油菜苗期耐鹽堿性綜合評價與根際土壤有機酸含量比較
2022-02-18 07:23:56吳鵬博李立軍張艷麗
作物雜志 2022年1期
吳鵬博 李立軍 張艷麗
(內蒙古農業大學農學院,010018,內蒙古呼和浩特)
土壤鹽堿化制約農業生產的發展,影響全球各大洲干旱地區100多個國家的9.55億hm2農田,其中約有 37%的土地為堿性鹽土[1-3]。世界三大鹽堿土聚集區之一的松嫩平原是我國面積最大的堿性鹽堿地,其面積有繼續擴大的趨勢[4-5]。土壤鹽堿化類型為 NaHCO3或 Na2CO3等堿性鹽比 NaCl或Na2SO4等中性鹽對植物的傷害更大[6-8],目前農業生態學家對中性鹽的脅迫機制和原因給予了較多關注,而對堿性鹽脅迫的關注較少。
油菜(Brassica napus L.)是我國發展鹽堿農業的理想候選作物[9]。雙低油菜對鹽堿的耐受等級為中抗及以上[10],油菜耐鹽堿能力改良的起點較高,但不同油菜種質抗鹽堿能力存在差異。在大豆[11]、棉花[12]和燕麥[13]等作物已有關于植物耐鹽堿性鑒定及指標篩選的報道,而關于油菜耐鹽堿性的研究較少。苗期被認為是大多數作物生長受鹽堿脅迫影響最關鍵的時期[14]。因此,通過對不同油菜種質進行苗期堿性鹽脅迫,綜合評價其耐鹽堿性,篩選耐鹽堿性較強的油菜種質,對油菜耐鹽堿品種的選育具有重要意義。
植物有機酸代謝調節在抵抗逆境脅迫時起關鍵作用。例如,植物可以通過根系分泌的有機酸對土壤進行金屬解毒[15-17],并可清除土壤中放射性污染物鈾[18]。有機酸的積累是植物對高pH值環境的一種適應性表現,例如堿脅迫下沙棘[8]和星星草[19]等植物體內均可積累大量的有機酸,只是積累的有機酸種類不盡相同,其中主要包括草酸、檸檬酸、蘋果酸和琥珀酸等。有研究[20]表明,有機酸可作為pH調節劑積累于細胞中,以補充土壤在高pH環境下根系分泌 H+及其他代謝產物對根外微環境調節的不足。也有研究發現,有些堿敏感植物[21]和低抗堿鹽性植物[22]不會積累有機酸。根際土壤因化學和生物學性質最活躍而受到越來越多的重視[23]。本研究對堿性鹽脅迫下油菜根際土壤有機酸含量的變化進行分析,探討有機酸在油菜抗鹽堿過程中的生理作用,為解讀油菜對堿性鹽脅迫的適應機制提供理論依據。
1 材料與方法
1.1 供試材料與土壤
8個供試油菜種質由華中農業大學提供,分別是華油雜62、青雜9號、崇2、17崇1、16-P32外、16-P22外、金油158和華雙5號,分別編號為RA01、RA02、RA03、RA04、RA05、RA06、RA07和 RA08。選取其中對堿性鹽抗性最強和最弱的2個油菜種質測定根際土壤有機酸含量。
試驗土壤采自內蒙古農業大學新區農場,田間持水量為25.4%。土壤理化性質為pH 8.08、電導率91.0μS/cm、有機質8.92g/kg、全氮0.70g/kg、全磷0.54g/kg、全鉀10.18g/kg、堿解氮59.61mg/kg、速效磷3.83mg/kg、速效鉀98.3mg/kg。
1.2 試驗設計
試驗在內蒙古農業大學農學院溫室進行,溫室的晝夜溫度為(25±2)℃和(20±2)℃,光照 14h/d,相對濕度 60%。采用盆栽方法,將 2種堿性鹽NaHCO3和Na2CO3(分析純)按摩爾比2:1混合,然后與土壤攪拌均勻后裝入內徑18.5cm、高23cm的花盆中。抗性指標篩選試驗中,設置0(A0,對照)和 4g/kg(A4)2個堿性鹽在土壤中的濃度處理,于2020年6月28日播種,8月3日取樣,篩選堿性鹽脅迫下耐性最強與最弱的油菜種質。
以篩選的耐性最強和最弱的種質為材料,設置4個土壤中混合堿性鹽的濃度,分別是0(A0,對照)、2(A2)、4(A4)和 6g/kg(A6)。于 2020 年9月10日播種,10月15日取樣。按照田間持水量達到90%計算每盆的澆水量,待達到適宜播種條件后播種。選取飽滿均勻的油菜種子播種3盆,每盆6穴,每穴3粒,齊苗后間苗,保證每盆6株。生長指標和根際土壤有機酸含量的測定均3次重復,每個重復4株幼苗。處理后土壤pH和電導率見表1。
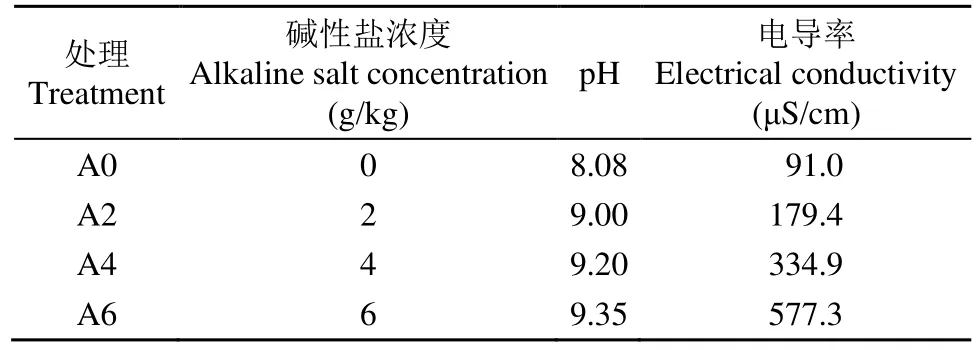
表1 處理后土壤pH和電導率Table 1 The pH and electrical conductivity in soil after treating
1.3 測定指標與方法
1.3.1 出苗率 播種后 7d統計出苗數。計算出苗率,出苗率(%)=(種子出苗數/供試種子總數)×100。
1.3.2 生長指標 取出植株,將植株清洗干凈,用紙巾擦干后使用電子游標卡尺測定莖粗,用直尺測量株高和主根長,將植株分為地上部和地下部,稱鮮重,然后在105℃殺青30min,再于80℃下烘干12h,稱干重。
1.3.3 根際土壤有機酸含量 抖下根系附著的疏松土壤并用毛刷收集與根系緊密結合的土壤,稱取10g土壤樣品,加入0.1%磷酸水溶液100mL,振蕩5min后在2000轉/min下離心30min,之后過水相0.45μm濾膜,利用島津LC-20AT高效液相色譜儀測定有機酸組分和含量。試驗用的草酸、蘋果酸、酒石酸、抗壞血酸、甲酸、乙酸、檸檬酸及琥珀酸標準品均為分析純。流動相:0.01mol/L磷酸氫二銨,用1mol/L磷酸調pH至2.70,用0.45μm濾膜過濾,使用前用超聲波脫氣;流速0.8mL/min;分析柱為 XAqua-C18(250mm×4.6mm,5μm);柱溫30℃;紫外檢測波長 210nm,帶寬 40nm;進樣量20μL。
1.4 數據處理
采用Microsoft Office Excel(2010)對試驗數據進行整理作圖。利用IBM SPSS Statistics 20.0統計軟件進行方差檢驗、相關性分析和主成分分析,同時采用隸屬函數法對主成分分析中得分系數較高的指標進行耐鹽堿性綜合評價,參照桑曉慧等[12]的方法計算。IA=Di /Si,μ(d)=(IA-IAmin)/(IAmax-IAmin),IA為耐鹽堿系數,μ(d)為隸屬函數值,Di為堿性鹽脅迫下的性狀值,Si為無脅迫下的性狀值,IAmin和IAmax分別為耐鹽堿系數的最小值和最大值。
2 結果與分析
2.1 堿性鹽脅迫下油菜種質耐鹽堿指標主成分分析
對8個油菜種質的8個生長指標進行主成分分析,通過確定具有代表性的指標來綜合評價其耐鹽堿能力。由表 2可知,2個主成分累計貢獻率為91.179%,可以解釋原8個變量的絕大部分信息,損失量僅為8.821%。第1主成分特征值為6.254,貢獻率為78.181%,與第1主成分緊密相關的因子包括出苗率、植株鮮重、植株干重、根干重、株高和主根長,得分系數分別為0.152、0.157、0.155、0.156、0.152和0.144,可綜合表現出堿性鹽脅迫下油菜苗期的生物量積累和生長狀況。第2主成分特征值為1.040,貢獻率為12.998%,莖粗載荷最大,得分系數為0.954。因此,可將出苗率、植株鮮重、植株干重、根干重、株高、主根長和莖粗作為油菜抗堿性鹽脅迫評價的特征因子。
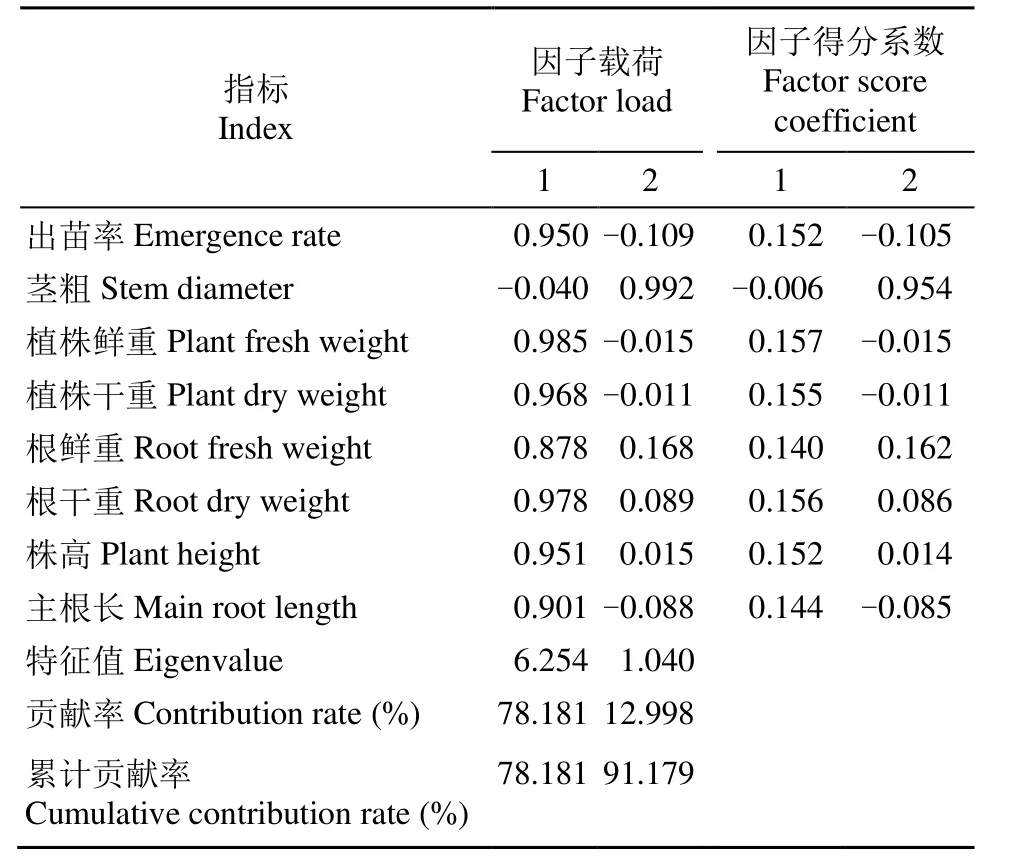
表2 堿性鹽脅迫下油菜種質各指標的主成分分析Table 2 Principal component analysis of various indexes of rapeseed germplasms under alkaline salt stress
2.2 油菜種質苗期指標的耐鹽堿系數
由表3可知,在堿性鹽脅迫下,8個油菜種質的出苗率、莖粗、植株鮮重、植株干重、根干重、株高和主根長的均值與對照相比均有所降低。各指標在種質間的變化和排序不同。

表3 8個油菜種質各指標的耐鹽堿系數Table 3 Mixed saline-alkali tolerance coefficients of each indicator of eight rapeseed germplasms
2.3 堿性鹽脅迫下油菜種質隸屬函數及排序
根據主成分分析,從8個指標中篩選出貢獻率較大的出苗率、莖粗、植株鮮重、植株干重、根干重、株高和主根長7項指標進行隸屬函數分析,求平均值,對8個油菜種質進行耐鹽堿性綜合評價,從結果(表4)看出,華油雜62耐鹽堿性最強,16-P32外最弱。耐鹽堿性綜合評價排序為華油雜62>華雙5號>崇2>16-P22外>青雜9號>金油158>17崇1>16-P32外。
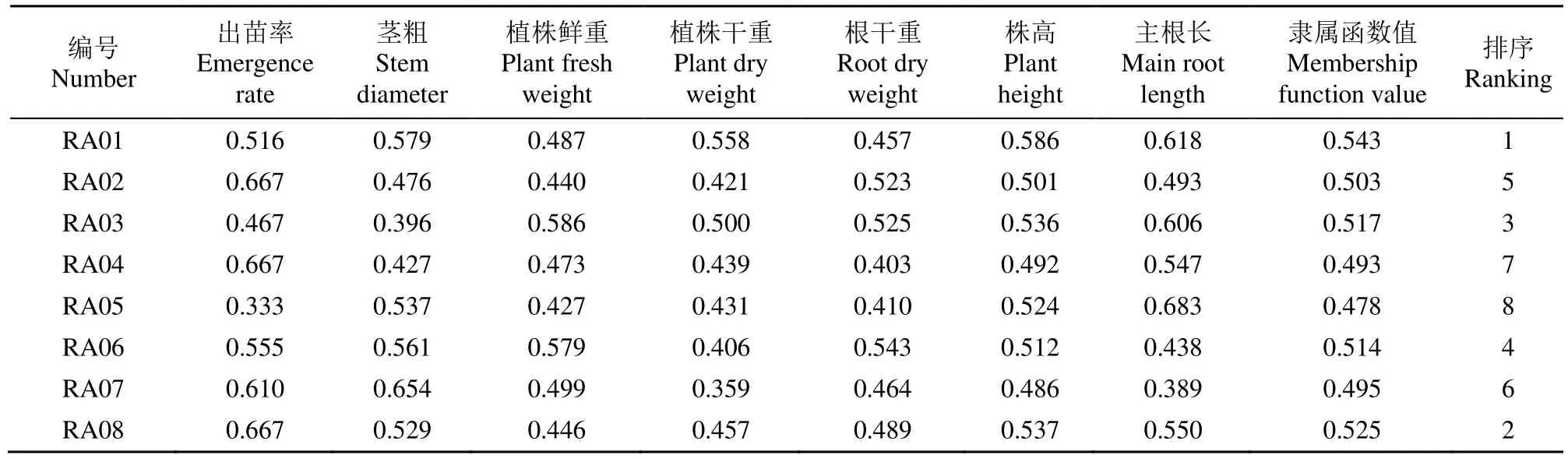
表4 堿性鹽脅迫下8個油菜種質隸屬函數及排序Table 4 Membership function and ranking of eight rapeseed germplasms under alkaline salt stress
2.4 堿性鹽脅迫對華油雜 62和 16-P32外苗期表型的影響
隨著堿性鹽濃度的增加,2個油菜種質生長速度減慢,植株弱小,其中華油雜62的表現好于16-P32外(圖1)。
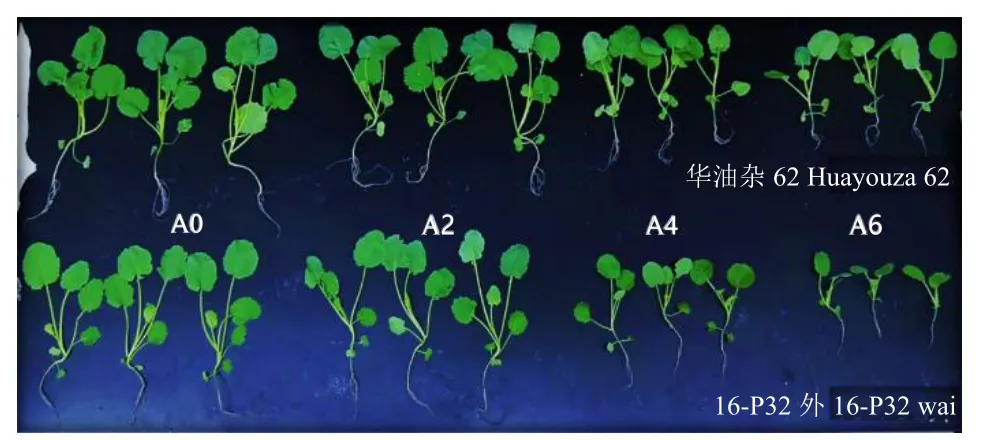
圖1 不同濃度堿性鹽脅迫下華油雜62和16-P32外的表型差異Fig.1 Differences in phenotypes of Huayouza 62 and 16-P32 wai under different alkaline salt concentrations
2.5 堿性鹽脅迫對華油雜 62和 16-P32外苗期根際土壤有機酸含量的影響
華油雜62和16-P32外根際土壤各有機酸含量均受到堿性鹽處理的顯著影響,并且處理和種質差異對油菜根際土壤草酸、琥珀酸和酒石酸有顯著的交互作用(表5)。2個油菜種質根際土壤中草酸、甲酸、檸檬酸和琥珀酸含量均有隨堿性鹽濃度增大而升高的趨勢,各處理琥珀酸含量與A0均有顯著差異,A4、A6處理檸檬酸含量與A0有顯著差異,華油雜 62各處理甲酸含量與A0均有顯著差異,16-P32外的甲酸含量僅A6處理較A0具有顯著差異。草酸是堿性鹽脅迫下2個油菜種質根際土壤中含量較多的有機酸,華油雜62和16-P32外在A6處理下的草酸含量分別占總有機酸含量的 59.34%和60.67%。華油雜62各處理的根際土壤草酸含量較A0分別顯著增加67.08%、176.53%和1229.54%,16-P32外分別顯著增加41.31%、55.74%和868.63%,表明高濃度堿性鹽能強烈誘導2個油菜種質根際土壤草酸含量增加。華油雜62根際土壤草酸含量在A0和A2處理下低于16-P32外,但A4、A6較高濃度堿性鹽處理下高于 16-P32外,甲酸含量僅在A6處理下低于16-P32外,檸檬酸和琥珀酸含量則是各處理均高于16-P32外。
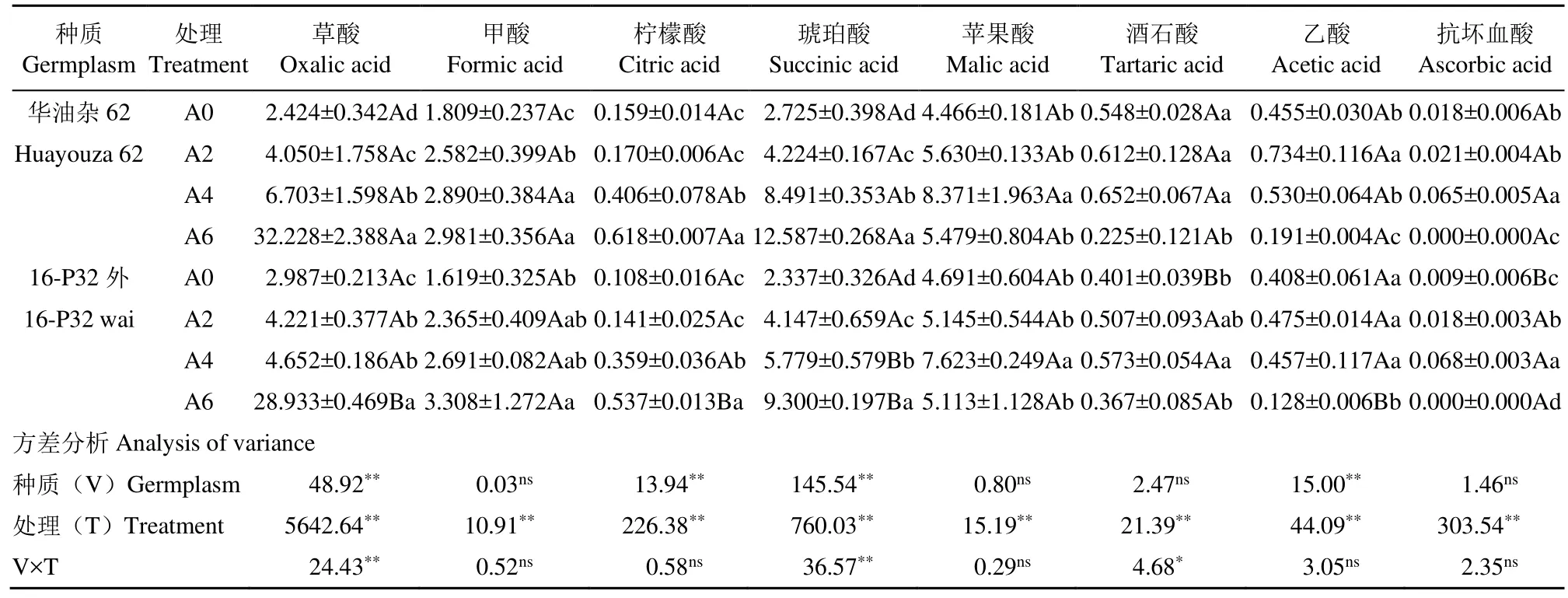
表5 堿性鹽脅迫下油菜根際土壤有機酸含量Table 5 Organic acid contents of rapeseed rhizosphere soil under alkaline salt stress μg/mL
由表5可知,2個油菜種質根際土壤中蘋果酸、酒石酸、乙酸和抗壞血酸含量在不同濃度堿性鹽脅迫下均表現出先升高再降低的趨勢,其中蘋果酸、酒石酸和抗壞血酸含量均在A4處理下達到峰值,華油雜62分別達到8.371、0.652和0.065μg/mL,較A0分別增加 87.44%、18.98%和 261.11%,16-P32外分別達到7.623、0.573、0.068μg/mL,較A0處理分別增加62.50%、42.89%和655.55%。乙酸含量在A2處理下達到峰值,華油雜62和16-P32外分別為0.734和0.475μg/mL,較A0分別增加61.32%和16.42%。抗壞血酸含量很低,華油雜62根際土壤蘋果酸含量在A0處理下低于16-P-32外,在A2、A4和A6處理下均高于16-P32外;酒石酸含量僅在A6處理下低于16-P32外;乙酸含量則在各處理下均高于16-P32外。
3 討論
3.1 堿性鹽脅迫下油菜生長指標的變化
鹽堿脅迫使作物生長速度減慢、抑制作物根系吸收水分,從而導致其形態發生變化,因此作物的生長指標可作為耐鹽堿指標[24-26]。作物對鹽堿的抗性受多種因素的影響,用單一指標不能準確反映和評價植物的耐鹽堿能力。研究發現,株高、生物量和發芽率可作為玉米萌發期耐鹽堿鑒定指標[27];根長、生物量和株高可作為無芒雀麥苗期耐鹽堿鑒定指標[13]。本研究中使用多個指標綜合評價 8份油菜種質的耐鹽堿能力,通過主成分分析篩選貢獻率較大的指標,對其進行隸屬函數分析來評價油菜種質耐鹽堿性,與王婧澤等[28]的研究方法一致。
3.2 堿性鹽脅迫下油菜苗期根際土壤有機酸含量的變化
有機酸是一類在線粒體三羧酸循環中產生的帶有羧基的低分子化合物,其合成、積累、轉運及分泌對逆境條件下植物適應環境調節方面具有重要作用[29]。研究[30]表明,當植物受到堿性鹽脅迫時,植物會通過提高根部有機酸分泌量和質子分泌能力來維持體內pH穩定及離子平衡。本研究從8份油菜種質中篩選出對堿性鹽耐性最強和最弱的2個油菜種質華油雜62和16-P32外,測定其根際土壤有機酸含量,發現草酸的積累量最高。有機酸中的草酸不僅可以通過絡合作用來達到緩解鋁脅迫的目的[31],還可以改變鹽堿土壤的理化性質,降低土壤 pH。通過澆灌草酸可以幫助盆栽葡萄緩解鹽堿脅迫[32]。結合本研究中油菜根際土壤草酸的大量積累,推測澆灌草酸對油菜適應鹽堿脅迫同樣具有一定作用。高濃度堿性鹽脅迫導致2個油菜種質根際土壤中草酸、甲酸、檸檬酸和琥珀酸4種有機酸含量較A0均顯著提高,這可能是因為油菜根系分泌的草酸、甲酸、檸檬酸和琥珀酸可以平衡堿性鹽脅迫下過多的陽離子,維持了植物pH穩定,蘋果酸、酒石酸和乙酸含量隨著堿性鹽濃度的升高呈先升高后下降的趨勢,說明低濃度堿性鹽脅迫下油菜根系可以做出快速反應,通過分泌有機酸來應對高pH傷害,而高濃度堿性鹽脅迫會傷害油菜根系,使根系的代謝能力大大降低,從而抑制有機酸的合成。根外pH的調節也受根系分泌有機酸的影響,本研究中隨著堿性鹽濃度的增加,酒石酸和乙酸含量驟然下降或許也有堿性鹽抑制2個油菜種質根系分泌有機酸活力的原因。華油雜62根際土壤的草酸、蘋果酸、檸檬酸、琥珀酸和乙酸含量在高濃度堿性鹽脅迫下高于 16-P32外,酒石酸和甲酸含量低于16-P32外,說明在堿性鹽脅迫下,2個油菜種質根際土壤有機酸對于降低 pH傷害有不同的側重,不同油菜種質對適應堿性鹽脅迫有不同的生理表現。
星星草對堿脅迫特殊的生理反應是有機酸中檸檬酸的積累[19];沙棘在堿脅迫下蘋果酸和檸檬酸含量較低,乙酸含量有很大提高[8]。本試驗中,2個油菜種質在高濃度堿性鹽處理后以分泌草酸為主,故推測草酸可能在油菜響應鹽堿脅迫方面起的作用更大。草酸為堿地膚[33]和堿蓬[34]在應對鹽堿脅迫時體內主要的有機酸,表明有機酸代謝的調節在不同植物中可能發揮不同的作用。
在堿性鹽脅迫下,多種有機酸和總有機酸含量上升,抗性強與抗性弱種質表現不同。有關有機酸的積累機制等有待于進一步研究。
4 結論
對主成分分析中得分系數較高的指標出苗率、莖粗、植株鮮重、植株干重、根干重、株高和主根長等采用隸屬函數法進行耐鹽堿性綜合評價,排序為華油雜62>華雙5號>崇2>16-P22外>青雜9號>金油158>17崇1>16-P32外。堿性鹽脅迫下油菜有機酸合成增加,以緩解堿性鹽脅迫帶來的pH傷害,在草酸、酒石酸、甲酸、蘋果酸、抗壞血酸、乙酸、檸檬酸及琥珀酸8種有機酸中,草酸的積累量最高,而抗壞血酸的積累量很低。除酒石酸和乙酸含量在A6處理下較A0有所下降外,其余各有機酸含量均上升。高濃度堿性鹽脅迫下華油雜62的草酸、蘋果酸、檸檬酸、琥珀酸和乙酸含量高于16-P32外,酒石酸和甲酸含量低于16-P32外。分泌有機酸是油菜調節根外pH的重要途徑,是油菜應對堿性鹽脅迫的重要生理響應。
