特早熟甘藍型春油菜恢復系核心種質構建
2022-02-18 07:23:42林曉陽杜德志柳海東
作物雜志 2022年1期
林曉陽 杜德志 柳海東 李 鈞
(青海大學農林科學院/春油菜遺傳改良重點實驗室/國家油菜改良中心青海分中心/農業農村部春油菜科學觀測實驗站/青海省春油菜研究開發中心/青海省春油菜工程技術研究中心,810016,青海西寧)
油菜作為我國重要的油料作物,每年種植面積可達667萬hm2(1億畝)[1-2]。春油菜屬于一年生草本植物,種植區域主要分布在青海、甘肅、新疆、內蒙古及西藏等氣溫較低、無霜期較短的地區,約占我國油菜種植面積的10%~15%[3-5]。在我國高海拔、無霜期較短的春油菜區,油菜種植主要以白菜型油菜為主,存在產量低、品質差的問題;甘藍型油菜(Brassica napus L.)較白菜型油菜產量高、品質優,但生育期較長制約了其在高海拔地區的推廣種植。選育能在高海拔地區正常成熟的高產優質甘藍型油菜品種對提高高海拔地區春油菜產量具有重要意義。
甘藍型油菜的F1代具有較強的雜種優勢。自傅廷棟等[6]發現波里馬雄性不育系以來,恢復系的合理選擇和應用成為三系配套雜交種選育的一個重要環節。種質資源作為新品種選育的遺傳基礎,通過構建核心種質,可以用部分樣本代表遺傳資源群體,為親本的雜交組配提供理論指導[7]。目前,我國已構建了水稻[8]、大豆[9]和小麥[10]等主要農作物核心種質。核心親本作為核心種質的子集,主要是為組配強優勢雜交種做準備。同時其篩選并不是一成不變的,不同育種目標有不同側重點。隨著具有更優良性狀品種的選育,可以不斷擴充和更新具有目標性狀的核心種質庫,增強優勢雜交親本的組配幾率,從而提高育種效率[11]。核心種質的建立主要借助于 SSR(simple sequence repeat)和 SNP(single nucleotide polymorphism)等分子標記手段。曾曉珊等[12]利用48對SSR引物構建了27個水稻核心種質的指紋圖譜,從中篩選到的優勢核心種質均為秈稻;楊勇[13]利用173對SSR引物構建了甘藍型油菜核心種質的指紋圖譜。上述研究為秈稻和甘藍型油菜的雜交組合及雜種優勢的預測提供了科學指導。
構建核心種質,重要的是如何從群體中選出核心子集。對于給定的群體,已經提出了許多用于核心集的算法。如Brown提出的僅取決于群體大小而不取決于每個群體多樣性的 P方法和 L方法;Schoen和 Brown提出的 M 方法等[14]。而 Core Hunter算法直接從整個群體中采樣,依托 REMC(replica exchange monte carlo)本地搜索技術,可以防止搜索陷入局部最優狀態;同時,可自主選擇優化多個度量,并進行權重的分配,進而找到具有較高親本間平均遺傳距離和較高總體多樣性的核心集;與其他算法相比,該方法靈活、高效,且所保留的核心子集明顯較小[15]。用能在高海拔地區正常成熟的優質特早熟甘藍型油菜資源作為親本,杜德志等[16]選育出“青雜3號”、“青雜4號”、“青雜7號”、“青雜8號”和“青雜10號”5個特早熟甘藍型油菜雜交種,提高了高海拔地區油菜的產量及品質。選擇優良的特早熟甘藍型油菜恢復系對特早熟甘藍型油菜品種的培育具有重要意義。本研究在簡化基因組測序技術的基礎上,利用 Core Hunter算法對97份特早熟甘藍型油菜恢復系進行核心種質的構建,可以更好地對特早熟甘藍型春油菜恢復系種質資源進行評價和利用,減少育種工作中恢復系與不育系選配雜交組合的工作量,同時為特早熟甘藍型春油菜優良組合的親本選配提供恢復系資源。
1 材料與方法
1.1 試驗材料
材料來自青海省農林科學院春油菜研究所。供試的97份材料為早熟甘藍型油菜品系與青藏高原白菜型油菜“門源小油菜”進行種間雜交(母本為甘藍型D9946、父本為白菜型浩油11號),其中F1代在帳子內進行自由授粉,在下一代中選取偏甘藍型植株進行多代自交,即后代系譜選育而成的特早熟甘藍型春油菜恢復系品系。供試材料初花期在42~50d。
1.2 田間試驗
2019年4月25日,將97份恢復系播于青海省農林科學院春油菜研究所試驗田(101°45′ E,36°43′ N),采用隨機區組設計,每個恢復系3個重復,每個重復2行區,行長2m,行間距30cm,株距15cm。
1.3 測序分析
1.3.1 酶切方案設計 對97份材料幼嫩葉片進行取樣 1~2cm2,送至北京百邁客生物科技有限公司進行測序分析。選取甘藍型油菜寧油 7號基因組(http://ibi.zju.edu.cn/bnpedigome/download.php?con=ny7)為參考基因組進行酶切預測,組裝基因組大小為993.78Mb,GC含量36.32%。
1.3.2 Illumina平臺測序及產出數據的質量分析
酶切預測后,選取合適的酶切組合對97份基因樣品進行酶切。將酶切后片段(specific-locus amplified fragment標簽)的3′端加ploy A、連接 Dual-index[17]測序接頭,經過特定的PCR引物擴增后,進行純化處理再混合,然后將擴增產物進行瓊脂糖凝膠電泳進行分離,選取目的片段進行切膠,經文庫質檢合格后選用水稻日本晴(Oryza sativa ssp. japonica)作為對照,將測序得到的數據在Illumina測序平臺進行測序分析。
1.3.3 SLAF標簽和SNP標記開發 對測序得到的數據進行酶切效率評估。利用 SOAP[18]軟件將Control測序中產生的讀長(reads)與甘藍型油菜參考基因組作比對。根據生物信息學分析結果,利用BWA[19]將SLAF-seq測序得到的reads以油菜基因組作為參考基因組進行比對,利用 GATK[20]和SAMtools[21]2種方法開發全基因組范圍的SNP標記,選取基因型頻率第2高的基因型所在頻率(一般MAF>0.05)[22]作為具有代表性的SNP,將二者交集作為最終可靠的SNP標記庫。
1.3.4 核心種質構建及評價指標 利用 Core Hunter軟件構建該群體的核心種質并進行評價[23],從改良的羅杰斯距離(MR)、愛德華和卡瓦利距離(CE)、香農指數(SH)、期望雜合度(HE)、多態性信息含量(PIC)、有效等位基因(NE)和等位基因覆蓋度(CV)等方面進行考量。MR將每個等位基因視為一個單獨的維度,是標準歐幾里德距離的改進,CE則采用選擇性漂移模型,樣本的突變率低且選擇性壓力快速變化;SH是1種合適的度量,試圖盡可能多地保留子集中的稀有等位基因;HE將每個基因座內的多樣性考慮在內;CV表示核心子集中的等位基因與原始集合相比所占的百分比;PN與CV相反,是未覆蓋的等位基因[19]。
2 結果與分析
2.1 酶切建庫與測序評估
確定以RsaⅠ+HaeⅢ為酶切組合,在Illumina測序平臺以水稻日本晴作為對照進行測序分析,得到大小為364~414bp的SLAF標簽119 741個,各染色體上SLAF標簽的數量分布見圖1,SLAF標簽在基因組中分布大致均勻。結果顯示,1條序列兩端在參考基因組上的比對跨度介于50~1000bp的讀長占總讀長雙端比對的比對效率為95.16%;1條序列兩端在參考基因組上的比對跨度小于50bp,或大于1kb的讀長占總讀長的比例為1.21%;未比對到基因組上的讀長占總讀長的比例為3.63%;比對效率基本正常。與水稻日本晴數據雙端比對的比對效率為 95.16%,酶切效率為 87.54%,因此 SLAF建庫正常。
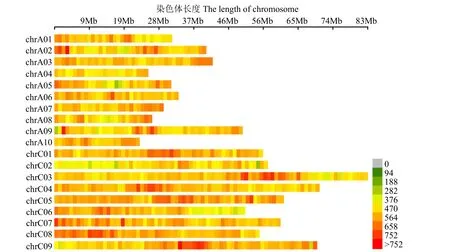
圖1 SLAF標簽在染色體上的分布Fig.1 SLAF tags distribution on chromosome
為保證項目數據分析可靠,采用去掉接頭126bp×2的讀長進行數據分析和評估。共得到367.5Mb讀長數據,測序平均Q30(堿基測序出錯概率為0.001)為94.70%;平均GC含量43.27%。用于評估試驗建庫準確性的水稻日本晴測序獲得0.43Mb的讀長數據量。測序質量值越高對應的堿基測序錯誤率越低,故此次建庫及測序合理[23]。
2.2 SLAF標簽與SNP標記的鑒定
選擇油菜基因組為參考基因組進行電子酶切預測,共得到249.91Mb讀長,獲得527 872個SLAF標簽,標簽的平均測序深度為 14.91X,其中多態性的SLAF標簽共有171 414個,以及842 248個群體SNP。SLAF標簽在A、C基因組上分別有174 963和306 011個(表1);C基因組上標簽數量明顯多于A基因組,約為A基因組的2倍。利用A、C基因組分別開發多態性SLAF標簽70 441個和86 086個。
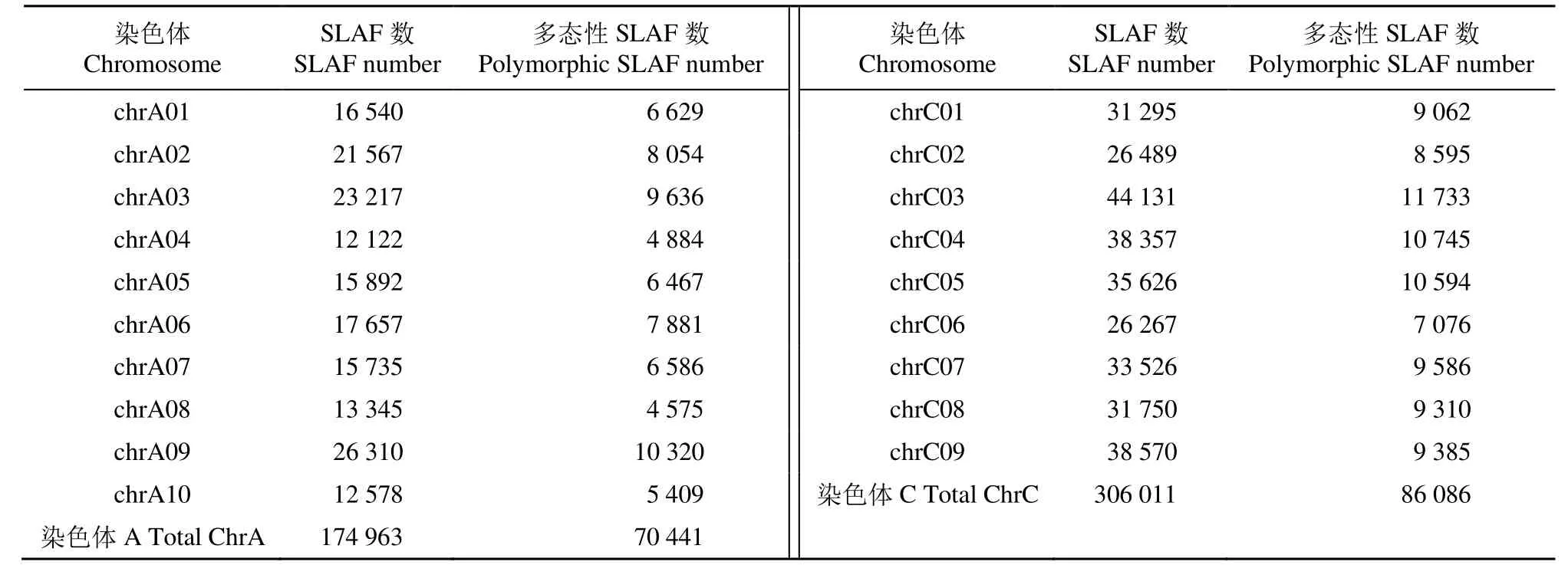
表1 SLAF標簽和多態性SLAF標簽染色體分布Table 1 Chromosome distribution of SLAF tags and polymorphic SLAF tags
共獲得842 248個SNP標記,根據SNP在染色體上的分布,繪制SNP在染色體上的分布圖(圖2)。A基因組上SNP分布明顯較C基因組更為富集。此外,SNP完整度介于23.77%~38.73%;雜合率介于5.20%~15.33%,純合度較高。
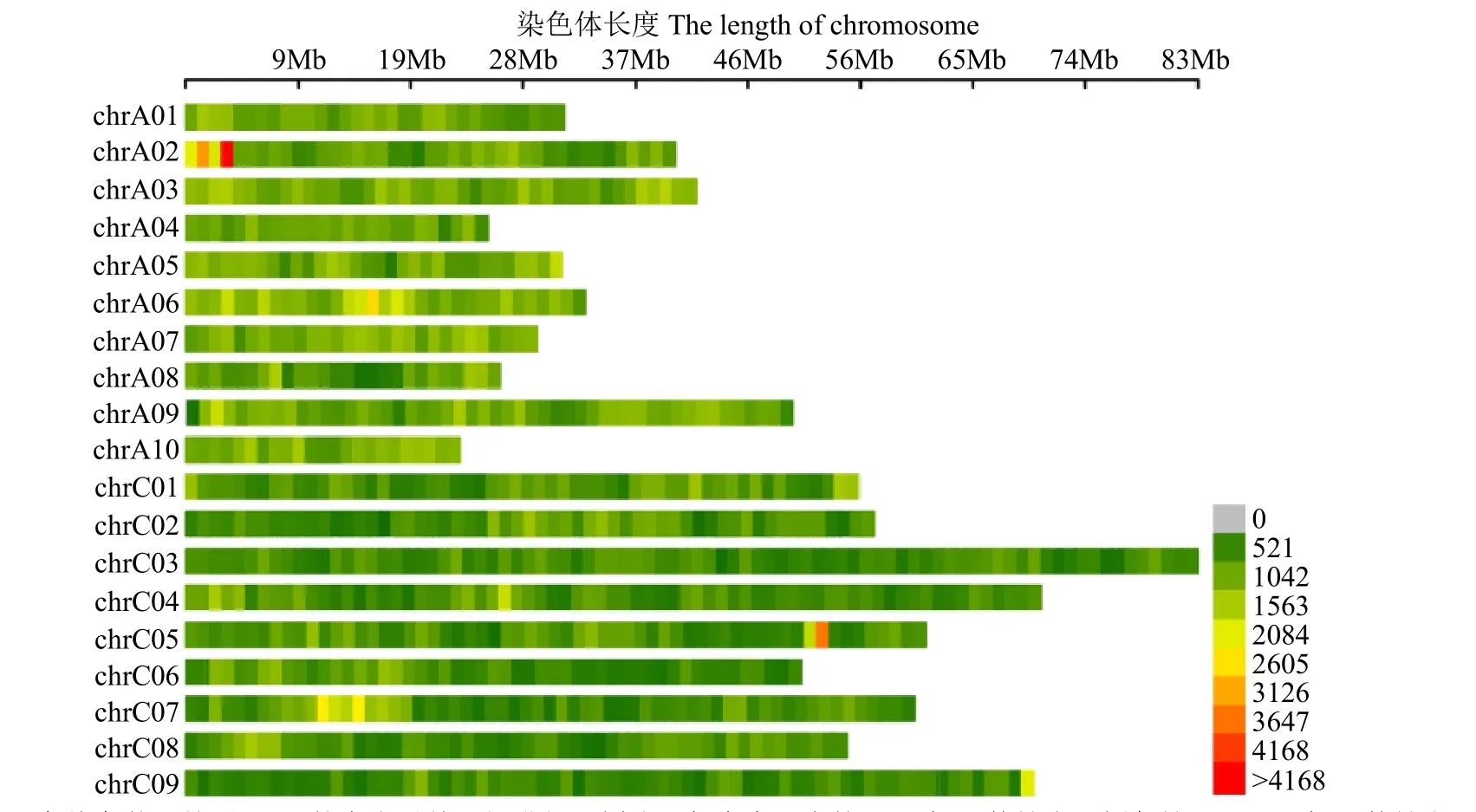
圖2 SNP在染色體上的分布Fig.2 SNP distribution on chromosome
2.3 系統發育和遺傳結構分析
使用MEGA X[24]軟件對得到的SNP位點進行遺傳分析,基于neighbor-joining的算法,在bootstrap重復1000次、Kimura 2-parameter模型下,構建97份特早熟甘藍型油菜種質資源的系統發育樹,分析不同種質之間的遺傳進化關系(圖3)和群體結構。97份材料可分為5類:第1類只包含1份種質;第2類包含26份種質;第3類包含17份種質;第4類包含17份種質;第5類包含36份種質。
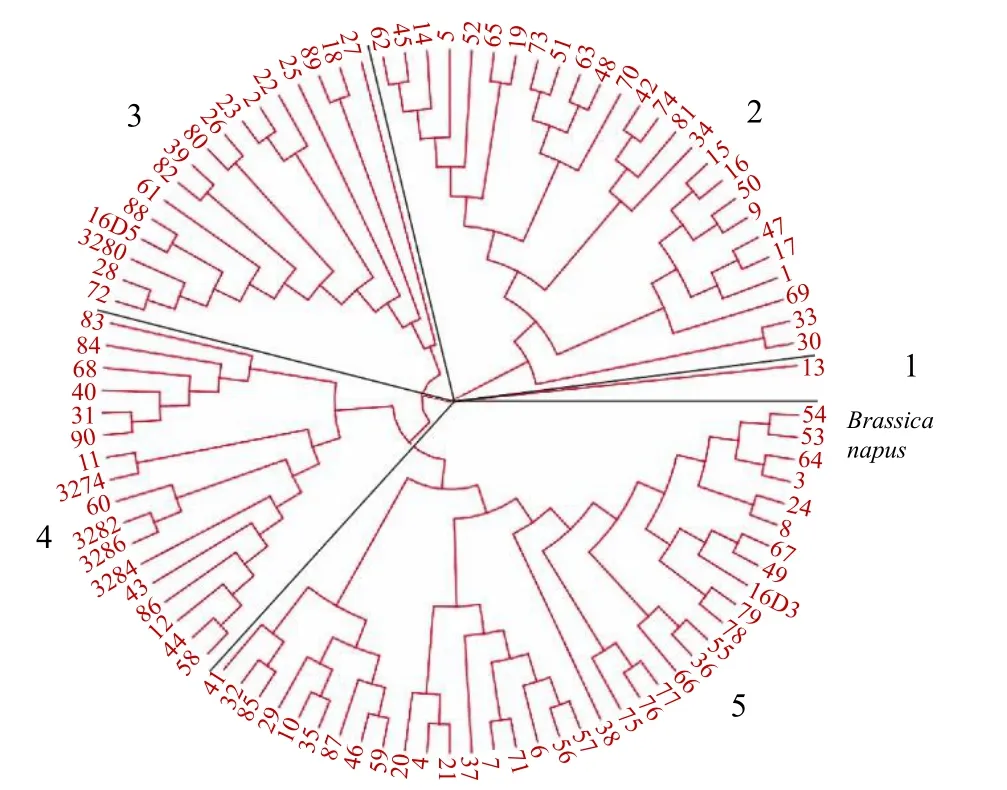
圖3 基于SLAF-seq技術的甘藍型春油菜恢復系NJ進化樹Fig.3 NJ phylogenetic tree of Brassica napus L. restorer based on SLAF-seq
2.4 恢復系間遺傳距離分析
基于neighbor-joining算法計算97份甘藍型油菜恢復系之間的遺傳距離。遺傳距離介于 0.0026~0.6628,平均遺傳距離為0.3510。由表2可知,97份恢復系間排列前10位的遺傳距離值均≥0.60。可以看出,遺傳距離前10位的組合主要集中于編號86、12、44、58 和編號 10、78、16D3、49、67 材料之間的組合。
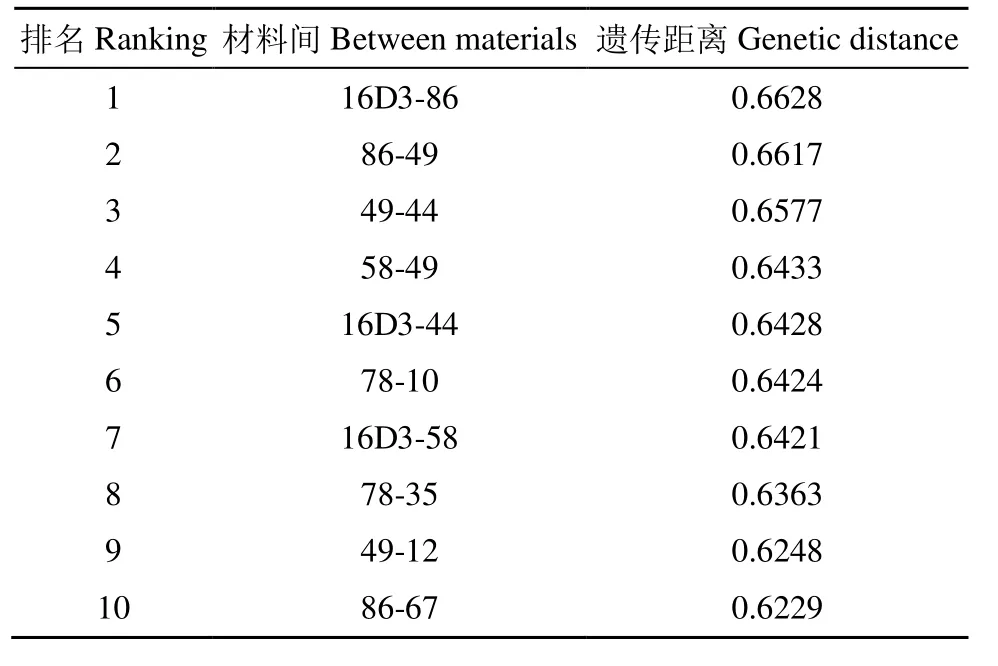
表2 甘藍型油菜各恢復系之間排位前10位的遺傳距離Table 2 Genetic distances of the top 10 restorer lines in Brassica napus L.
2.5 核心種質構建與評價
根據有效SNP,利用Core Hunter軟件構建97份甘藍型油菜恢復系核心種質,得到2個較優核心子集,分別命名為C30和C40。其中,C30包括29份恢復系資源(圖4),C40包括38份恢復系資源(圖5)。
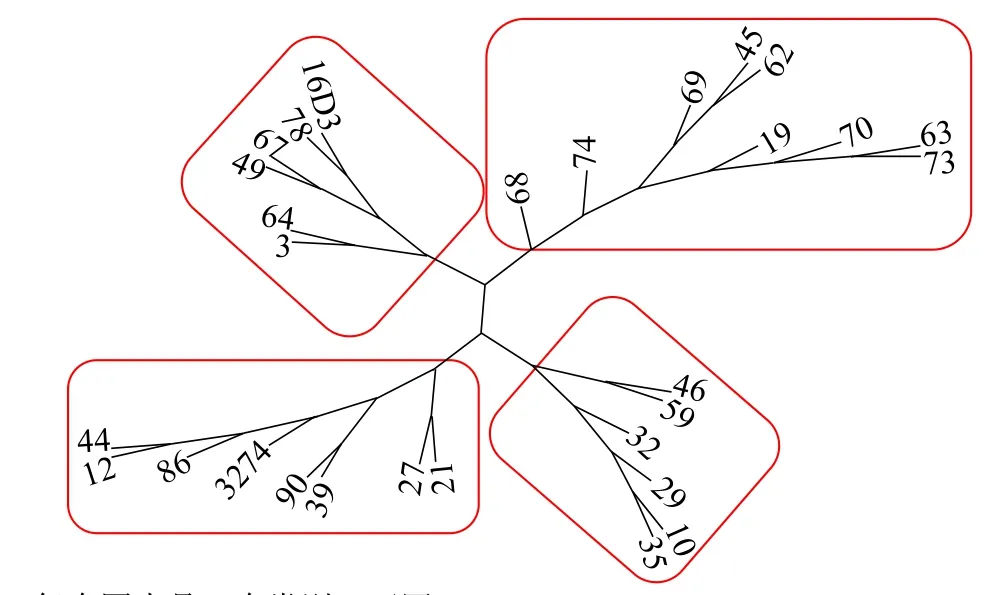
圖4 C30核心種質聚類圖Fig.4 Clustering map of C30 core collection
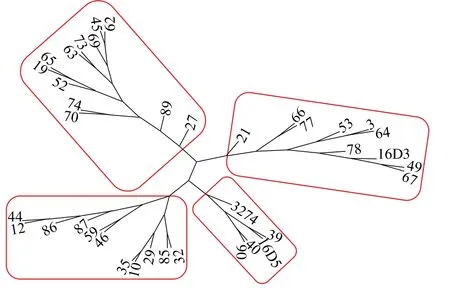
圖5 C40核心種質聚類圖Fig.5 Clustering map of C40 core collection
由表3可知,C30和C40核心子集的PIC分別為0.20和0.29。其中,C30核心種質的MR大小在0.15~0.47范圍內,CE范圍為0.16~0.48,2個子集相差不大。2個核心子集的HE相同,均為0.33;SH均為11.35。CV分別為99.89%(C30)和99.96%(C40),均達到 99%以上。進一步對C30和 C40進行聚類分析,2個核心子集均可分為4類(圖4-圖5)。原始樣品聚類類型中編號13的樣品單列一類,未被選入C30和C40核心種質群中;其他4類所包含材料類型和原始材料間聚類基本一致。因此,C30和C40這2個核心子集分別所包含的樣品可以代表97份優良甘藍型油菜恢復系的核心種質(表3);由于2個核心子集的各項數據差異不大,且C30子集的材料均被包含在C40中,最終將C30作為核心子集。這些結論為后期最優雜交組合配制中種質的選配提供依據。
3 討論
核心種質的篩選有助于更好地利用現有種質資源,核心種質群離不開核心種質的構建。構建核心種質即從整體中選取一定數量較少的材料代表一個作物種及其近緣野生種的最大遺傳多樣性[25]。對于油菜核心種質的研究,國內外主要通過形態分析和分子標記的技術手段來獲得大量的遺傳背景信息。新疆農業大學在2001年以RAPD資料建立起新疆部分栽培油菜的核心資源[26];何余堂等[27]從199個白菜型油菜資源中初步建立了含有 28個品種的核心資源;趙緒濤等[23]對118份甘藍型油菜進行核心種質的構建。本研究通過SLAF-seq技術獲得了249.91Mb的讀長數據,開發得到527 872個SLAF標簽和842 248個群體SNP,為提高育種遺傳效率和開發特早熟甘藍型油菜特異性 SNP提供參考,可進一步加快甘藍型油菜的遺傳育種改良。
雖然97份材料來源于同一組合,但5類材料之間仍具有一定的遺傳差異,可指導雜交育種,遺傳差異大的材料雜交后代可能具有超親優勢。使用Core Hunter軟件,得到了C30和C40這2個核心子集,從各項指數看,這2個子集都能夠代表原始材料,最終選取C30子集作為核心子集。但是甘藍型油菜相比于其他2種類型的油菜,具有明顯較小的遺傳差異。因此局限于材料的來源,為更好地增加遺傳多樣性,還需要繼續擴寬親本特早熟甘藍型油菜的種質資源,增加遺傳豐度。
青海特早熟甘藍型油菜較其他地區油菜品種能夠早熟1個月,具有在2900m海拔以上且年平均溫度在0.5℃以下的區域種植的特點,比該地區主栽品種浩油11號增產約10%[28]。特早熟甘藍型油菜的育種目標是要使植株對光、溫的需求接近白菜型油菜的水平,其中生育期是主要的育種目標[29],在特早熟甘藍型油菜育種中選取開花期作為熟性選擇的指標[30]。目前在我國高海拔地區已經選育出了特早熟甘藍型油菜品種“青雜4號”和“青雜7號”,“青雜4號”適合種植在海拔2900~3100m的地區,“青雜7號”適合種植在海拔2750~2950m的地區,但在3100m以上的高海拔地區仍然主栽白菜型油菜,因此,選育更早熟的甘藍型油菜品種對于這些地區油菜品種改良具有重要意義。
4 結論
通過 SLAF-seq對 97份特早熟甘藍型油菜恢復系構建了核心親本子集,有利于特早熟恢復系種質資源的保存、評價和利用,從而減少育種工作中恢復系與不育系選配雜交組合的工作量,為生產中代替高海拔地區白菜型油菜的優良特早熟甘藍型油菜雜交種提供更多可能和選擇,擴大春油菜區甘藍型油菜的種植區域,以及提高高海拔地區油菜的產量和品質。
利用SNP構建核心親本子集,得到包含29份材料的C30子集和包含38份材料的C40子集,通過各項數據表明,這2個核心子集能代表整個資源的群體遺傳多樣性水平,并最終將C30子集作為核心子集。構建了97份特早熟甘藍型油菜恢復系的核心種質,為種質資源的開發、利用和保存提供了參考。
