌發,幼苗主要有鬼針草、狗尾草、豬毛菜、苦苣菜、風毛菊等;6月狗尾草、豬毛菜、苦苣菜幼苗大量死亡,刺槐、杠柳幼苗萌發較多,幼苗主要有鬼針草、刺槐、杠柳、牻牛兒苗等;7月遠志、狗尾草、香青蘭開始萌發,幼苗主要有刺槐、遠志、狗尾草、香青蘭、阿爾泰狗娃花等;到8月時,香青蘭大量死亡,敗醬、抱莖小苦荬、豬毛蒿開始萌發,幼苗主要有刺槐、敗醬、抱莖小苦荬、豬毛蒿、狗尾草等;9月幼苗更新達到高峰,主要物種有刺兒菜、刺槐、抱莖小苦荬、敗醬等;10月幼苗大量死亡,僅有敗醬、刺槐、長芒草和田旋花4種幼苗存活。
20 a刺槐林下,4月幼苗主要有豬毛菜、裂葉堇菜、細裂葉蓮蒿、牻牛兒苗等;5月豬毛菜大量死亡,草木樨狀黃耆、狗尾草等幼苗開始萌發,幼苗主要有裂葉堇菜、細裂葉蓮蒿,牻牛兒苗、草木樨狀黃耆、狗尾草等;6月刺槐幼苗大量萌發,同時臭草、翠雀、杠柳開始萌發,主要幼苗有刺槐、裂葉堇菜、臭草、翠雀等;7月抱莖小苦荬開始萌發,主要幼苗有刺槐、翠雀、裂葉堇菜、抱莖小苦荬、臭草等;8月和7月類似,但臭草死亡較多,有新物種黃花蒿開始萌發;9月刺槐幼苗死亡較多,狗尾草、臭草、阿爾泰狗娃花開始萌發,主要幼苗有抱莖小苦荬、裂葉堇菜、狗尾草、刺槐、臭草、阿爾泰狗娃花等;到了10月份,有大量幼苗死亡,幼苗主要是臭草、刺槐、裂葉堇菜、狗尾草、阿爾泰狗娃花、長芒草、黃花蒿。
30 a刺槐林下,4月主要幼苗有裂葉堇菜、長芒草、翠雀、臭蒿、阿爾泰狗娃花、細裂葉蓮蒿等;5月裂葉堇菜大量死亡,刺槐、苦苣菜、興安胡枝子、狗尾草幼苗出現,主要幼苗有刺槐、長芒草、翠雀、苦苣菜、興安胡枝子等;6月裂葉堇菜重新萌發,又有杠柳、龍葵開始萌發,幼苗主要有裂葉堇菜、刺槐、杠柳、龍葵、翠雀等;7月大量幼苗死亡,僅有裂葉堇菜、長芒草存活;8月有刺槐、鶴虱、裂葉堇菜、刺兒菜幼苗萌發;9月主要幼苗有裂葉堇菜、刺槐、臭草等;10月長芒草大量萌發,主要幼苗有長芒草、裂葉堇菜、鶴虱、刺槐等。
40 a刺槐林下,4月幼苗主要有野菊、艾、臭草、裂葉堇菜、蒙古蒿、長芒草等;5月刺槐、翠雀幼苗開始萌發,主要有臭草、長芒草、艾、裂葉堇菜、刺槐、翠雀等;6月刺槐、翠雀大量萌發,榆樹、臭椿幼苗開始萌發,幼苗主要有刺槐、翠雀、榆樹、臭椿、裂葉堇菜等;7月翠雀、榆樹、臭椿幼苗大量死亡,幼苗主要有茅莓、刺槐、延安小檗、裂葉堇菜、細裂葉蓮蒿、中華小苦荬等;8月翠雀、榆樹幼苗重新萌發,幼苗主要有翠雀、刺槐、榆樹、延安小檗、臭草、鶴虱等;9月主要幼苗是刺槐、臭草、長芒草、茜草、野菊等;10月幼苗大量死亡,僅有臭草、阿爾泰狗娃花、香青蘭、刺槐等幼苗存活。
從4—10月出現的幼苗種類來看,10 a林以刺槐、狗尾草、豬毛菜為主,幼苗數量分別占幼苗總數的26.3%,17.5%,10.9%;20 a林以豬毛菜、刺槐、臭草、狗尾草、裂葉堇菜為主,幼苗數量分別占幼苗總數的14.8%,15.4%,13.1%,11.4%,10.6%;30 a林以裂葉堇菜、長芒草、刺槐為主,幼苗數量分別占幼苗總數的30.5%,16.9%,14.8%;40 a林以刺槐、翠雀、苦苣菜為主,幼苗數量分別占幼苗總數的26.4%,10.6%,10.2%。可以看出,不同林齡林下幼苗中均有大量刺槐幼苗萌發,隨林齡的增加,其他主要幼苗發生明顯變化,依次為狗尾草、豬毛菜→豬毛菜、臭草、狗尾草、裂葉堇菜→長芒草、裂葉堇菜→翠雀、榆樹(表5)。
2.3 幼苗物種數和幼苗密度的月動態變化
2.3.1 幼苗物種數月動態變化 刺槐林下幼苗物種數的月動態變化存在差異(表6)。在10 a刺槐林中,6月幼苗物種數最多,10月最低;4月、6月、9月幼苗物種數顯著高于5月、10月(p<0.05)。20 a刺槐林中,4月、6月、9月幼苗物種數較高,其中6月幼苗物種數顯著高于10月(p<0.05)。30 a刺槐林中,4月、6月、10月幼苗物種數相對較高且顯著高于7月、8月(p<0.05)。40 a刺槐林中,4月、5月、6月、9月存活的幼苗種類顯著高于7月、10月(p<0.05)。可以看出,在不同林齡刺槐林下,4月、6月、9月幼苗物種數均較高,但30 a刺槐林下幼苗種類在10月也較多。隨林齡增加,幼苗物種數呈降低趨勢,其中10 a林幼苗物種數顯著大于40 a林(p<0.05)。
2.3.2 幼苗密度的月動態變化 刺槐林下幼苗密度的月動態變化存在差異(表7)。10 a刺槐林中,4月幼苗密度最高,10月幼苗密度最低;4月、6月、7月幼苗密度較高且顯著高于5月、9月、10月(p<0.05)。20 a刺槐林中,9月幼苗密度最高,4—8月幼苗密度呈降低趨勢;10月幼苗密度顯著低于9月(p<0.05)。30 a刺槐林中,4月、6月、10月幼苗密度相對較高;7月、8月幼苗密度顯著低于4月份(p<0.05)。40 a刺槐林中,6月幼苗密度顯著高于其他月份(p<0.05)。可見,在不同林齡刺槐林下,4月、6月幼苗密度相對較高;除30 a刺槐林外,其他林齡林下10月幼苗密度均最低。隨林齡增加,幼苗密度呈降低趨勢,其中10 a,20 a林幼苗平均密度顯著大于30 a,40 a林(p<0.05)。
2.4 幼苗更新能力和存活能力
由表2可知,不同林齡5—10月均有刺槐幼苗出現,存活時間較長,是刺槐林下優勢物種。裂葉堇菜幼苗在20 a,30 a刺槐林下4—10月均有出現,在40 a刺槐林下4月、5月、7月、8月出現,說明在刺槐林恢復中期裂葉堇菜有著很強的更新和存活能力。田旋花幼苗在10 a刺槐林下出現頻率較高,但在20 a,30 a,40 a刺槐林下極少出現,說明在刺槐林恢復初期其更新能力較強。臭草幼苗在20 a,30 a,40 a刺槐林下出現頻率較高,說明在刺槐林恢復中、后期臭草有著很強的更新和存活能力。長芒草幼苗在30 a,40 a刺槐林下存活時間長,同時隨著林齡的增加,長芒草在刺槐林下4—10月出現頻率增加,說明隨林齡增加,長芒草的自然更新和存活能力增強。豬毛菜幼苗在10 a,20 a刺槐林4月大量萌發但隨后很快死亡;風毛菊、苦苣菜、豬毛蒿幼苗幾乎只存活在10 a刺槐林下,在4月、5月,7—9月偶爾出現;抱莖小苦荬主要在10 a,20 a刺槐林下7—9月開始萌發,且在出苗后的2個月左右全部死亡;鬼針草主要存活在10 a刺槐林中,4月少量萌發,5月、6月大量萌發,但在7月全部死亡;阿爾泰狗娃花、刺兒菜、遠志、敗醬、翠雀、杠柳、艾、麻花頭、糙隱子草、蒙古蒿等在出苗1—2月內死亡;這些幼苗存活時間較短,難以成功建植。

表4 刺槐林下幼苗物種組成月動態變化
3 討論與結論
(1) 幼苗和地上植物都以菊科、禾本科、豆科為主;物種組成都是以多年生草本為主,一年生草本次之;這與白文娟等在黃土丘陵溝壑區的研究結果一致[32]。幼苗與地上植被間相似性系數基本在0.5以下,說明幼苗與地上植被間相似性較小。造成這種現象的因素是多方面的。第一,部分地上植被的種子未能進入種子庫中[33];第二,黃土丘陵溝壑區土壤侵蝕嚴重,土壤水分、養分均較低,地上植被的種子進入土壤種子庫后在萌發成幼苗前就已經死亡[32];此外物種生活特性[34]、優勢物種與林下植物的的競爭作用等[5]也會對幼苗與地上植物的相似性產生一定影響。
(2) 不同林齡林下幼苗中均有刺槐幼苗萌發,隨林齡增加,其他主要幼苗種類差異明顯。這種差異性可能與地上植被組成[44]以及物種生物學特性有關。Maranon等[35]研究表明地上植被與土壤種子庫有著密切聯系,而幼苗更新的基礎是土壤種子庫[4]。如10 a刺槐林下,豬毛菜、苦苣菜、鬼針草等地上植物提供的種子在4—5月大量萌發;20 a,30 a,40 a刺槐林下裂葉堇菜、臭草存在地上植物中,為土壤種子庫提供種子,這兩種幼苗在刺槐林恢復中,后期有著較強的更新與存活能力。同時不同林齡刺槐郁閉度、植被密度不相同,也會影響到林下幼苗的生長環境,如林下光照的分布、溫度及土壤水分等隨林齡發生變化[29],對林下幼苗的生長更新有著一定的影響。
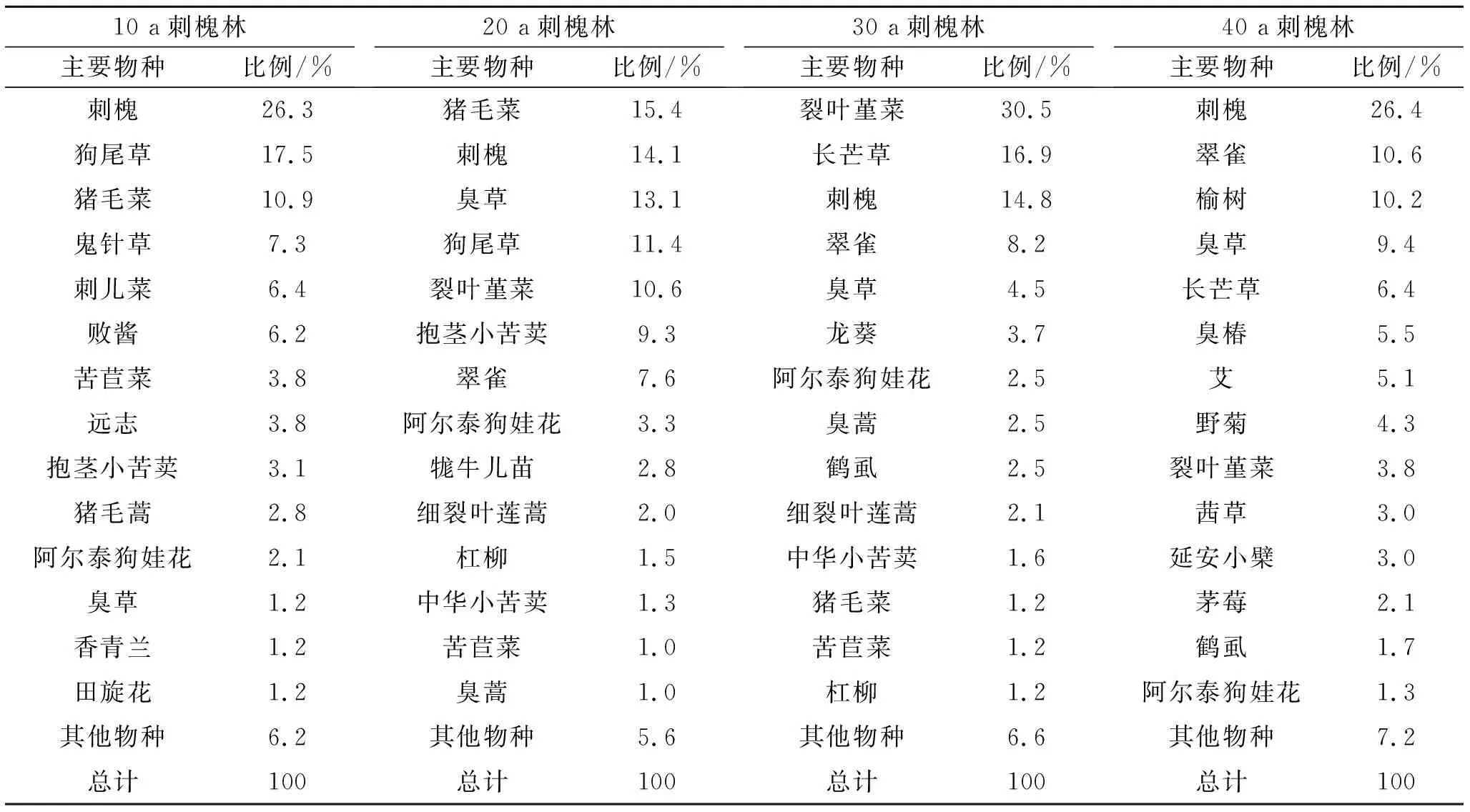
表5 幼苗主要物種及其所占幼苗總數的比例

表6 幼苗物種數月動態變化
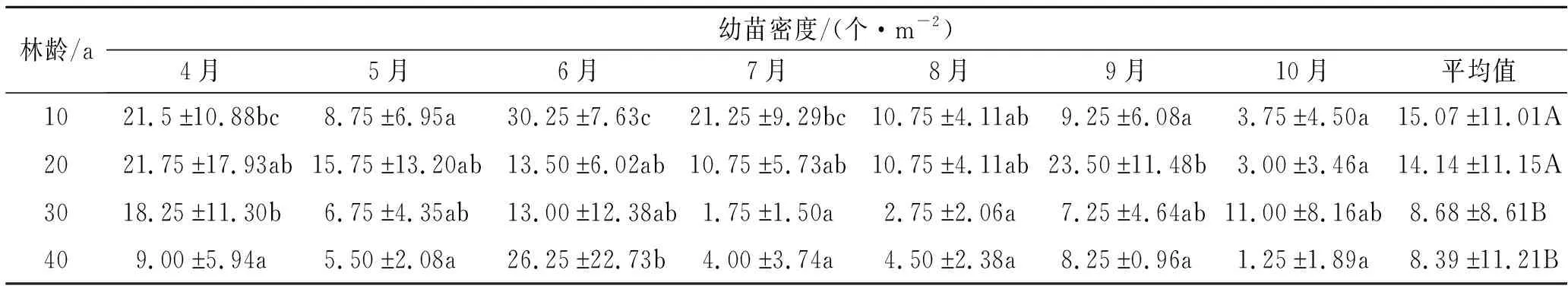
表7 幼苗密度月動態變化
(3) 不同林齡林下幼苗物種數、密度的月動態變化存在差異。幼苗物種數在4月、6月、9月幼苗物種數較高;幼苗密度在4月、6月幼苗密度相對較高。幼苗物種數、密度的動態變化與氣溫、降雨、幼苗特性有著密切關系。安塞4—5月降雨和溫度較低,利于狗尾草、鬼針草等幼苗的萌發[36-37],但低溫、低降雨量也導致豬毛菜、苦苣菜等幼苗大量死亡,因此幼苗物種數、密度均較低;6月溫度、降雨量有所回升,適宜林下幼苗如刺槐、裂葉堇菜、翠雀等萌發生長,幼苗物種數、密度較高;7月、8月氣溫達到高峰,雖然此時降雨量也高,但是地上植被生長旺盛,蒸騰作用強,幼苗之間競爭劇烈,不利于種子萌發和幼苗生長,幼苗物種數和密度較低;9月降雨減小,光照強烈,不利于大部分幼苗的萌發、存活,但也有翠雀、榆樹、阿爾泰狗娃花等幼苗出現,故幼苗物種數較高而幼苗密度較小;10月氣溫、降雨均低,不利于幼苗的萌發與生長,故幼苗物種數、密度較低。不同林齡刺槐林下幼苗物種數、幼苗密度均表現為隨林齡增加而呈減小趨勢,這可能與刺槐林郁閉度有關。林冠郁閉度顯著影響林下光照的分布[38],10 a,20 a刺槐林郁閉度較高,林下光照較弱,降低水分的蒸發,有利于幼苗的萌發,因此幼苗物種數、密度高于30 a,40 a。
(4) 幼苗存活、建植成功是幼苗更新及植物群落更新過程的關鍵環節[39]。本研究發現林下草本植物幼苗大多數存活時間較短,難以成功建植。在刺槐林恢復初期,田旋花幼苗易成功建植;在刺槐林恢復的中、后期,裂葉堇菜、臭草、長芒草幼苗成功建植;且這些建植成功的幼苗物種也存在于地上植物中。因此,在人工刺槐林植被恢復過程中,要營造健康發育的人工林及維持其可持續性,需充分考慮林下幼苗的更新以及是否能夠成功建植,采取相應的林地撫育管理措施,提高幼苗的存活種類與密度,促進人工林的自然恢復,這樣才能有利于該區植被的恢復和生態環境的改善。如在刺槐林恢復前期,應通過調節林地郁閉度、栽植密度改善水熱條件,提高田旋花、豬毛菜、豬毛蒿、鬼針草等以種子繁殖為主的物種種子的萌發率及幼苗的存活能力[40]。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
