化膿隱秘桿菌肝素結合蛋白的提取及鐵結合蛋白的黏附特性分析
2022-03-08 03:08:22沈克飛許國洋付利芝張素輝
畜牧獸醫學報 2022年1期
沈克飛,許國洋,付利芝,楊 柳,牟 豪,張素輝*
(1. 重慶市畜牧科學院, 重慶 402460; 2. 重慶市獸用生物制品工程技術研究中心,重慶 402460)
化膿隱秘桿菌(Trueperellapyogenes)是家養和野生動物化膿性感染的常見病原菌,感染造成組織器官膿腫、化膿性炎癥、纖維化、壞死及流產、死胎等[1]。近年來,化膿隱秘桿菌相關疾病在豬、牛、羊等動物上的發病率逐漸增加,給養殖業帶來較大經濟損失[2-8]。化膿隱秘桿菌是一種革蘭陽性菌,常見于動物上呼吸道、胃腸道和泌尿生殖道黏膜,是動物和人的條件性致病菌[1, 9]。
病原體與宿主硫酸乙酰肝素蛋白多糖(heparan sulfate proteoglycans, HSPGs)的相互作用能增進微生物黏附和侵入宿主細胞,進而促進發病[10-12]。HSPGs是廣泛分布于宿主細胞表面和細胞外基質的一類黏多糖,由與核心蛋白連接的硫酸乙酰氨基葡聚糖(glycosaminoglycans, GAGs)組成。GAGs是一種陰離子線性多糖,由重復的二糖組成,包括肝素、硫酸乙酰肝素等。研究發現,肝素可劑量依賴性地抑制化膿隱秘桿菌黏附HeLa細胞[13],提示化膿隱秘桿菌能利用自身的肝素結合蛋白黏附宿主細胞。最近,利用肝素瓊脂糖凝膠從化膿隱秘桿菌菌體蛋白中提取到包含鐵結合蛋白(iron-binding protein, IBP)在內的多種ATP結合盒(ATP binding cassette, ABC)轉運體的底物結合蛋白(substrate binding proteins, SBPs)。IBP是化膿隱秘桿菌的1種抗原[14],在化膿隱秘桿菌自然感染的山羊血清中能檢測到針對IBP的抗體,提示其在感染中會表達。目前尚無關于IBP參與化膿隱秘桿菌黏附宿主細胞的研究報道。因此,本研究在IBP黏附肝素、宿主細胞及IBP抗血清抑制黏附方面進行了分析,為化膿隱秘桿菌與宿主細胞的相互作用提供了新認識。
1 材料與方法
1.1 材料
1.1.1 細胞、菌株、抗血清 MDBK細胞、山羊源化膿隱秘桿菌、含pGEX-4T-1-IBP重組質粒的BL21(DE3)重組菌株及小鼠抗rIBP血清[14]、GST及其小鼠抗血清、自然感染化膿隱秘桿菌山羊血清[15]由重慶畜牧科學院獸醫研究所保存和提供。
1.1.2 主要試劑 新生胎牛血清、Roswell Park Memoria Institute-1640(RPMI-1640)培養基、胰蛋白酶均購自Hyclone公司;小鼠抗GST單克隆抗體、堿性磷酸酶標記的驢抗山羊IgG抗體或山羊抗小鼠IgG抗體購自EarthOx公司;Gluthathione-Sepharose 4B購自Pharmacia公司;肝素、還原性谷胱甘肽購自Sigma-Alorich公司;BCTP/NBT購自上海生工;肝素瓊脂糖珠購自索萊寶公司;Quick Start Bradford購自BioRad公司。
1.2 肝素結合蛋白的提取與質譜鑒定
化膿隱秘桿菌單菌落接種于含8%小牛血清的TSB培養基,37 ℃振搖培養48 h。7 000 ×g離心10 min,菌體沉淀用 Tris-HCl (0.02 mol·L-1,pH8.0)懸浮,洗3次。每300 mL培養物所得菌體沉淀用20 mL Tris-HCl 懸浮,加入蛋白酶抑制劑。冰水浴中超聲裂解菌懸液(超聲功率為300 W,工作4 s間歇5 s,超聲裂解15 min)。12 000 ×g離心30 min,上清用0.22 μm濾膜過濾,即為菌體蛋白。吸取200 μL充分混懸的肝素瓊脂糖凝膠,用Tris-HCl 洗3次,加入1.2 mL菌體蛋白,4 ℃下搖動孵育2 h,用含0.15 mol·L-1NaCl的Tris-HCl 洗3次, 用含0.5 mol·L-1NaCl的Tris-HCl洗脫。采用SDS-PAGE檢測提取的蛋白質。采用LC-MS/MS方法對肝素瓊脂糖凝膠提取的樣品進行質譜鑒定。
1.3 重組蛋白的制備
質譜鑒定結果顯示所提取的蛋白中存在IBP,按參考文獻[14]所描述的方法制備重組蛋白rIBP。簡單步驟如下:37 ℃下振搖培養至菌體濃度達OD600 nm約為1時,加入終濃度0.1 mmol·L-1IPTG,37 ℃下誘導表達3 h。培養物4 ℃5 000 ×g離心30 min,每100 mL培養液加20 mL PBS以懸浮菌體沉淀,冰水浴中超聲裂解菌懸液(超聲功率為300 W,工作4 s間歇5 s,超聲裂解15 min)。裂解物12 000 ×g離心10 min,向裂解物中加入終濃度1% Trition X-100,4 ℃振搖孵育30 min,5 000 ×g離心30 min,收集上清。每20 mL上清加入300 μL Gluthathione-Sepharose 4B,4 ℃攪動孵育1 h。PBS洗滌3次后用10 mmol·L-1還原性谷胱甘肽洗脫。采用SDS-PAGE分析純化效果,采用透析法去除其中的還原性谷胱甘肽,使用Bradford法定量蛋白質濃度。
1.4 免疫印跡檢測IBP
純化的rIBP經SDS-PAGE后轉印至硝酸纖維素膜上,采用5%脫脂奶粉封閉2 h,TBST洗3次后與自然感染化膿隱秘桿菌山羊血清或rIBP免疫小鼠血清(1∶1 00稀釋)孵育,TBST洗3次后與堿性磷酸酶標記的驢抗山羊IgG抗體或山羊抗小鼠IgG抗體(1∶2 000稀釋)孵育,最后置于BCTP/NBT顯色,以檢測血清中存在抗IBP抗體。
將化膿隱秘桿菌菌體蛋白及肝素瓊脂糖凝膠提取的菌體蛋白經SDS-PAGE后轉印至硝酸纖維素膜上,采用5%脫脂奶粉封閉2 h,TBST洗3次后與自然感染化膿隱秘桿菌山羊血清或小鼠抗rIBP血清(1∶100稀釋)孵育,TBST洗3次后與堿性磷酸酶標記的驢抗山羊IgG抗體或山羊抗小鼠IgG抗體(1∶2 000稀釋)孵育,最后置于BCTP/NBT顯色,以驗證化膿隱秘桿菌菌體蛋白及肝素瓊脂糖凝膠提取的蛋白中存在IBP。
1.5 rIBP肝素結合與結合抑制分析
充分混勻肝素瓊脂糖凝膠,吸取懸浮液30 μL加入離心管中,用預冷的100 μL PBS洗滌3次。向洗好的肝素瓊脂糖凝膠加入30 μL 0.3 mg·mL-1rIBP,顛倒混勻,4 ℃下緩慢搖動孵育1 h。4 ℃下3 000 ×g離心5 min,肝素瓊脂糖凝膠沉淀用200 μL 預冷NaCl溶液洗3次,NaCl濃度依次為0.025、0.05、0.1、0.2、0.3、0.4、0.5 mol·L-1。向凝膠加入40 μL SDS-PAGE上樣緩沖液,沸水浴10 min,采用SDS-PAGE分析rIBP與肝素的結合情況。
用PBS溶液溶解肝素配制肝素溶液。按表1加入各組分,按肝素濃度分為4個處理組、肝素濃度依次為0、10、30、90 mg·mL-1。混勻,4 ℃下緩慢搖動孵育1 h。4 ℃下3 000×g離心10 min,保留沉淀。按上述方法進行免疫印跡分析,一抗為小鼠抗GST單克隆抗體(1∶2 000稀釋),二抗為堿性磷酸酶標記的山羊抗小鼠IgG抗體(1∶2 000稀釋)。

表1 肝素結合抑制分析
rIBP和GST抗血清用PBS進行1∶10稀釋。將肝素瓊脂糖凝膠、rIBP(0.3 mg·mL-1)、稀釋的rIBP或GST抗血清加入1.5 mL離心管中,各組分均為60 μL,混勻,4 ℃下緩慢搖動孵育1 h。4 ℃下3 000 ×g離心10 min,凝膠用0.1 mL 預冷0.9%NaCl洗3次。向凝膠加入40 μL SDS-PAGE上樣緩沖液,沸水浴10 min,采用免疫印跡分析抗血清對rIBP結合肝素的抑制作用,一抗為小鼠抗GST單克隆抗體(1∶2 000稀釋),二抗為堿性磷酸酶標記的山羊抗小鼠IgG抗體(1∶2 000稀釋)。
1.6 rIBP細胞黏附分析
用胰酶(0.05%胰酶,0.02% EDTA)消化長滿單層的MDBK細胞,用RPMI-1640洗3次,將細胞制備成1010個·mL-1備用。將蛋白質(GST、rIBP)的濃度調整為0.3 mg·mL-1。按表2加入各組分,按蛋白質濃度分為4個處理組,蛋白質濃度依次為0、0.015、0.045 和0.135 mg·mL-1。顛倒混勻各組分,4 ℃下緩慢搖動孵育1 h。4 ℃下800 r·min-1離心10 min, 沉淀用1 mL 預冷PBS洗3次。樣本經SDS-PAGE后轉印至硝酸纖維素膜上,用5%脫脂奶粉封閉2 h,TBST洗3次后與小鼠抗GST單克隆抗體(1∶2 000稀釋)孵育,TBST洗3次后與堿性磷酸酶標記的山羊抗小鼠IgG(1∶2 000稀釋)孵育,最后置于BCTP/NBT顯色。

表2 rIBP細胞黏附分析
1.7 rIBP抗血清抑制細菌黏附細胞分析

2 結 果
2.1 肝素結合蛋白的提取與質譜鑒定
SDS-PAGE顯示,利用肝素瓊脂糖凝膠從化膿隱秘桿菌菌體蛋白中提取到蛋白,提取后蛋白質條帶數量明顯減少,部分條帶得到明顯富集(圖1A、B)。蛋白質質譜鑒定得出,所提取的蛋白包含IBP等多種ABC轉運體SBP(表3)。
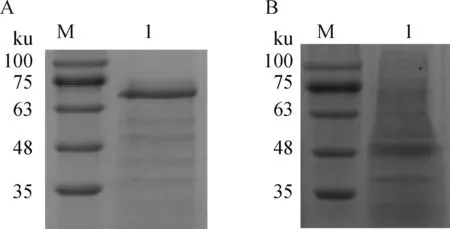
A.肝素瓊脂糖凝膠提取的蛋白;B. 菌體蛋白;M. 蛋白質相對分子質量標準;1. 蛋白質樣品A. Proteins extracted by heparin agarose gel; B. Bacterial proteins; M. Protein marker; 1. Protein sample圖1 SDS-PAGE分析肝素瓊脂糖凝膠提取的化膿隱秘桿菌蛋白質Fig.1 SDS-PAGE analysis of the proteins extracted from Trueperella pyogenes by heparin agarose gel
2.2 rIBP的制備
采用Gluthathione-Sepharose 4B從37 ℃下0.1 mmol·mL-1IPTG誘導表達3 h的重組菌株中純化得到rIBP(圖2)。

M.蛋白質相對分子質量標準;1. rIBPM.Protein marker; 1. rIBP圖2 rIBP的純化結果Fig.2 Purification of rIBP
2.3 免疫印跡檢測IBP
采自自然感染化膿隱秘桿菌山羊血清或免疫rIBP后的小鼠血清,在免疫印跡中能識別rIBP(圖3A),表明血清中含有抗rIBP抗體。自然感染山羊血清識別rIBP也表明在感染過程中化膿隱秘桿菌表達IBP。
免疫印跡顯示,肝素瓊脂糖凝膠提取的蛋白能被自然感染化膿隱秘桿菌的山羊血清識別出2條帶(圖3B),能被 rIBP小鼠抗血清識別1條帶(圖3C)。 rIBP小鼠抗血清所識別的條帶和自然感染的山羊血清所識別的其中1條帶的大小一致,表明化膿隱秘桿菌在含8%小牛血清的TSB培養基中37 ℃振搖培養有IBP表達,且IBP能用肝素瓊脂糖凝膠從化膿隱秘桿菌培養物中提取。

A.抗血清與rIBP的反應性;B. 自然感染化膿隱秘桿菌山羊血清識別的化膿隱秘桿菌蛋白;C. 小鼠抗rIBP血清識別的化膿隱秘桿菌蛋白;M. 蛋白質相對分子質量標準;1. 自然感染化膿隱秘桿菌山羊血清;2.小鼠抗 rIBP血清;3. 肝素瓊脂糖凝膠提取的蛋白;4.菌體蛋白A.The reaction between antiserum and rIBP; B. Proteins of T. pyogenes recognized by serum of goat infected naturally with T. pyogene; C. Proteins of T. pyogenes recognized by mice anti-rIBP serum; M. Protein marker; 1. Serum of goat naturally infected with T. pyogenes; 2. Anti-rIBP serum from mice; 3.Proteins extracted by heparin agarose gel; 4.Bacterial proteins圖3 抗血清識別的化膿隱秘桿菌蛋白(免疫印跡)Fig.3 Proteins of T. pyogenes recognized by the antiserum (immunoblotting)
2.4 肝素結合與結合抑制分析
SDS-PAGE顯示,隨著洗液中NaCl濃度從0.025 mol·L-1逐漸提高,肝素瓊脂糖珠結合的rIBP的量逐漸下降,至NaCl達到0.5 mol·L-1時,基本不可見rIBP;同時,GST沒有結合肝素瓊脂糖珠(圖4A)。這表明rIBP結合肝素瓊脂糖珠與標簽GST無關,且結合能被生理濃度的NaCl部分解離下來。
肝素與肝素瓊脂糖凝膠、rIBP共孵育后,免疫印跡中的特異條帶亮度隨肝素濃度升高而降低,表現出劑量依賴關系(圖4B),表明游離肝素與瓊脂糖凝膠中固相肝素競爭結合rIBP,對結合的抑制作用是特異的。
免疫印跡顯示,與PBS相比,rIBP、GST抗血清都能抑制rIBP結合肝素瓊脂糖凝膠,且與兩者孵育后肝素瓊脂糖凝膠結合的rIBP量無明顯差異(圖4C),表明針對rIBP的抗體不能抑制rIBP結合肝素,抗血清對結合的抑制作用是由血清中的其他物質造成的。
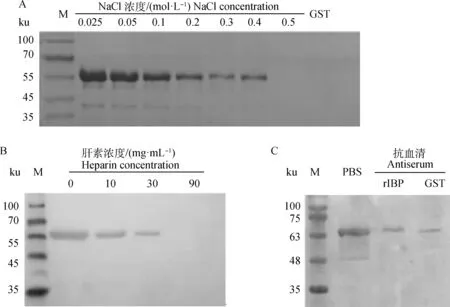
A. rIBP結合肝素瓊脂糖凝膠分析;B. 肝素抑制rIBP結合肝素瓊脂糖凝膠;C. 抗血清抑制 rIBP結合肝素瓊脂糖凝膠(rIBP. 小鼠抗 rIBP血清;GST. GST抗血清);M. 蛋白質相對分子質量標準A. Binding assay of rIBP to heparin agarose gel; B. Heparin inhibits rIBP binding to heparin agarose gel; C. Antiserum inhibits rIBP binding to heparin agarose gel (rIBP. Anti-rIBP serum from mice; GST. Anti-GST serum); M. Protein marker圖4 rIBP肝素結合與結合抑制分析Fig.4 Analysis of rIBP binding heparin and the binding inhibition
2.5 細胞黏附分析
免疫印跡顯示,rIBP與MDBK細胞孵育后,出現特異條帶,且條帶亮度隨rIBP劑量的增加而增量,表現出良好的劑量依賴關系;GST與MDBK細胞孵育后未出現明顯條帶(圖5),表明rIBP黏附MDBK細胞與GST標簽無關,能特異性黏附MDBK細胞。
2.6 rIBP抗血清抑制細菌黏附細胞分析
抗血清抑制細胞黏附試驗結果顯示,與rIBP或GST抗血清(1∶10稀釋)預孵育的化膿隱秘桿菌和與PBS預孵育的化膿隱秘桿菌相比,黏附到MDBK細胞上的數量下降極顯著(P<0.05);與rIBP抗血清預孵育的化膿隱秘桿菌和與GST抗血清預孵育的化膿隱秘桿菌黏附到MDBK細胞上的數量下降極顯著(P<0.05)(圖6)。這表明rIBP抗血清能抑制化膿隱秘桿菌黏附到MDBK細胞,進一步證實rIBP能黏附MDBK細胞。

M.蛋白質相對分子質量標準M. Protein marker圖5 rIBP黏附MDBK細胞分析(免疫印跡)Fig.5 Adhesion assay of rIBP to MDBK cells (Western blotting)
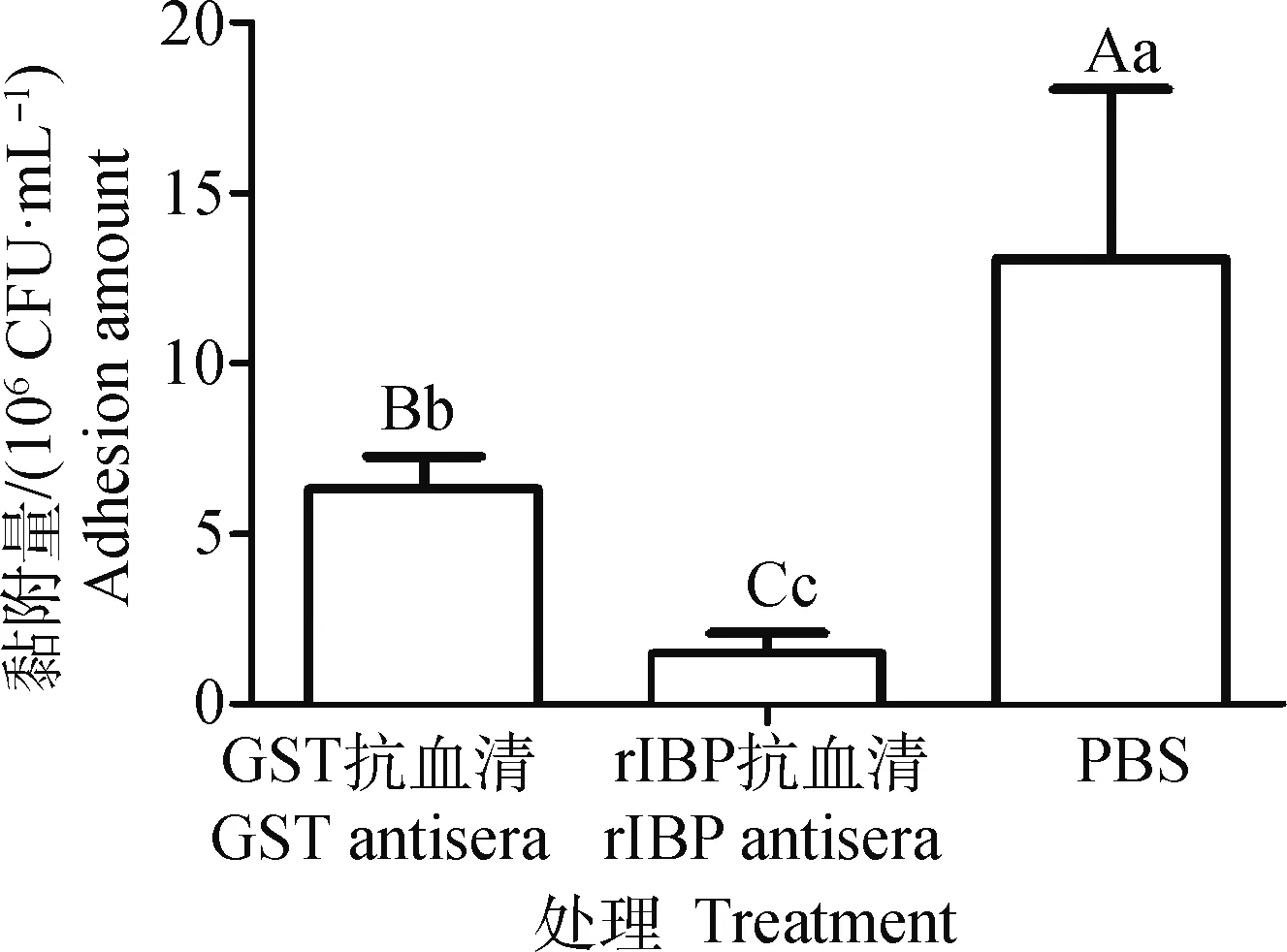
圖中不同大寫字母(A、B、C)表示差異極顯著(P<0.01),不同小寫字母(a、b、c)表示差異顯著(P<0.05)Different capital letters (A, B or C) mean P<0.01, different lower letters (a, b or c) mean P<0.05 in the column chart圖6 rIBP抗血清抑制化膿隱秘桿菌黏附細胞的作用分析Fig.6 Inhibitory effect of anti-rIBP serum on cell adhesion of T. pyogenes
3 討 論
肝素、硫酸乙酰肝素是結構類似的GAGs[16]。硫酸乙酰肝素在多細胞生物進化的早期出現,與核心蛋白共價結合,是所有多細胞生物細胞膜結構的基本成分[17]。據計算人肝細胞表面表達4×105個硫酸乙酰肝素分子[18]。在感染中,很多病原體利用自身的表面蛋白黏附宿主HSPGs,以使自身附著靶細胞并侵入組織[10-12]。這些研究結果指出很多病原在進化中適應了以HSPGs作為受體的一般策略。
病原體表面蛋白與HSPGs黏附的分子基礎是,這些表面蛋白結構域柔性環中的富含Arg、Lys區域負責與HSPGs結合,蛋白質上帶正電荷的氨基酸簇與HSPGs上帶負電荷的GAGs形成離子鍵,在黏附基序上要求帶正電荷的氨基酸簇與疏水性氨基酸相靠近,疏水性氨基酸側鏈和糖鏈非極性基團間的相互作用穩定了蛋白質和聚糖的結合[19-20]。與HSPGs結合的病原體表面蛋白能通過肝素瓊脂糖凝膠層析從膜蛋白中提取[20]。之前用商品化細菌膜蛋白提取試劑盒提取的化膿隱秘桿菌膜蛋白是高鹽溶液[14],高濃度鹽會降低蛋白質與肝素瓊脂糖凝膠的結合力。本研究利用肝素瓊脂糖凝膠從化膿隱秘桿菌裂解物上清提取蛋白,從SDS-PAGE和免疫印跡分析結果來看,提取后蛋白質條帶數量明顯減少,部分條帶得到明顯富集。經蛋白質質譜和免疫印跡鑒定,提取到包含IBP在內的10種ABC轉運體的SBPs,表明化膿隱秘桿菌的這些蛋白質在含8%小牛血清的TSB培養基中37 ℃振搖培養有大量表達。
ABC轉運體負責細胞內外物質攝取、轉運,由ABC型膜透酶(permease)、ATP結合蛋白質(或稱ATP酶,ATPase)和底物結合蛋白(substrate binding protein, SBP)組成,不同類型的ABC轉運體轉運不同的底物。SBPs是一類脂蛋白(lipoproteins)[21]。細菌脂蛋白含有N-末端脂質修飾,即N-酰基-S-二酰基-甘油基-半胱氨酸(N-acyl-S-diacyl-glyceryl-cysteine)。脂蛋白在原核生物中含量豐富。Stoll等[22]利用脂蛋白搜索程序ParSeq[23]對金黃色葡萄球菌N315基因組進行了分析,鑒定出70多個推測的脂蛋白,但只有55個含有正確長度的信號肽,35個脂蛋白可被注釋為鐵、鋅、氨基酸、寡肽、甘氨酸甜菜堿、糖和磷壁酸的轉運蛋白。
在革蘭陽性細菌中,脂蛋白錨定在細胞質膜的外表面上,并可延伸到細胞壁內外,與外部成分(如營養素和宿主蛋白質)結合[21],是一類保守的細菌黏附蛋白家族[24-25]。在免疫印跡中,自然感染化膿隱秘桿菌的山羊血清能識別出rIBP,這表明IBP能在感染過程中表達,可能在感染過程中發揮黏附宿主細胞的作用。IBP(rIBP)能結合肝素和宿主細胞,表明IBP可能通過與宿主HSPGs結合而促進化膿隱秘桿菌黏附宿主細胞。另外,與肝素瓊脂糖凝膠結合的rIBP能被生理濃度的NaCl部分解離下來,提示IBP與HSPGs結合可能涉及化膿隱秘桿菌在黏膜定植和感染部位上與宿主細胞的動態相互作用,而與宿主細胞的動態相互作用對病原菌的體內存活和疾病的發展至關重要[24]。rIBP能結合肝素和宿主細胞,但rIBP抗血清不能明顯抑制其結合肝素而能抑制其黏附宿主細胞,表明IBP還有其他宿主受體。
4 結 論
采用肝素瓊脂糖凝膠從化膿隱秘桿菌裂解物中提取到IBP等多種蛋白,證實在自然感染中化膿隱秘桿菌表達IBP,IBP特異性結合肝素、黏附MDBK細胞,rIBP抗血清不能明顯抑制rIBP結合肝素,但能抑制化膿隱秘桿菌黏附MDBK細胞。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
