海帶中抑藻活性化合物的分離及抑藻活性分析
2022-02-18 12:58:18孫穎穎朱文軒周靜莊麗雯毛奕淋趙秀芳王長海
海洋學報 2022年2期
關鍵詞:生長
孫穎穎 ,朱文軒 ,周靜,莊麗雯,毛奕淋,趙秀芳,王長海
(1.江蘇海洋大學 江蘇省海洋生物技術重點實驗室,江蘇 連云港 222005;2.江蘇海洋大學 江蘇省海洋生物產業技術協同創新中心,江蘇 連云港 222005;3.江蘇省海洋資源開發研究院,江蘇 連云港 222000;4.連云港市質量技術綜合檢驗檢測中心,江蘇 連云港 222005;5.南京農業大學 資源與環境科學學院,江蘇 南京 210095)
1 引言
赤潮已成為三大世界性近海污染問題之一,對近海水域生態環境和海洋養殖業造成了極大的威脅[1-2]。報道表明,在300 多種可形成赤潮的海洋浮游微藻中,超過70 種能產生毒素[3],藻毒素導致了魚類、貝類和其他海洋生物的大量死亡[1],也會引發人類疾病,甚至導致死亡。國內外研究者提出了許多預防和控制赤潮的方法(Prevent and Control HABs Methods,PCM),包括物理法(黏土[4])、化學法(表面活性劑[5])、生物法(病毒[6]、細菌[7]和殺藻[8]或抑藻化合物[3,9])等。
近30 年來,分離自大型海藻的一些抑藻活性化合物,被認為是開發環境友好型赤潮微藻抑藻劑的理想材料之一[10-11]。例如,小珊瑚藻(Corallina pilulifera)中的溴仿[12]、江蘺(Gracilaria lemaneiformis)中的8-hydroxy-4E 和6E-octadien-3-on[13]、網地藻(Dictyota dichotoma)中的Dictyolactone[14]、葉狀鐵釘菜(Ishige sinicola)中的α-單甘酯[15]以及條斑紫菜(Porphyra yezoensis)中的gossonorol[16]等。在前期工作中,我們整理了1960-2019 年被Web of Science、Springer、Google Scholar 和CNKI 等數據庫收錄的大型海藻抑藻研究,發現120 多種可抑制赤潮微藻生長的大型海藻中,大型褐藻為40 種[10],它們依次歸屬于網地藻目(Dictyotales)、水云目(Ectocarpales)、墨角藻目(Fucales)、鐵釘菜目(Ishigeales)、海帶目(Laminariales)和線翼藻目(Tilopteridales)(圖1)。其中,海帶目的大型褐藻占20%,例如,腔昆布(Ecklonia cava)[17]、鵝掌菜(Ecklonia kurome)[18]、Eckloniopsis radicosa[15]、籠目海帶或日本厚葉海帶(Kjellmaniella crassifolia)[19]、海帶(Laminaria japonica)[19-22]、掌狀昆布(Laminaria digitata)[20]、皺海帶(Laminaria religiosa)[23]、裙帶菜(Undaria pinnatifida)[21]。
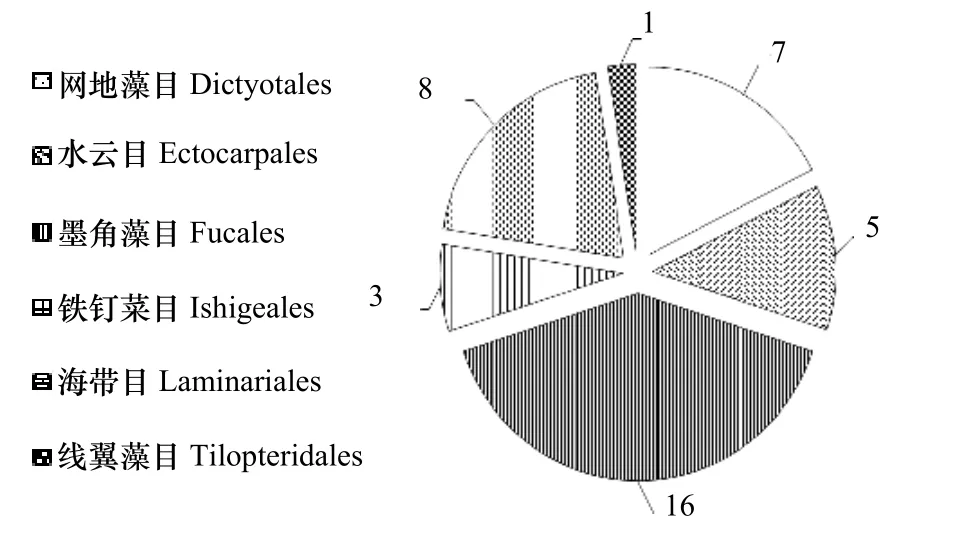
圖1 可抑制赤潮微藻的大型褐藻的目類歸屬分布Fig.1 Distribution of brown marine macroalgae with antialgal activity against red tide microalgae according the orders
不同研究者已經發現海帶(L.japonica)對某些海洋微藻具有一定的抑藻活性[19-22],李鳳超[19]對海帶中防污物質進行了初步分離和鑒定。然而,目前為止,并未見抑藻活性化合物從海帶中分離獲得。基于此,本文采用活性追蹤分離方法,以米氏凱倫藻為抑藻檢測對象,從海帶中分離抑藻活性化合物。進一步,分析它們對塔瑪亞歷山大藻(Alexandriumtamarense)、強壯前溝藻(Amphidinium carterae)、赤潮異彎藻(Heterosigma akashiwo)、米氏凱倫藻(Karenia mikimotoi)、球形棕囊藻(Phaeocystis globsa)和中肋骨條藻(Skeletonema costatum)生長的抑制作用,獲得每種化合物對赤潮微藻生長的半抑制效應濃度(EC50~96h),為環境友好型赤潮微藻抑藻劑的研發奠定實驗基礎和提供技術支撐。
2 材料與方法
2.1 實驗材料
塔瑪亞歷山大藻、強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻無菌株由上海光語生物科技有限公司提供,在f/2 培養基中培養,培養溫度為(22±2)℃,光照強度為40 μmol/(m2·s),光暗比為12 h∶12 h。
天然海水靜置1 個月以上,經脫脂棉和300 目篩絹過濾,高溫滅菌、冷卻,使用前將pH 和鹽度分別調節至8.5 和30 備用。
海帶半干品購置于江蘇碧藍海洋生物科技有限公司,自來水清洗后,室內自然晾曬2 d。50℃下烘干后,粉碎至0.3 mm 粒徑備用。
2.2 抑藻活性化合物的分離純化
2.2.1 提取
將海帶干粉末2 000 g 和6 L 甲醇加入到動態熱回流提取濃縮機(TX-NX-10)中,在30℃下浸提6 h。上述浸提過程重復3 次,合并浸提液。浸提液經過濾和減壓蒸干,得到黑色黏稠狀浸膏68.22 g。向此浸膏中加入90%甲醇水溶液300 mL,充分振蕩,4℃下靜置過夜,4℃下離心10 min、過濾,去除沉淀,獲得上清液280 mL。60℃下減壓蒸干后,溶解于蒸餾水中,配制濃度為80 g/L 溶液(提取物溶液)進行抑藻活性檢測和液液萃取分離。抑藻活性檢測時,提取物濃度依次設定為1.0 mg/mL、2.0 mg/mL、4.0 mg/mL、8.0 mg/mL和16.0 mg/mL。
2.2.2 液液萃取分離
將乙酸乙酯加入到上述提取物溶液中,萃取3 次,乙酸乙酯加入體積依次為60 mL、20 mL 和20 mL。乙酸乙酯層合并收集后,然后在40℃下減壓蒸干,獲得2.58 g 組分A。下層40℃下減壓蒸發除去乙酸乙酯后,加入正丁醇萃取3 次,正丁醇加入體積依次為60 mL、20 mL 和20 mL。上層和下層均在60℃下減壓蒸干,分別獲得2.23 g 組分B 和11.34 g 組分C。上述3 種液液萃取分離組分,分別溶解于無水甲醇中,制備濃度為20 g/L 的溶液進行抑藻活性檢測(液液萃取分離組分濃度設定為2 mg/mL)和硅膠柱層析分離。
2.2.3 硅膠柱層析分離
組分B 加載于硅膠柱(200~300 目,3.0 cm×40 cm)上,以氯仿和甲醇(體積比為1∶5,以下均為體積比)為洗脫液,流速為1.0 mL/min,每管接收餾分10 mL。組分C 采用硅膠層析柱(200~300 目,5.0 cm×80 cm)進行分離,氯仿和甲醇(1∶5)為洗脫液,流速為1.0 mL/min,每管接收餾分20 mL。洗脫2 倍柱體積后,所得餾分40℃下減壓濃縮,進行硅膠GF254薄層層析檢測,展開劑為氯仿和甲醇(1∶10)。合并相同餾分后,40℃下減壓蒸干除去溶劑,獲得再分組分B1和B2、再分組分C1、C2和C3。上述5 個組分分別稱取10 mg,依次溶解于1 mL 甲醇溶液中,配制濃度為10 g/L 的溶液。進行抑藻活性檢測時,再分組分濃度設定為1.0 mg/mL。結果表明,3 個再分組分(B1、B2和C2)對米氏凱倫藻具有較為明顯的抑藻活性。
2.2.4 Sephadax LH-20 凝膠柱層析分離
將上述3 個再分組分分別加載于Sephadax LH-20 凝膠層析柱上(2.0 cm×25 cm),均以甲醇為洗脫液。洗脫2 倍柱體積后,所得餾分40℃下減壓濃縮,經硅膠GF254薄層層析檢測,合并、減壓蒸干后,依次獲得凝膠分離組分B11、B12、B13、B21(0.56 g)、C21和C22。將它們分別稱取5 mg,溶解于0.5 mL 甲醇中,進行抑藻活性檢測,濃度設定為0.25 mg/mL。結果表明,凝膠分離組分B13、B21和C21顯著地抑制了米氏凱倫藻的生長(其中,B21已經完成分離純化)。
2.2.5 薄層層析純化制備
采用劃線法將凝膠分離組分B13和C21加載于硅膠薄層層析預制板(100 mm×200 mm)上,分別以石油醚和乙酸乙酯(1∶2)及氯仿和甲醇(4∶1)為展開劑。經展開,凝膠分離組分B13和C21分別在硅膠預制板上呈現2 個清晰且相距較明顯的條帶。將相應條帶分別刮下收集,用丙酮浸泡,過濾和減壓蒸干后,制備得到樣品B131(0.24 g)、B132(0.19 g)、C211(0.008 g)和C212(0.011 g)。適量稱取5 個樣品,分別溶解于甲醇溶液后,點樣于硅膠GF254上,依次以氯仿和甲醇(1∶1)、環己烷和乙酸乙酯(1∶2)及正丁醇和醋酸和水(1∶1∶0.5)為展開劑進行展開,發現此5 個樣品均呈現單一斑點;同時,采用通用型顯色劑(10%硫酸溶液和碘)檢測,發現它們同樣呈現單一斑點,表明純度達到了薄層純。抑藻活性檢測時,樣品濃度設定為0.25 mg/mL。
2.3 結構鑒定
采用高分辨電噴霧質譜(HR-ESI-MS)和核磁共振譜(1H-NMR 和13C-NMR)鑒定樣品結構。LTQObitrap XL 光譜儀測定HR-ESI-MS,Bruker AV Ⅲ 600質譜儀測定NMR(四甲基硅烷(TMS)為標準品)。
2.4 抑藻活性
2.4.1 分離純化過程中抑藻活性檢測
在分離純化過程中,以米氏凱倫藻為抑藻活性測試微藻,接種細胞數量為(10~12)×104cells/mL。將待測組分溶解于甲醇后,移取1 μL 到微藻培養體系(米氏凱倫藻1 mL+f/2 培養基9 mL),終濃度按照以上相應分離過程進行設定。對照組為10 mL 培養體系中加入1 μL 甲醇,每個實驗設定3 個平行樣。所有培養瓶置于光照培養箱中進行培養,溫度為(22±2)℃,光照強度為62 μmol/(m2·s),光暗比為12 h∶12 h,培養10 d。
2.4.2 抑藻化合物對赤潮微藻的抑藻活性分析
將化合物(市售,分析純)溶解于甲醇溶液,配制濃度為250mg/mL,再依次稀釋為50mg/mL、10mg/mL和2mg/mL。分別量取1μL上述溶液,加入到4999μL培養體系中(赤潮微藻500μL+f/2培養基4499μL),化合物終濃度依次為50μg/mL、10μg/mL、2μg/mL和0.4μg/mL。塔瑪亞歷山大藻、強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻的起始細胞數量分別為(8~10)×104cells/mL、(20~24)×104cells/mL、(8~11)×104cells/mL、(10~14)×104cells/mL、(45~49)×104cells/mL 和(26~29)×104cells/mL。對照組為4 999 μL 培養體系中加入1 μL 甲醇溶液,每個實驗設定3 個平行樣。同時,設定相同濃度的硫酸銅作為陽性對照組。所有培養瓶置于光照培養箱中進行培養,培養4 d,其他培養條件同2.4.1 節。同時,顯微鏡下(×400)觀察藻細胞形態。
2.5 實驗指標測定
每日取樣,計數赤潮微藻細胞數量,計算抑藻劑對赤潮微藻的生長抑制率(I),計算公式為
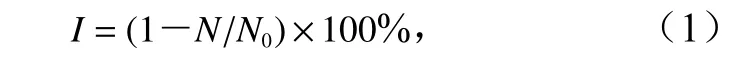
式中,N為處理組藻細胞數量;N0為對照組藻細胞數量。
采用文獻[24]的方法計算硫酸銅和化合物對赤潮微藻生長的EC50~96h。
2.6 數據處理
實驗數據采用SPSS 11.5 軟件包進行獨立樣本檢驗統計分析,p<0.05 為顯著性差異,p<0.01 為極顯著性差異。
3 結果和分析
3.1 海帶中抑藻活性化合物活性追蹤的分離純化
3.1.1 海帶提取物對米氏凱倫藻生長的影響
從圖2 可以看出,隨濃度增加,海帶提取物對米氏凱倫藻生長的抑制作用明顯增強(p<0.05)。當濃度為8.0 mg/mL 時,提取物對米氏凱倫藻的生長抑制率為56.2%(第10 天)。結果表明,海帶提取物具有明顯抑制米氏凱倫藻的抑藻活性,需要進行后續分離。
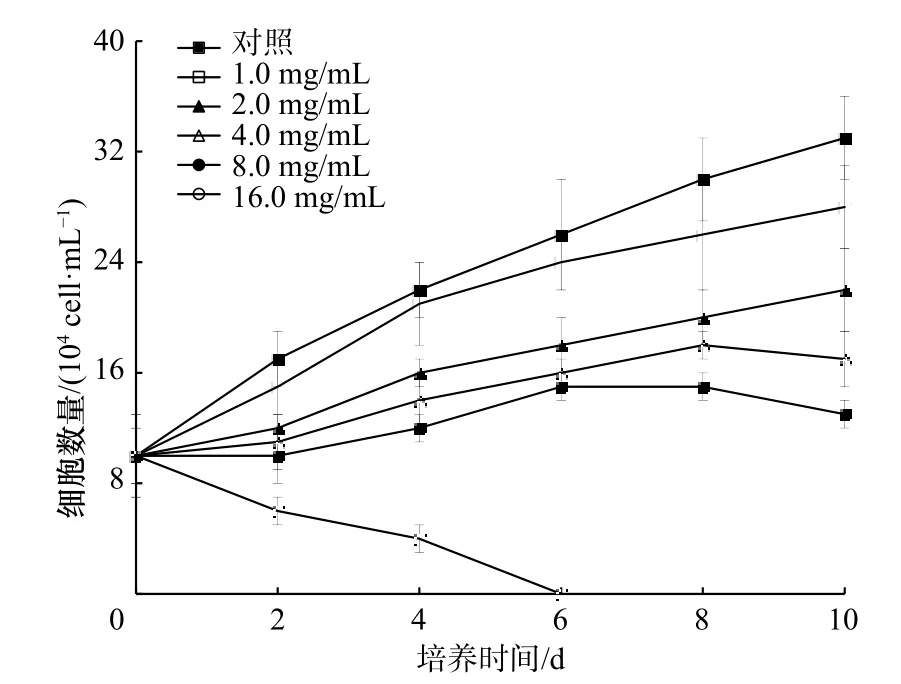
圖2 海帶甲醇提取物對米氏凱倫藻生長的影響Fig.2 Effect of the methanol extracts of Laminaria japonica on the growth of Karenia mikimotoi
3.1.2 液液萃取分離組分對米氏凱倫藻生長的影響
在圖3 中,組分B 和組分C 明顯抑制了米氏凱倫藻的生長(p<0.05)。第10 天時對米氏凱倫藻的生長抑制率超過76%。因此,將組分B 和組分C 進行硅膠柱層析分離。
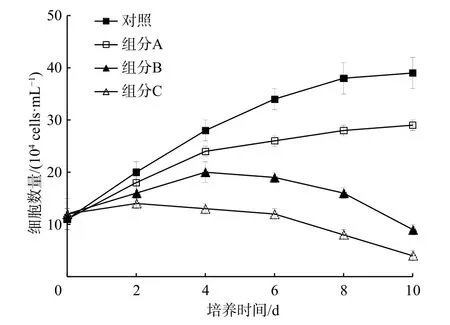
圖3 液液萃取分離組分對米氏凱倫藻生長的影響Fig.3 Effect of several liquid-liquid extraction components from the methanol extracts on the growth of Karenia mikimotoi
3.1.3 硅膠柱層析和Sephadax LH-20 凝膠柱層析分離組分以及薄層層析純化樣品對米氏凱倫藻生長的影響
由表1 可知,組分B 和組分C 經硅膠柱層析分離后,得到5 個再分組分。抑藻活性檢測表明,再分組分B1、B2和C2對米氏凱倫藻具有明顯的抑藻活性(p<0.05)。
將上述3 個再分組分B1、B2和C2分別進行Sephadax LH-20 凝膠柱層析分離,組分B1分離為B11、B12和B13;組分B2分離為B21;組分C2分離為C21和C22(表1)。在6 個組分中,B13、B21和C21均對米氏凱倫藻生長表現出明顯的抑制作用(p<0.05),其中B21薄層層析檢測顯示僅有1 個斑點。隨后,將B13和C21進行硅膠薄層層析制備,
經薄層層析純化制備,得到4 個樣品,B131、B132、C211和C212(表1)。濃度為0.05 mg/mL 時,它們對米氏凱倫藻的生長表現出明顯的抑藻活性(p<0.05)。
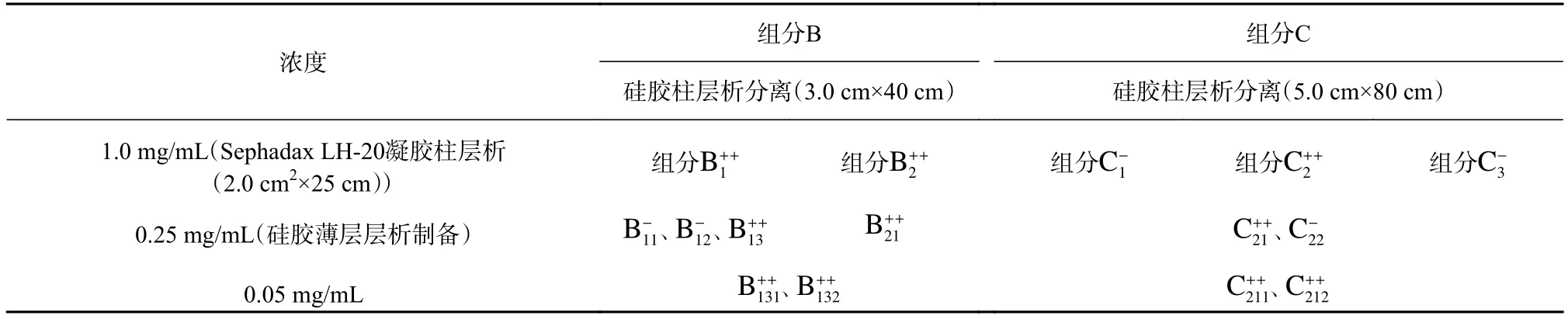
表1 組分B 和組分C 的分離純化過程中抑藻活性檢測Table 1 Antialgal activity test of the fractions B and C in the isolation and purification process
綜上,采用一系列分離方法,從海帶中純化到對米氏凱倫藻的生長具有抑制作用的活性化合物,分離純化具體過程見圖4。
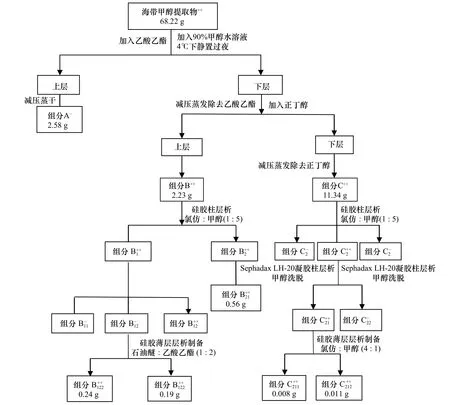
圖4 海帶中抑藻活性化合物的分離純化Fig.4 Isolation and purification of the antialgal compounds from Laminaria japonica
3.2 結構鑒定
樣品B131,無色粉末。1H-NMR (600 MHz,DMSOd6)δH:3.51 (2H,t,J=6.0 Hz,H-7);13C NMR (150 MHz,DMSO-d6)δC:177.2 (C-11)、41.8 (C-2)、23.4 (C-3)、30.4(C-4)、30.2 (C-5)、36.8 (C-6)、63.5 (C-7)。以上數據與文獻[25]基本一致,故鑒定樣品B131為7-羥基庚酸。
樣品B132,無色粉末。1H-NMR (600 MHz,DMSOd6)δH:0.88 (3H,t,J=6.6 Hz,H-14)、2.35 (2H,m,H-2);13C-NMR (150 MHz,DMSO-d6)δC:177.9 (C-1)、33.8 (C-2)、24.9 (C-3)、29.2 (C-4)、29.4 (C-5)、29.9 (C-6)、29.7(C-7)、29.6 (C-8)、31.4 (C-9)、29.5 (C-10/11)、32.1 (C-12)、22.8 (C-13)、14.3 (C-14)。這些數據與十四酸的光譜數據基本一致[26],因此,將樣品B132鑒定為十四酸。
樣品B21,白色粉末。1H-NMR (600 MHz,DMSOd6)δH:2.32 (2H,t,J=7.8 Hz,H-2)、1.63 (2H,m,H-3)、0.87 (3H,t,J=7.2 Hz,H-16);13C-NMR (150 MHz,DMSO-d6)δC:179.3 (C-1)、34.2 (C-2)、25.1 (C-3)、29.5(C-4)、29.6 (C-5)、29.8~30.1 (C-6~12)、29.8 (C-13)、32.3 (C-14)、23.1 (C-15)、14.5 (C-16)。樣品B21的核磁共振譜的數據與文獻[27]基本一致,因此,鑒定其為十六酸。
樣品C211(白色蠟狀固體),1H-NMR (600 MHz,DMSO-d6)δH:2.31 (2H,t,J=7.8 Hz,H-2)、1.63 (2H,m,H-3)、1.28 (28H,m,H-4~H-17)、0.88 (3H,t,J=7.2 Hz,H-18);13C-NMR (150 MHz,DMSO-d6)δC:177.1 (C-1)、34.0 (C-2)、31.9 (C-16)、29.1~29.7 (C-4~C-15)、24.7(C-3)、22.7 (C-17)、14.5 (C-18)。這些數據與文獻[28]一致,故將樣品C211鑒定為十八酸。
樣品C212,無色油狀物。1HNMR (600 MHz,MeOD)δH:0.86 (3H,t,J=6.6 Hz,H-18)、1.23 (8H,m,m,H-4~H-7),1.24 (20H,m,H-12~H-17)、1.61 (2H,m,H-3)、1.94(4H,m,H-8,11)、2.33 (2H,t,J=7.2 Hz,H-2)、5.28 (2H,dd,J=8.4,5.4 Hz,H-9,H-10)。以上數據與油酸的核磁共振譜數據基本一致[29]。
3.3 5 種化合物對6 種赤潮微藻的抑藻活性分析
在圖5 中,5 種化合物對6 種赤潮微藻生長表現出明顯的選擇性抑制作用,其中7-羥基庚酸表現出最為強烈的抑制作用(p<0.05),且這種抑制作用隨著濃度的增加而明顯增強(p<0.05)。第4 天,在濃度為50 μg/mL 時,7-羥基庚酸對塔瑪亞歷山大藻、強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻的生長抑制率分別為53.3%、84.3%、71.2%、72.3%、74.4%和87.5%;十四酸明顯地抑制了強壯前溝藻、米氏凱倫藻和中肋骨條藻的生長(p<0.05),在濃度50 μg/mL 時,對它們的生長抑制率超過52%(第4 天);十六酸僅明顯抑制了強壯前溝藻的生長(p<0.05),對強壯前溝藻的生長抑制率為52.5%(50 μg/mL,第4 天),對其他5 種赤潮微藻的生長抑制率低于40%;十八酸和油酸對6 種赤潮微藻沒有明顯的生長抑制作用(p>0.05),在濃度為50 μg/mL 時,對它們的生長抑制率低于40%。
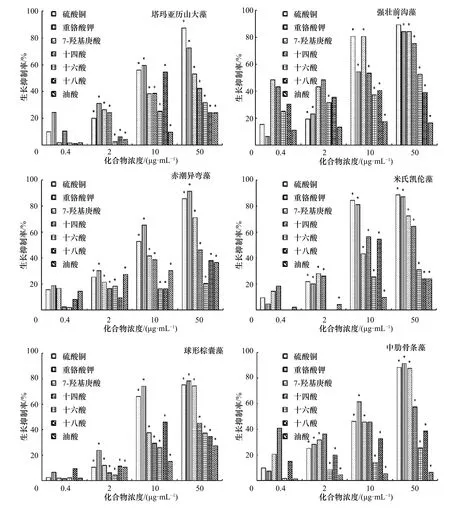
圖5 不同化合物對6 種赤潮微藻生長的抑制作用Fig.5 Growth inhibition of the different compounds against six species of red tide microalgae
進一步,計算得出了7-羥基庚酸等5 種脂肪酸對6 種赤潮微藻生長的半抑制效應濃度(EC50~96h)(表2)。硫酸銅和重鉻酸鉀是藻類毒性實驗中常用的陽性對照組,硫酸銅對6 種赤潮微藻生長的EC50~96h在表2列出,由前期研究可知,重鉻酸鉀對強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻生長的EC50~96h依次為3.9 μg/mL、36 μg/mL、16 μg/mL、38 μg/mL 和2.7 μg/mL[9,30]。除十四酸、十六酸和十八酸對塔瑪亞歷山大藻生長的EC50~96h已被報道[31],7-羥基庚酸等5 種脂肪酸和硫酸銅對赤潮微藻生長的EC50~96h是第一次被明確。在本文中,十四酸和十六酸對塔瑪亞歷山大藻生長的EC50~96h與已有報道非常接近[31],但十八酸對塔瑪亞歷山大藻生長的EC50~96h與已有報道相差很大[31],具體原因目前還在分析中。
國際經合組織(OECD)指出,當EC50~96h分別為小于1 μg/mL、1~10 μg/mL、10~100 μg/mL 和大于100 μg/mL時,化合物對藻類具有極高毒性、高毒性、中等毒性和極低毒性[32]。根據此標準,7-羥基庚酸、十四酸和十六酸對6 種赤潮微藻具有中等或高等毒性,其中7-羥基庚酸對塔瑪亞歷山大藻和強壯前溝藻、十四酸對強壯前溝藻均為高毒性。與重鉻酸鉀對赤潮微藻生長的EC50~96h比較,7-羥基庚酸和十四酸對強壯前溝藻、赤潮異彎藻、米氏凱倫藻和球形棕囊藻生長的EC50~96h更小,十六酸對米氏凱倫藻生長的EC50~96h更小,這表明7-羥基庚酸和十四酸在抑制強壯前溝藻等4 種赤潮微藻、十六酸在抑制米氏凱倫藻上比重鉻酸鉀更具有抑藻優勢。同時,從表2 可以看出,7-羥基庚酸和十四酸對強壯前溝藻的EC50~96h明顯小于硫酸銅對該赤潮微藻的EC50~96h。綜合來看,7-羥基庚酸和十四酸在抑制強壯前溝藻上具有明顯優于重鉻酸鉀和硫酸銅的抑藻優勢。
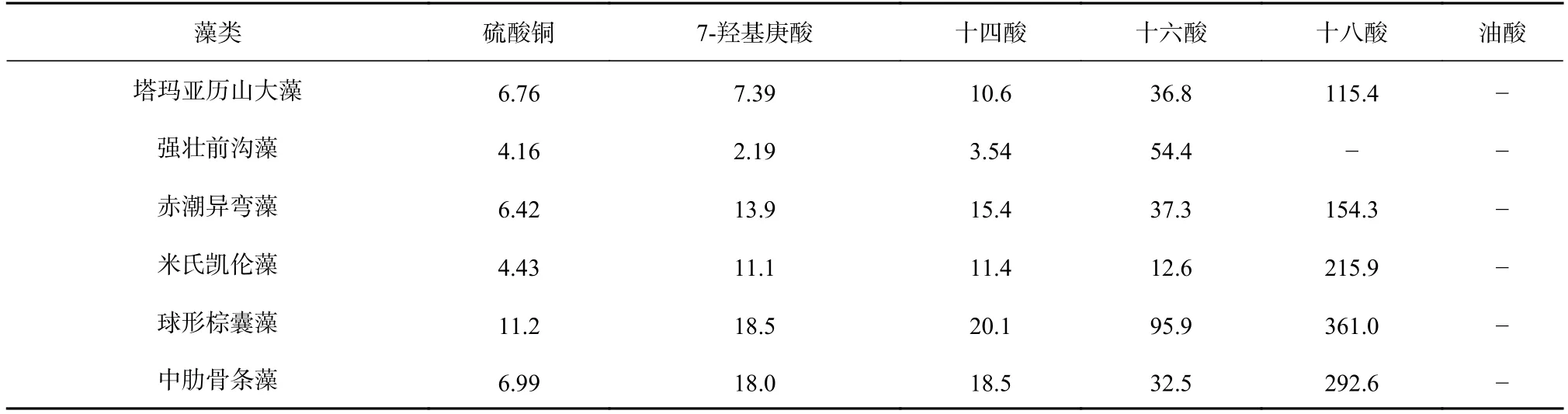
表2 不同化合物對6 種赤潮微藻生長的半抑制效應濃度(EC50~96h,單位:μg/mL)Table 2 Different compounds isolated from Laminaria japonica for the six species of red tide microalgae (EC50-96h,unit:μg/mL)
同時,我們發現在脂肪酸作用下,藻細胞形態發生了某些明顯變化。在7-羥基庚酸作用下,強壯前溝藻、赤潮異彎藻、米氏凱倫藻和球形棕囊藻細胞形態改變見圖6。在圖6 中,能清晰看到強壯前溝藻和赤潮異彎藻細胞膜模糊、類似溶解,橫鞭毛脫落,形狀改變明顯;米氏凱倫藻細胞破碎;球形棕囊藻細胞變長、伴有色素體缺陷等現象。在十四酸和十六酸能明顯抑制的赤潮微藻中,藻細胞形態也發現了這些類似的變化。
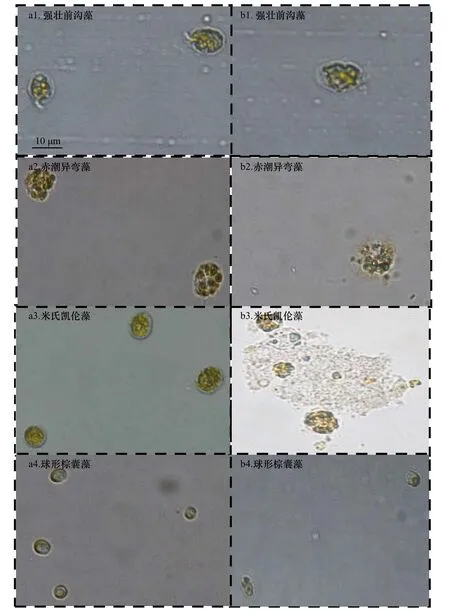
圖6 7-羥基庚酸對4 種赤潮微藻細胞形態的影響Fig.6 Effect of 7-hydroxyheptanic acid on the photographs of the four species of red tide microalgae
4 討論
大型海藻是抑制赤潮微藻活性化合物的良好來源之一[32-35],某些抑藻活性化合物結構已經被鑒定,例如,α-亞麻酸[23,36]、dictyolactone[14]、α-單甘酯[15]、二十二碳酸甲酯[12]等。這些研究表明,大型海藻中抑藻活性化合物結構多樣,包括脂肪酸、萜類和其他化合物[12,14-15,23,37-38]。目前,已經從葡萄藻(Botryococcus braunii)[14]、褐藻(Cladosiphon okamuranus)[39]、小珊瑚藻(Corallina pilulifera)[39]、江 蘺(Gracilaria lemaneiformis)[13]、裂片石莼(Ulva fasciata)[23]、孔石莼(Ulva pertusa)[9]和滸苔(Ulva prolifera)[40-41]等大型海藻中分離到脂肪酸類抑藻活性物質(圖1)。在本文,采用抑藻活性追蹤分離方法(圖2 至圖4,表1),第一次從海帶中分離到7-羥基庚酸、十四酸、十六酸、十八酸和油酸(純度為薄層純),其中7-羥基庚酸、十四酸和十八酸是第一次從大型海藻中分離獲得。十六酸已從葡萄藻[34]、江蘺[13]、腸滸苔(Ulva intestinalis)[41]和滸苔[40]中分離得到;在龍須菜(Gracilaria lemaneiformis)分離到油酸[42]。
目前,脂肪酸對淡水微藻生長的抑制作用[43-47]和對赤潮微藻生長的抑制作用[12,23,31,34-35,39-41,48-49]已有一些研究報道。在本文中,除塔瑪亞歷山大藻和赤潮異彎藻外,7-羥基庚酸、十四酸、十六酸、十八酸和油酸對強壯前溝藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻生長的抑制作用是第一次被研究。在脂肪酸對淡水微藻—分散藻羊角月牙藻(Selenastrum capricornutum)急性毒性研究中,研究者認為,脂肪酸的抑藻作用與脂肪酸的碳鏈長度有關[47]。尹玉麗[31]指出,飽和脂肪酸對塔瑪亞歷山大藻生長抑制強弱的順序依次為十四酸、十六酸、十八酸。何宗祥和張庭廷[44]及姜聞新等[45]也認為,脂肪酸的抑藻作用與脂肪酸碳鏈長度有關。在本研究中,我們發現7-羥基庚酸和十四酸的抑藻活性要明顯強于其他3 種脂肪酸的抑藻活性,十六酸、十八酸和油酸僅具有微弱的抑藻活性。然而,十六酸、十八酸和油酸對試驗赤潮微藻的抑藻活性并未有明顯的強弱差異(圖5)。因此,本文研究結果僅在一定程度上符合“脂肪酸碳鏈越短,抑制作用越強”的說法。同時發現,在濃度為50 μg/mL 時,7-羥基庚酸對強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻的生長抑制率與硫酸銅和重鉻酸鉀對此5 種赤潮微藻的生長抑制率接近(第4 天,圖3)。
在脂肪酸對微藻生長的抑制作用機理研究中,某些研究者認為,脂肪酸主要通過細胞膜上離子通道的途徑產生抑制作用[32,50-51],具體為脂肪酸引起離子通道結構變化,導致細胞膜通透性發生改變,某些重要離子,譬如Ca2+流出,致使藻細胞內與這些離子相關的生理活動受到影響,從而引發藻細胞死亡。進一步研究指出,由于藻細胞質膜中十六酸、亞油酸等脂肪酸含量較高,外加這些脂肪酸時,它們更易與質膜結合導致其構像發生變化,致使藻細胞離子通道結構發生變化[52]。因此,某些研究指出,脂肪酸對微藻生長的抑制作用與試驗微藻細胞的脂肪酸組成有關系[44-45,49]。在本研究中,我們發現不同脂肪酸作用下藻細胞出現細胞膜模糊、細胞破碎或色素體缺陷等現象(僅列出7-羥基庚酸作用下藻細胞形態的某些改變,圖6),很明顯藻細胞受到了脂肪酸不同程度的影響。6 種試驗赤潮微藻中,除強壯前溝藻脂肪酸組成未見報道外,其他5 種赤潮微藻中,十六酸含量最高,多數含有十四酸和十八酸,僅塔瑪亞歷山大藻和赤潮異彎藻含有油酸[53-57]。5 種脂肪酸對6 種赤潮微藻的抑藻強弱順序為7-羥基庚酸、十四酸、十六酸、十八酸、油酸,根據本文現有結果尚無法確定本文試驗的赤潮微藻脂肪酸組成與脂肪酸對它們的生長抑制作用之間的關系,還需要在后續工作中進行研究。
表2 表明,7-羥基庚酸、十四酸和十六酸對6 種試驗赤潮微藻具有中等或高等毒性,其中7-羥基庚酸和十四酸對試驗的赤潮微藻具有更為強烈的毒性。與硫酸銅和重鉻酸鉀對試驗赤潮微藻生長的EC50~96h相比,7-羥基庚酸和十四酸對強壯前溝藻生長的EC50~96h影響更小。這就表明,在抑制強壯前溝藻上,7-羥基庚酸和十四酸更具有抑藻優勢。同時,與以往研究相比[9-10,16],7-羥基庚酸對強壯前溝藻生長的EC50~96h不僅小于孔石莼、條斑紫菜和江蘺中分離到的抑藻活性化合物對該種赤潮微藻生長的EC50~96h,而且也小于硫酸銅和重鉻酸鉀對強壯前溝藻生長的EC50~96h。硫酸銅和重鉻酸鉀能迅速殺死赤潮微藻,然而,它們對非赤潮微藻的殺滅或抑制作用,會影響整個生態系統。7-羥基庚酸作為海帶生長過程中產生的代謝物,在自然條件下能降解,不會長期積累,生態安全性好,可替代硫酸銅和重鉻酸鉀,有望成為一種新型的環境友好型赤潮微藻抑藻劑。
我們的一個前期研究總結了近20 年大型海藻抑藻活性化合物[10],發現分離自江蘺的gossonorol 和7,10-epoxy-ar-bisabol-11-ol[31]對強壯前溝藻生長的EC50~96h與本研究中7-羥基庚酸和十四酸對該赤潮微藻生長的EC50~96h較為接近,gossonorol 和7,10-epoxyar-bisabol-11-ol 是倍半萜類化合物、7-羥基庚酸和十四酸是脂肪酸,從結構上來看,羥基很可能是產生抑制作用的基團。此4 種抑藻活性化合物的分離純化過程有一定的相似性,均是采用溶劑浸提、液液萃取、硅膠柱層析或薄層層析分離等方法,但具體分離過程并不相同。讓人遺憾的是,先前工作和本研究都沒有進行抑藻機理分析,它們在對赤潮微藻生長的抑制效應方面是否存在相似性尚不得知,需要后續進行深入研究。
近50 年來,利用大型海藻與赤潮微藻間抑制作用來生物防控赤潮微藻的研究有了長足進步,取得了一系列研究成果。然而,大型海藻抑藻活性化合物分離純化研究仍然非常欠缺,致使利用大型海藻抑藻活性化合物開發環境友好型赤潮微藻抑藻劑受到了極大限制,因此,亟需海洋科學領域的研究者開展相關研究來解決這個問題。
5 結論
(1)采用活性追蹤的分離方法,第一次從海帶中分離純化得到7-羥基庚酸、十四酸、十六酸、十八酸和油酸,純化為薄層純。
(2)7-羥基庚酸具有較為廣泛的抑藻活性,在濃度為50 μg/mL 時,第4 天時對塔瑪亞歷山大藻、強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻的生長抑制率超過53%。十四酸(50 μg/mL)也對強壯前溝藻、米氏凱倫藻和中肋骨條藻的生長表現出明顯的抑制作用,對它們的生長抑制率超過57%(第4 天)。
(3)明確了7-羥基庚酸、十四酸、十六酸、十八酸和油酸對塔瑪亞歷山大藻、強壯前溝藻、赤潮異彎藻、米氏凱倫藻、球形棕囊藻和中肋骨條藻生長的半抑制效應濃度EC50~96h。發現7-羥基庚酸和十四酸在抑制強壯前溝藻上比硫酸銅和重鉻酸鉀更具有抑藻優勢。
致謝:感謝中國醫學科學院藥用植物研究所馬國需博士對樣品結構解析的幫助。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
