舟山群島東側海域蟹類群落結構及其與環境因子的關系
2022-02-18 12:58:02牛威震韓磊俞存根鄭基許永久顏文超
海洋學報 2022年2期
牛威震,韓磊,俞存根*,鄭基,許永久,顏文超
(1.浙江海洋大學 水產學院,浙江 舟山 316022)
1 引言
舟山群島地處長江口與杭州灣入海口,其漁場環境受臺灣暖流、浙江沿岸流和黃海冷水團的交互影響,水質肥沃,具有較高的初級生產力水平和豐富的餌料生物,是眾多魚類、蝦蟹類等生物優良的產卵場和索餌場。蟹類是海洋無脊椎動物的重要類群之一,從潮間帶到大陸架淺海以及大洋深海底均有分布,它不僅是海洋生物群落中的重要生物類群,很多種類還是人類自古以來的重要捕撈對象。東海的蟹類資源十分豐富,根據俞存根等[1]研究可知,東海分布的蟹類有324 種,可食用蟹類約有20 種。對東海及舟山漁場的蟹類資源研究報道已有不少,如沈嘉瑞和劉瑞玉[2]、戴愛云等[3]對東海蟹類的分類做過研究,俞存根等[4-12]對東海蟹類種類組成、區系特征、數量分布、群落結構特征進行了報道,并研究了細點圓趾蟹(Ovalipes punctatus)、日本蟳(Charybdis japonica)、銹斑蟳(Charybdisferiatus)、武士蟳(Charybdis miles)等主要經濟蟹類的生物學特性,評估了東海蟹類的資源量等。薛利建和盧占暉[13]借助生物多樣性分析了東海中部蟹類群落結構特征;陳小慶等[14]通過劃分生態群落研究了東海中南部蟹類群落結構與環境因子之間的關系;張洪亮等[15]分析了浙江南部近岸蟹類春季群落結構特征,并與東海海域的生物多樣性和優勢種等進行了比較;盧衎爾等[16]研究報道了浙江近海蟹類群落結構與環境因子的關系。舟山漁場的蟹類資源,主要集中在舟山沿岸海域,如劉惠等[17-18]在研究舟山沿岸海域甲殼類群落結構特征、生態位及其與種間聯結性時有所涉及。本文根據2018 年4 月(春季)和10 月(秋季)在舟山群島東側海域開展漁業資源底拖網調查所獲得的蟹類數據資料,開展調查海域蟹類資源種類組成、優勢種、生物多樣性等群落結構特征,同時分析了蟹類數量分布及其群落結構與溫、鹽等環境因子之間的關系。因關于舟山群島東側海域蟹類群落分析的文章較少,希望對本研究區域蟹類群落結構,特別是舟山蟹類群落結構與舟山及長江口毗連海域生態系統關系的分析可作為基礎研究資料進行分享。本研究可進一步豐富該海域蟹類群落生態學資料,為蟹類資源的合理利用和科學管理提供科學依據。
2 材料與方法
2.1 數據來源
本文所用數據采于2018 年4 月(春季)和10 月(秋季)在舟山群島東側海域開展的漁業資源底拖網調查所獲得的蟹類資料,調查共設置16 個站位,調查范圍為29°10′25.54′′~30°45′0.00′′N,122°53′3.88′′~124°48′28.31′′E,調查船為“浙漁科2”科考船,主機功率為1 007 kW,漁船有配備衛星導航設備、探測儀(測深儀)、甚高頻VHF 以及其他助漁助航設備,拖網網具規格為1 020 目×80 mm。調查時每站位作業時拖拽約1 h,拖速約為3.5 kn,將每個調查站位所捕獲的漁獲物全部裝入樣品袋,并記錄站位號和樣品袋號,冰鮮保存后帶回實驗室進行生物學測定。測重儀器為電子天平,精確度為0.1 g。調查方法按《海洋漁業資源調查規范》(SC/T 9403-2012)執行。溫度、鹽度及水深調查測量與漁業資源底拖網調查同步進行,溫度、鹽度及水深等環境因子數據采用溫鹽深儀(Conductance Temperature Depth,CTD)同步獲取,調查站位設置如圖1 所示。
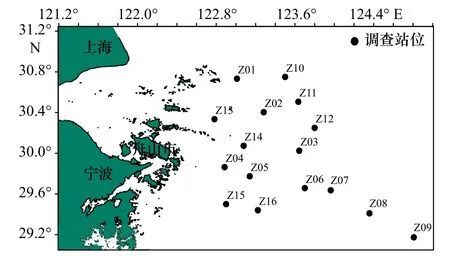
圖1 舟山群島東側海域漁業資源調查站位分布Fig.1 Distribution of fishery resources survey stations in the eastern sea area of the Zhoushan Islands
2.2 數據處理
2.2.1 優勢種計算公式
蟹類優勢種采用相對重要性指數(Index of Relative Importance,IRI)來確定,其計算公式為
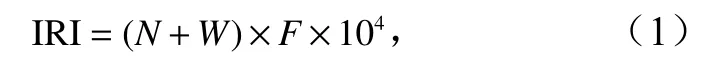
式中,N和W分別代表為某種蟹類的尾數及重量占蟹類總尾數及總重量的百分比;F為某種蟹類出現的站位數占調查總站位數的百分比。IRI 大于1 000 的視為優勢種,100~1 000 的視為常見種[19]。
2.2.2 生物多樣性分析
采用Shannon-Wiener 多樣性指數(H′)、Pielou 均勻度指數(J′)、Margalef 豐富度指數(D)來計算分析調查海域的蟹類生物多樣性,公式分別為
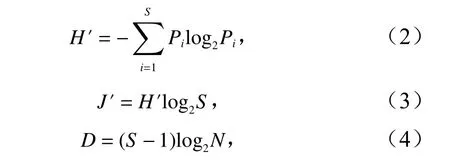
式中,Pi是第i種蟹類生物量占總生物量的比例;S為蟹類總種數;N為蟹類總尾數。
2.2.3 群落結構相似性分析
本文運用Bray-Curtis 相似性系數,并采用Cluster等級聚類和非度量多維標度分析(Non-Metric Multi-Dimensional Scaling,NMDS)方法分析調查海域蟹類組成特征。計算公式為
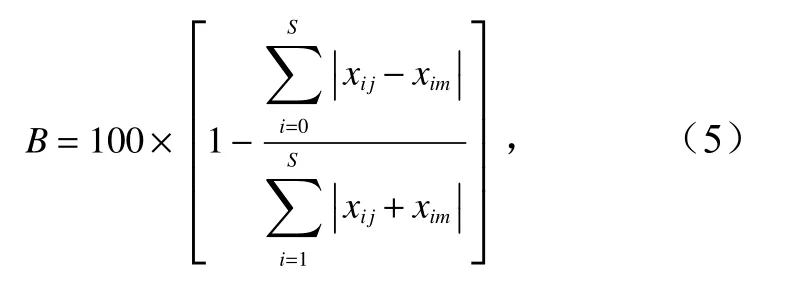
式中,B表示Bray-Curtis 相似性系數;S表示種類數;xij表示第i物種在j站位中的漁獲重量;xim表示第i物種在m站位中的漁獲重量。以脅迫系數衡量NMDS 二維點陣分布效果,通常認為脅迫系數小于0.05時,具有很好的代表性;脅迫系數為 [0.05,0.1) 時,結果基本可信;脅迫系數為 [0.1,0.2) 時,具有一定解釋意義。
2.2.4 種類更替率
以種類更替率分析蟹類種類的季節更替,公式為

式中,A為更替率,表示與上一個季節比較的更替情況;C為兩季節間種類增加數或減少數;N為兩季節間相同種類數。
2.2.5 典范對應分析
采用典范對應分析(Canonical Correspondence Analysis,CCA)方法分析蟹類群落與環境因子之間的關系,具體計算利用軟件CANOCO 5.0,排序結果用站位-環境因子關系雙序圖表示。
本文蟹類生態習性特征參考文獻[20]的相關研究;蟹類種名錄和種類更替率參照文獻[21-22]的研究報道;調查站位圖和蟹類數量分布圖采用Surfer 11.0 軟件繪制,生物多樣性分析采用Excel 2003 和Primer 5.0 軟件完成,春、秋季漁獲量之間顯著性檢驗使用SPSS 21.0 完成。
3 結果
3.1 種類組成及優勢種
根據2018 年4 月(春季)與10 月(秋季)兩個航次底拖網調查取樣樣品,共鑒定出蟹類40 種,隸屬于12 科23 屬,其中,春季29 種,隸屬于9 科18 屬,秋季35 種,隸屬于11 科21 屬。物種組成上,兩季均以梭子蟹科種類數最多(春、秋季均為12 種),其次是玉蟹科(春、秋季均為5 種),春、秋季蟹類種類數更替率為20%。蟹類生物種類組成如表1 所示。
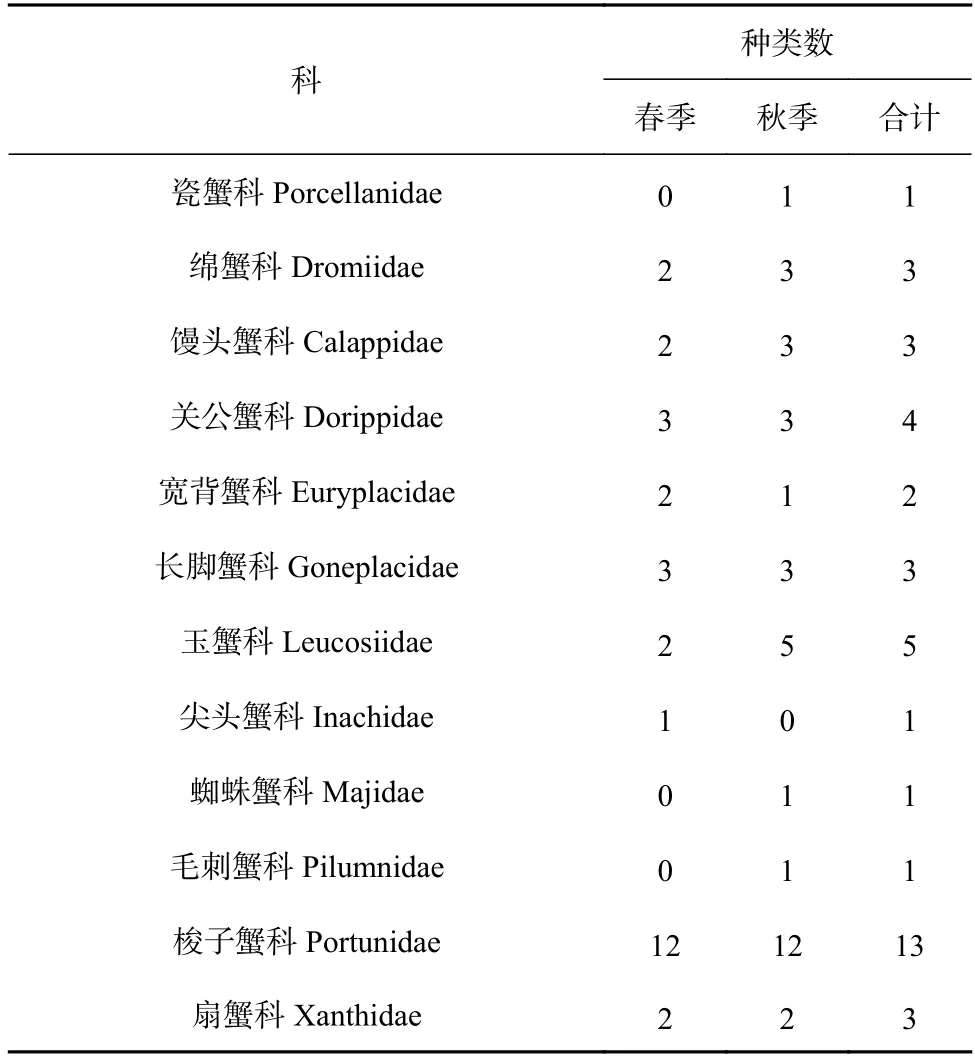
表1 調查海域春秋季蟹類種類數的組成Table 1 The composition of crab species in spring and autumn in the investigated sea area
蟹類優勢種組成如表2 所示,春季優勢種為雙斑蟳(Charybdis bimaculata),常見種有日本英雄蟹(Achaeusjaponicus)、綿蟹(Dromia dehaani)、日本蟳、銀光梭子蟹(Portunus argentatus)、長手隆背蟹(Carcinoplax longimana)、三疣梭子蟹(Portunus trituberculatus)6 種;秋季優勢種為雙斑蟳和三疣梭子蟹,常見種為日本蟳。從生態習性上來看,調查海域春、秋季蟹類區系特征主要為廣溫、廣鹽性的暖水種。
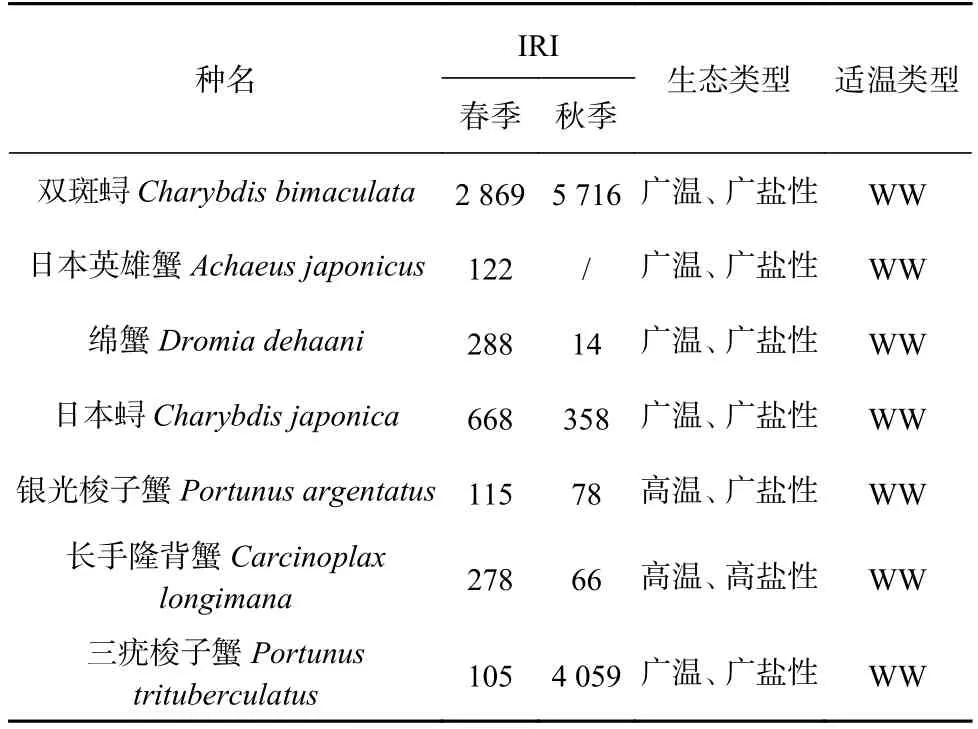
表2 調查海域主要蟹類相對重要性指數及生態習性Table 2 The relative importance index and ecological habits of the main crabs in the investigated sea area
3.2 生物多樣性
調查海域附近各站位春、秋季蟹類生物多樣性及平面分布如表3、圖2 至圖4 所示。

表3 調查海域各站位蟹類生物豐富度指數(D)、均勻度指數(J')和多樣性指數(H')Table 3 Richness index (D),evenness index (J') and diversity index (H′) of crab in each station in the investigated sea area
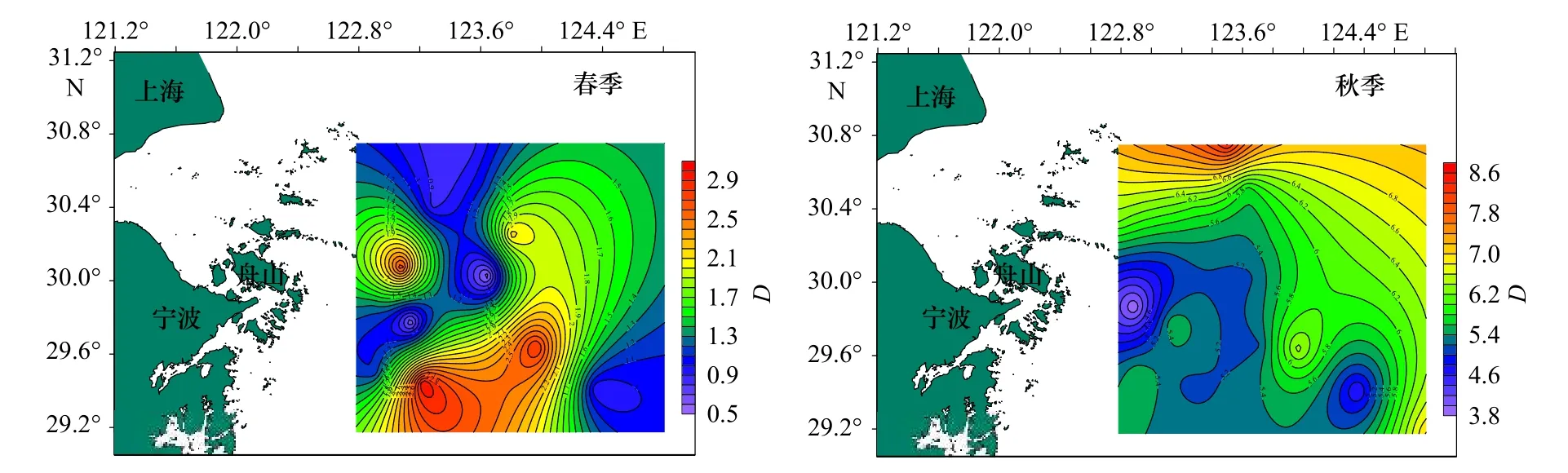
圖2 調查海域蟹類豐富度指數(D)的平面分布Fig.2 Plane distribution of crab richness index (D) in the investigated sea area
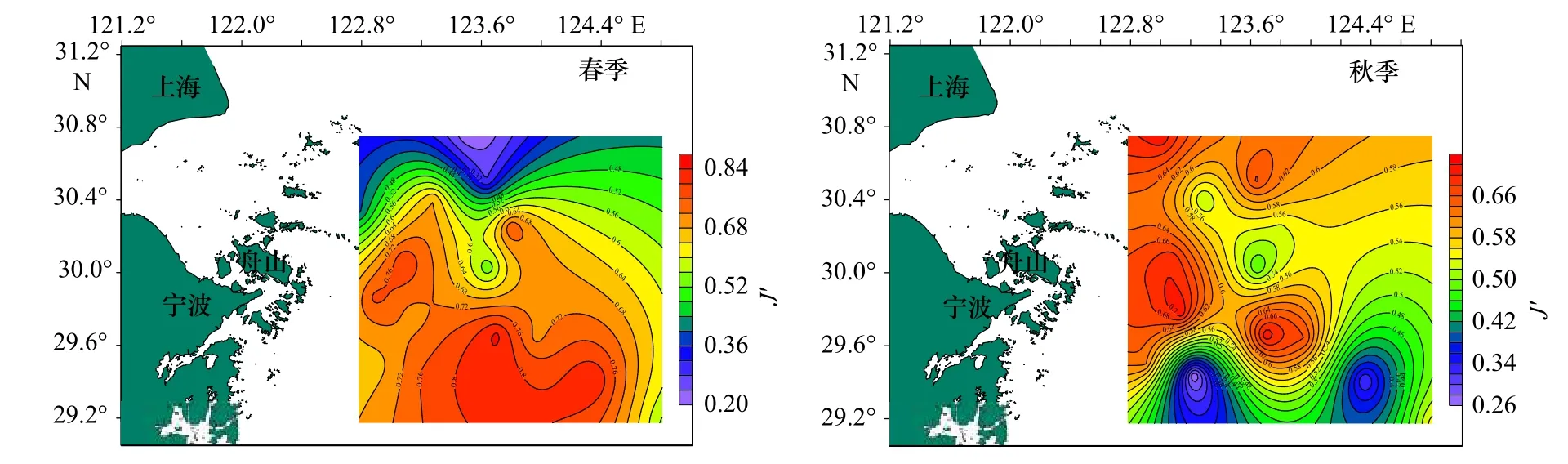
圖3 調查海域蟹類均勻度指數(J')的平面分布Fig.3 Plane distribution of crab evenness index (J') in the investigated sea area
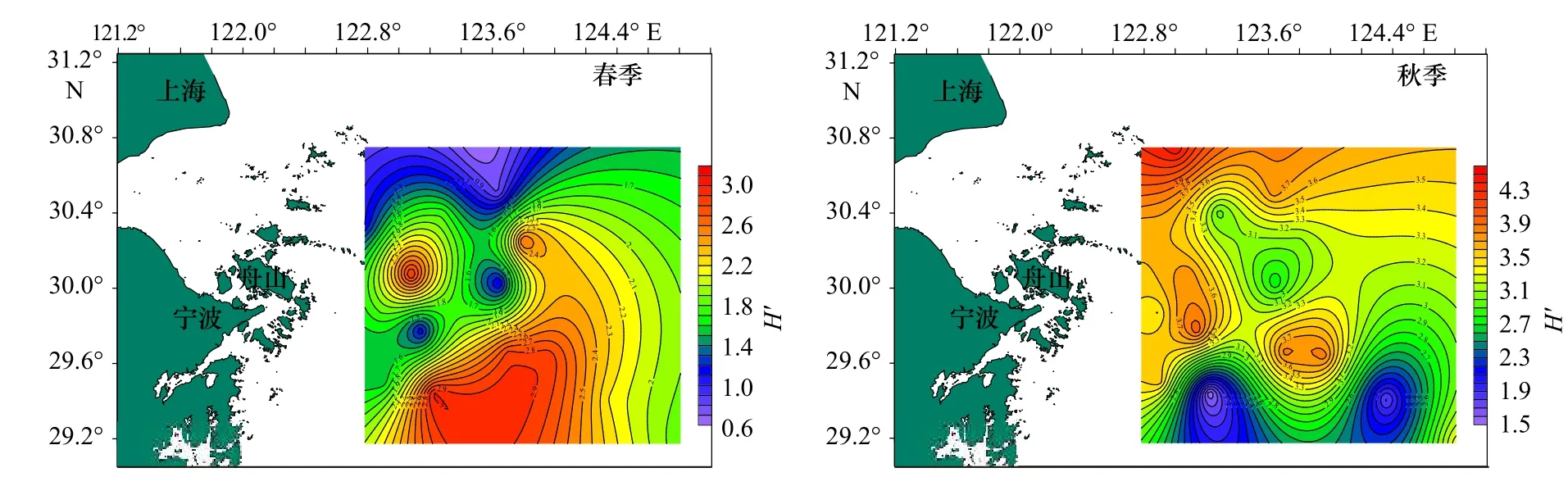
圖4 調查海域蟹類多樣性指數(H')的平面分布Fig.4 Plane distribution of crab diversity index (H') in the investigated sea area
從表3 中可以看出,春季,蟹類種類豐富度指數(D)分布在0.54~3.01 之間,平均值為1.51,以Z16 站位最高,Z03 站位最低;均勻度指數(J')分布在0.21~0.85 之間,平均值為0.63,以Z06 站位最高,Z10 站位最低;生物多樣性指數(H')分布在0.60~3.02 之間,平均值為1.87,以Z16 站位最高,Z10 站位最低。
秋季,蟹類種類豐富度指數(D)分布在3.82~8.42 之間,平均為5.81,以Z10 站位最高,Z04 站位最低;均勻度指數(J')分布在0.26~0.72 之間,平均為0.58,以Z06 站位最高,Z16 站位最低;多樣性指數(H')分布在1.51~4.46 之間,平均值為3.34,以Z01 站位最高,Z16 站位最低;總的來看,春、秋季調查海域不同站位蟹類的指數差異較大,且兩個季節的不同指數最高值和最低值分別出現在不同站位。
總體來說,春、秋季生物指數平面分布季節變化明顯,秋季3 項指數較高處比春季較高處均有向北部移動跡象,并且3 項指數均為北部海域高于南部海域。比較春、秋兩季的生物指數,除均勻度相差不大外,豐富度及多樣性均表現為秋季大于春季。
3.3 時空分布
春、秋季調查海域蟹類每小時漁獲重量分布如圖5所示。春季,調查海域各站位平均每小時蟹類重量漁獲范圍為132.6~10 875.2 g/h,平均漁獲率為2 015.1 g/h,最高漁獲率出現在Z09 站位,為10 875.2 g/h,Z07 站位、Z12 站位、Z10 站位等依次遞減。其余站位每小時蟹類漁獲重量均低于調查海域的平均值,Z05 站位最低,僅為132.6 g/h。秋季,調查海域各站位平均每小時蟹類重量漁獲范圍為2 619.1~32 382 g/h,平均漁獲率為12 676.3 g/h,最高漁獲率出現在Z16 站位,為32 382 g/h,Z01 站位、Z03 站位、Z14 站位、Z08 站位和Z05 站位等依次遞減,其余站位每小時魚類漁獲重量均低于調查海域的平均值,其中以Z10 站位最低,僅為2 619.1 g/h。
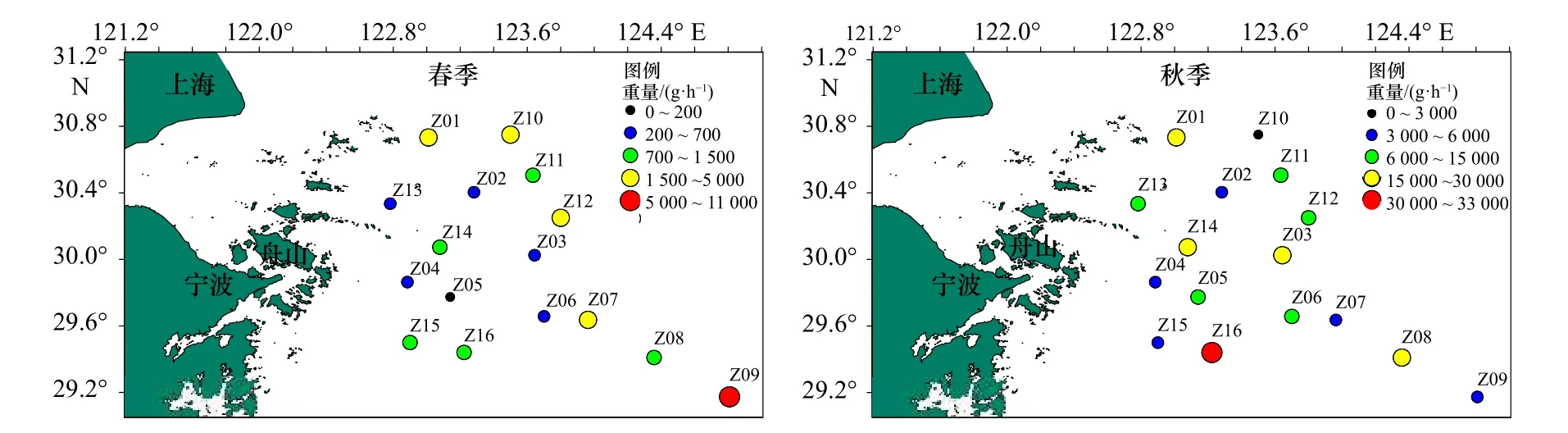
圖5 調查海域各站位蟹類重量分布Fig.5 Weight distribution of crab at different stations in the investigated sea area
春、秋季調查海域蟹類每小時漁獲尾數分布如圖6 所示。春季,調查海域各站位平均每小時蟹類尾數漁獲范圍為15~4 784 ind./h,平均漁獲率為502 ind./h,最高漁獲率出現在Z09 站位,為4 784 ind./h,其次是Z10 站位,為1 550 ind./h,其余站位平均每小時蟹類漁獲重量均低于調查海域的平均值,其中以Z01 站位最低,僅為15 ind./h。秋季,調查海域各站位平均每小時蟹類尾數漁獲范圍為335~4 373 ind./h,平均漁獲尾數為1 147 ind./h,最高漁獲率出現在Z12 站位,為4 373 ind./h,漁獲率較高的還有Z01 站位、Z03 站位、Z11 站位、Z05 站位、Z04 站位和Z15 站位,其余站位平均每小時蟹類漁獲尾數均低于調查海域平均值,其中以Z04 站位最低,僅為335 ind./h。
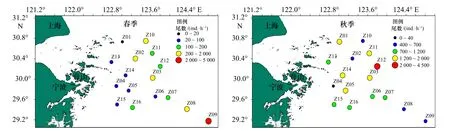
圖6 調查海域各站位蟹類尾數分布Fig.6 Mantissa distribution of crab at different stations in the investigated sea area
3.4 聚類、排序分析
蟹類群落Cluster 聚類分析結果和NMDS 二維點陣圖排序結果如圖7 所示,春季蟹類群落Cluster聚類分析顯示,各調查站位在35%相似性水平上可被分為4 個群組,第一群組為Z06 站位、Z07 站位、Z15站位、Z12 站位、Z14 站位和Z16 站位,代表性蟹類為雙斑蟳、長手隆背蟹及日本英雄蟹,第二群組有Z04 站位、Z05 站位、Z11 站位和Z13 站位,代表性蟹類是雙斑蟳和日本蟳,第三群組有Z02 站位、Z03 站位、Z08 站位、Z10 站位和Z09 站位,代表性蟹類是日本英雄蟹和雙斑蟳,第四群組僅Z01 站位,該站位僅捕獲到細點圓趾蟹1 種,數量僅有15 尾,由此可見,春季以雙斑蟳分布最為廣泛。
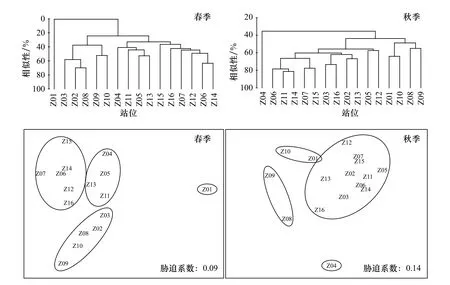
圖7 蟹類群落Cluster 聚類分析和非度量多維標度分析排序Fig.7 Cluster analysis and non-metric multi-dimensional scaling ordination of crab community
秋季,各調查站位在50%相似性水平上也可被分為4 個群組,第一群組為Z08 站位和Z09 站位,代表性蟹類是三疣梭子蟹和銀光梭子蟹,第二群組為Z01站位和Z10 站位,代表性蟹類是雙斑蟳,第三群組為Z04 站位,代表性蟹類是三疣梭子蟹,第四群組有Z02 站位、Z06 站位和Z16 站位等11 個站位,代表性蟹類是雙斑蟳和三疣梭子蟹,可見三疣梭子蟹在秋季分布最為廣泛。秋季蟹類群落相似性高于春季,兩季各群落代表性蟹類均有所不同。通過非度量多維標度分析結果可知,調查海域春季NMDS 脅迫系數為0.09,介于0.05~0.10 之間,說明其結果基本可信;秋季NMDS 脅迫系數為0.14,介于0.10~0.20 之間,說明其結果具有解釋意義。
SIMPER 分析顯示,春季對相似性貢獻率較大的種類為雙斑蟳、日本蟳和長手隆背蟹,貢獻率分別為47.1%、17.7%、6.3%;秋季對相似性貢獻率較大的種為雙斑蟳、三疣梭子蟹和日本蟳,貢獻率分別為45.7%、27.4%、10.7%。春、秋季蟹類群落相異性為65.5%,對相異性貢獻率較大的分歧種為三疣梭子蟹,貢獻率為14.2%,其余蟹類相異性貢獻率均低于8%。
3.5 環境因子結果
根據同步調查所獲得的溫度、鹽度數據,調查海域春季表層水溫分布范圍為14.96~18.05℃,平均為16.59℃,秋季表層水溫分布范圍為21.89~24.18℃,平均為22.89℃,秋季高于春季;調查海域春季底層水溫分布范圍為14.00~18.44℃,平均為16.87℃,秋季底層水溫分布范圍為21.42~23.36℃,平均為22.34℃,秋季高于春季;調查海域春季表層鹽度分布范圍為30.16~32.72,平均為31.51,秋季表層鹽度分布范圍為32.89~33.98,平均為33.44,春季與秋季相差不大;調查海域春季底層鹽度分布范圍為31.69~34.16,平均為33.75,秋季底層鹽度分布范圍為33.15~34.36,平均為33.81,平均底層鹽度春季與秋季基本持平。
將調查海域蟹類群落結構與表層水溫(ST)、底層水溫(BT)、表層鹽度(SS)、底層鹽度(BS)和水深(D)5 個環境因子用CANOCO5.0 軟件進行運算。CCA 排序結果如圖8 所示。春季,CCA1 特征值為0.345,CCA2 特征值為0.230,CCA1 和CCA2 與環境因子相關系數分別為0.930 和0.900。蒙特卡洛顯著性檢驗結果顯示,兩排序軸呈現極顯著差異。兩個排序軸共解釋了主要蟹類組成21.36%的變異。表層水溫聚集在CCA2 的負軸,與CCA2 呈較大負相關;底層水溫聚集在CCA1 的負軸,與CCA1 呈較大負相關。其中,Z15 站位、Z16 站位和Z06 站位聚集在CCA1 的負軸,與底層水溫呈明顯負相關,說明水溫是影響其分布的最主要因素;Z08 站位聚集在CCA2 的負軸,與表層鹽度呈明顯的正相關,說明鹽度是影響該站位的主要因素。
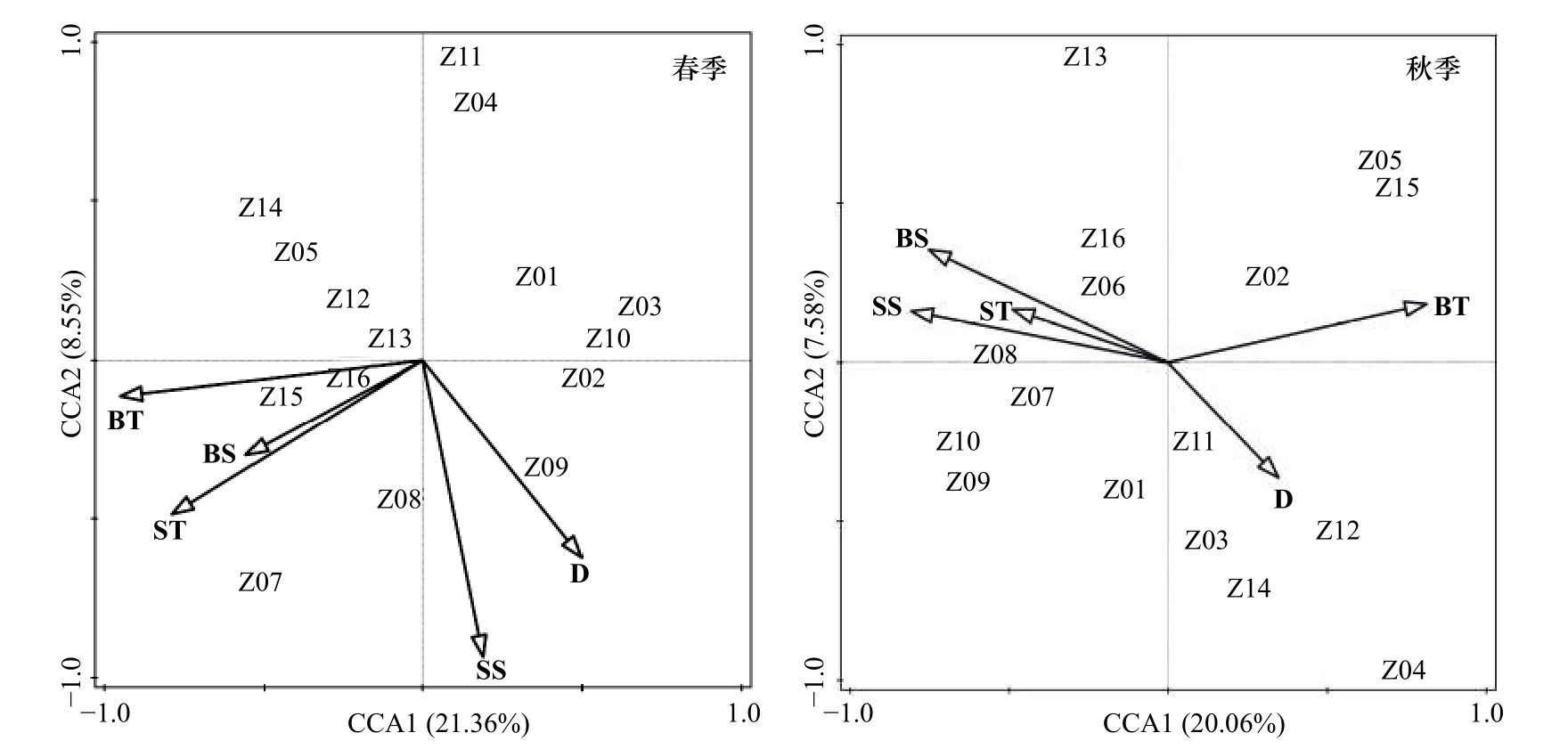
圖8 調查海域站位與環境因子典范對應分析排序Fig.8 Canonical correspondence analysis ranking of stations and environmental factors in the investigated sea area
秋季,CCA1 特征值為0.224,CCA2 特征值為0.136,CCA1 和CCA2 與環境因子相關系數分別為0.836 和0.818。蒙特卡洛顯著性檢驗結果顯示,兩排序軸接近于顯著差異。兩個排序軸共解釋了主要蟹類組成20.06%的變異。底層水溫聚集在CCA1 的正軸,與CCA1 呈較大正相關,表層鹽度、底層鹽度及表層水溫聚集在CCA1 的負軸,與CCA1 呈明顯負相關,其中,Z06 站位和Z08 站位聚集在CCA1 的負軸,與表層鹽度和底層鹽度呈明顯負相關,說明鹽度對其影響較大;春、秋兩季環境因子中水深對站位的影響效果較為一般。CCA 排序圖的分隔模式可能反映了主要蟹類對棲息地類型的偏好或者其生態習性的適應性。
4 討論
4.1 種類組成及優勢種分析
根據本次調查結果,在舟山群島東側海域春、秋季共有種和兩季特有種均為21 種,表明舟山群島東側海域種類豐富,地方性種類和季節性種類均占有一定比例,且蟹類區系組成均以廣溫、廣鹽性的暖水性種為主,這與俞存根等[1]開展的東海蟹類資源調查結果相符合,主要原因可能與調查海域受臺灣暖流和浙江沿岸水交互作用形成的混合水區有關。本次調查獲得的經濟蟹類主要有三疣梭子蟹、細點圓趾蟹、銹斑蟳、武士蟳、紅星梭子蟹(Portunus sanguinolentus)、日本蟳等,經濟蟹類重量占蟹類總重量的74.1%,說明該海域可以成為人們捕撈生產蟹類的漁場。本次調查結果中,兩季共同優勢種僅雙斑蟳1 種,一方面因為雙斑蟳在東海分布最為廣泛,其次雙斑蟳有著較長的繁殖期,春、夏、秋季都能捕到抱卵個體,其往往在夏季大量繁殖,秋季時出現漁獲高峰。而經濟價值較高的三疣梭子蟹僅為秋季的優勢種,其IRI 值相比春季大幅增加,主要原因是春季近岸海域水溫逐步回升,大量三疣梭子蟹從外海低溫深水區洄游到近岸淺水區和島礁區進行繁殖索餌,經過春、夏季的育肥階段,加之我國夏、秋季伏季休漁制度的保護,使得三疣梭子蟹在秋季大量出現,成為東海漁區重要的捕撈對象。本次調查中的蟹類資源優勢種數量與鄭獻之等[23]的研究結果相比有明顯減少,而與俞存根等[9]的調查結果對比,本調查中細點圓趾蟹成為東海北部附近海域的優勢蟹類。優勢種及經濟種類明顯減少,主要原因與過度捕撈和海洋環境污染有關,是否還有其他潛在影響因素,還有待進一步研究。
SIMPER 分析顯示,雙斑蟳、日本蟳和長手隆背蟹對春季相似性貢獻率較大,主要原因是雙斑蟳大多分布在黃海和東海海域,以東海分布最為廣泛;日本蟳在我國4 大海區均有分布,在東海主要分布在長江口漁場,是東海蟹類的第五大優勢種,主要汛期和產量高峰均在秋季;長手隆背蟹主要分布在東海和南海,數量以春、夏季最多,所以本次長手隆背蟹在春季貢獻率較高;春季為三疣梭子蟹的產卵繁殖季節,經過春、夏季的索餌育肥,在秋季達到產量高峰,所以三疣梭子蟹在秋季群落結構相似性中有較大貢獻。Cluster 聚類分析表明,調查海域秋季群落相似性大于春季(春季為34.1%,秋季為55.2%),也間接說明了秋季種類數和生物量水平高于春季。主要原因是秋季為三疣梭子蟹產量高峰期以及大規模的增殖放流提高了生物量,但是否還存在其他影響因素,仍有待進一步研究。
4.2 數量分布及生物多樣性變化
根據本次調查結果可知,春季調查海域蟹類數量為東南部外海占比較大,西部和西南部海域較少,秋季調查海域蟹類數量相比春季有一定西移和北移現象,在調查海域中部或南部近岸較高,東南部蟹類數量較低。其原因可能是每年冬季蟹類向南向東洄游至外海深水區進行越冬,翌年春季隨著水溫逐步上升,蟹類再次洄游到近岸島礁區和淺水區進行索餌產卵;夏、秋季臺灣暖流到達東海北部海域,并且夏、秋季大陸降水最多,長江沖淡水及杭州灣等陸地徑流達到最大,多個水團交互影響,加之夏季海域內浮游動植物豐度顯著上升,提高了海域內初級生產力,也吸引了大量蟹類到群島近岸海域進行索餌產卵活動。生物多樣性結果表明,春、秋季均勻度指數(J')平均值相差不大,而秋季豐富度指數(D)、多樣性指數(H')均大于春季,其主要原因可能是由于秋季臺灣暖流仍然影響東海北部海域,高溫、高鹽水吸引大量生物前來生殖產卵,也使廣溫、廣鹽性種類所占比例有所降低,使生物多樣性及豐富度顯著提高。以三疣梭子蟹為代表的秋季生物多樣性明顯大于春季,其原因一方面是春、夏季蟹類到近海海域產卵繁殖,加之夏季我國實施伏季休漁,減少了蟹籠和刺網等網具的作業頻率,另一方面浙江省早在2003 年就開始了三疣梭子蟹增殖放流活動,每年投放親體數量達千萬尾,從而提高了群體數量。
4.3 底層溫度和表層鹽度對蟹類群落的影響
典范對應分析結果表明,底層溫度和表層鹽度是影響該海域蟹類群落結構的主要環境因子。秋季平均溫度和平均鹽度均高于春季,究其原因可能為調查海域受臺灣暖流高溫、高鹽水和江浙沿岸水的共同影響,使該海域溫度和鹽度相對其他區域較高,侯偉芬等[24]在對舟山近岸漁場海域的水溫分布研究中也同樣證明了秋季水溫高于春季。調查海域遠離大陸,加之西北部及西部設立有嵊泗列島自然保護區和東極島自然風景區,水質環境較好,受人類活動干擾影響較小,因此吸引大多數蟹類在此棲息生存。鹽度方面,鹽度受陸地徑流的混合水臺灣暖流的高鹽水影響,以調查海域最北部Z01 站位和Z02 站位為代表,春季表層鹽度分別為31.71 和31.73,底層鹽度為33.47和33.61,秋季表層鹽度為33.37 和33.13,底層鹽度為33.61 和33.62。該規律同樣適用于調查海域最東部Z08 站位及Z09 站位,同時Z08 站位及Z09 站位秋季底層鹽度高達34.21 和34.26,明顯具備高鹽水特征,侯偉芬等[25]在對舟山近岸漁場海域鹽度的調查中也指出了由于受春季汛期和夏季臺風天氣降水較多影響,近岸海域鹽度低于遠海。水深方面,春、秋兩季中除Z04 站位和Z13 站位水深較淺(27 m、28 m)且漁獲較少,春季Z09 站位和Z10 站位水深最深且漁獲最多,其余站位平均水深均為60 m 上下,各站位水深相差不大,且水深層面與蟹類漁獲量分布無明顯規律,表現為無論水深多少,都有一定漁獲存在;同時通過典范對應分析也可發現,水深距離兩排序軸均較遠,相關性不明顯。該結果與丁朋朋等[26]對浙江南部近海蟹類以及鄭基等[27]對浙江披山海域蟹類研究的結果有所不同,原因可能是本調查海域不僅受臺灣暖流和浙江沿岸流影響,而且同時受到長江沖淡水及杭州灣入海水的影響,導致海域內鹽度有所下降,使鹽度對蟹類群落結構影響的相關性隨之降低。
4.4 蟹類生物對生態系統的作用
絕大部分蟹類屬雜食性和肉食性,少部分蟹類為植物食性,還有一部分蟹類攝取泥沙中的沉積物或水體中的懸浮物,在維持生態系統平衡和促進能量循環及物質流動中發揮著重要作用。蟹類不但可作為黃姑魚(Nibea albiflora)、褐菖鲉(Sebastiscus marmoratus)等高營養級魚類的餌料來源,還可捕食包括貝類、浮游生物和小型甲殼類在內的底棲生物,其中三疣梭子蟹和日本蟳還會捕食小型魚類、槍烏賊和多毛類等,食物范圍也較廣。此次調查海域島礁眾多,海域地處副熱帶季風氣候區,受大陸徑流和臺灣暖流等水系共同影響,浮游生物、底棲生物和游泳動物等種類繁多,是多種海洋生物產卵、繁殖、索餌和洄游的重要棲息地。蟹類生物作為水生生物食物網中重要的一環,既是生產者(魚類餌料生物)又是消費者(捕食底棲生物),對群落結構的穩定性起著至關重要的作用。在魚類資源嚴重衰退的今天,包括蟹類在內的甲殼類作為魚類資源的重要補充,其地位逐漸被人們所重視并成為人們重要的捕撈對象。與此同時,我們還應進一步加強對蟹類資源的保護,摸清蟹類生物繁殖與生長等生態學特性,掌握其種群變動規律,以期實現蟹類資源的可持續利用和生態環境的可持續發展。
致謝:顏文超、韓磊等同志參加海上樣品采集和室內種類鑒定,戴冬旭、張文俊、辛藝、孫蓓蓓、劉坤、于南京、張佩怡等同志參加室內種類鑒定和數據處理,謹致謝枕。
