外源硫化氫對成齡桃樹根系生長的影響
2022-02-08 11:58:32高曉蘭孫茂祥吳雪蓮肖元松彭福田
山東農業科學 2022年12期
高曉蘭,孫茂祥,吳雪蓮,肖元松,彭福田
(1.山東省果樹研究所,山東 泰安 271000;2.山東農業大學園藝科學與工程學院,山東 泰安 271018)
桃樹根系主要分布在表層土壤中,以水平生長為主,沒有明顯的主根,同級根系分枝粗細相近。桃樹根系的水平分布范圍一般與樹冠冠徑相近或略廣,垂直分布大部分根系在1 m以內。但桃樹根系的分布受環境條件的影響較大,在不同環境中表現出明顯的差異。
硫化氫(H2S)是繼一氧化氮(NO)和一氧化碳(CO)之后第3個被發現的氣體信號分子[1],參與干旱脅迫下十字花科植物光合作用的調節[2],而且可與植物激素交叉調控種子的萌發[3]。近年研究發現,硫化氫能夠調控植物根系的生長發育。李東波等[4]發現低濃度外源硫化氫可以提高根尖邊緣細胞存活率、根尖組織可溶性蛋白含量,進而促進豌豆胚根生長。硫化氫預處理通過減少根系ROS的積累和保護質膜完整性,減少根尖細胞的死亡來緩解Cd脅迫對根長的抑制,減少由ROS積累引起的細胞死亡[5]。Li等[6]研究表明硫化氫可與生長素(auxin,IAA)相互作用,調節甘薯和番茄植株的不定根和側根形成。硫化氫不僅對根系的伸長具有劑量效應,而且與植物側根的發生相關[7]。Lin等[8]研究發現,NaHS通過CO產生酶血紅素加氧酶-1(haem oxygenase-1,HO-1)誘導黃瓜不定根的發生。Fang等[9]發現硫化氫參與生長素誘導的番茄側根形成。Jia等[10]發現硫化氫通過調節ABP調節肌動蛋白依賴性生長素運輸引起擬南芥中根發育的改變。Mei等[7]研究表明,硫化氫通過過氧化氫誘導番茄幼苗側根形成。
桃樹根系生長狀況對養分和水分的吸收利用至關重要,硫化氫參與調控植物生長發育的很多生命過程。筆者前期研究發現,外源硫化氫可促進桃實生苗根系的生長[11]。但在大田條件下,硫化氫對成齡桃樹根系生長的作用效果尚不明確。為此,本試驗以硫化氫對根系生長的調控作用為切入點,在前期試驗的基礎上,以7年生‘瑞光33/毛桃’為試材,研究外源硫化氫對成齡桃樹根系生長的影響,同時利用實時熒光定量PCR技術,檢測外源硫化氫對桃實生苗根系生長素合成編碼基因YUC1和YUC2及生長素轉運編碼基因AUX1、PIN1、PIN2和PIN3的表達水平,探討硫化氫調控桃樹生長發育的作用效果,最終明確外源硫化氫對成齡桃樹根系生長的調控作用,以期為桃樹栽培生理研究提供有益參考。
1 材料與方法
1.1 外源硫化氫對成齡桃樹根系生長的影響試驗
1.1.1 試驗設計與處理方法 于2018年6月至10月在大田條件下進行。在前期研究篩選出硫化氫供體NaHS適宜濃度為0.2 mmol·L-1的基礎上,以7年生‘瑞光33/毛桃’為試材,供試土壤為粘壤,設置2個處理:0.2 mmol·L-1NaHS(NaHS)、清水對照(CK)。每隔15天處理一次,共處理5次,每處理7次重復,單株小區。每次使用0.2 mmol·L-1的NaHS(對照用3 L清水)3 L澆灌在樹干兩側30 cm處選取的40 cm見方的土壤中,為防止水分和釋放的硫化氫散失,處理后用聚乙烯薄膜覆蓋處理位置的土壤表面。
1.1.2 測定指標與方法 處理結束后進行破壞性取樣,取出整個根系,沖洗干凈,使用專業版WinRHIZO根系分析系統測定根系構型參數,包括總表面積(cm2)、根尖數、總體積(cm3)、分枝數、交叉數、根系平均直徑(cm)、側根數量、根系總長度(cm)。每處理重復3次,結果取平均值。
測定結束后,根系按直徑大小進行分級并解析(<2 mm為細根,2~5 mm為中等粗根,>5 mm為粗根),置于烘箱中烘干,測定各類根系的干重,每處理3次重復,結果取平均值。
1.2 外源硫化氫對桃根系生長素合成及轉運編碼基因表達的影響試驗
1.2.1 試驗設計與處理方法 為了研究外源硫化氫對桃根系生長素合成轉運及細胞周轉蛋白編碼基因的影響,以長勢基本一致、無病害的5片葉毛桃實生苗為試驗材料,幼苗定植于盆中,盆為圓柱形,內徑24 cm,高12.8 cm,盆中基質為石英砂。設置CK(每盆澆施100 mL的清水,對照)、NaHS(每盆澆施100 mL 0.2 mmol·L-1的硫氫化鈉)、HT(每盆澆施100 mL 0.2 mmol·L-1的硫化氫清除劑亞牛磺酸)3個處理,每處理30株。處理2h后,將各處理根系取樣清洗干凈后,放置在-80℃冰箱中備用。利用實時熒光定量PCR技術測定根系生長素合成編碼基因YUC1和YUC2及生長素轉運編碼基因AUX1、PIN1、PIN2和PIN3的表達水平。
1.2.2 實時熒光定量檢測 采用EASYspin Plus植物RNA快速提取試劑盒(北京愛德萊生物科技有限公司)提取樣品RNA,利用反轉錄試劑盒(Perfect Real Time,TaKaRa)獲得cDNA。實時熒光定量PCR采用SYBR Green PCR PremixEx Taq(寶生物公司),操作參照說明書。使用CFX96 Touch Real-Time PCR Detection System(Bio-Rad,USA)進行PCR反應。反應程序為:95℃預變性30 s;95℃變性5 s,退火30 s(退火溫度根據設計的引物溫度而定),退火延伸65℃5 s,40個循環。所有反應設3次技術學重復,所得數據采用2-ΔΔCt進行計算分析。引物參見表1。

表1 引物信息
1.3 數據分析
試驗數據采用Microsoft Excel進行圖表繪制,采用DPS軟件對數據進行差異顯著性檢驗(Duncan’s新復極差法,P<0.05)。采用Beacon Designer 7軟件進行熒光定量引物設計。
2 結果與分析
2.1 外源硫化氫對成齡桃樹根系表型性狀的影響
2.1.1 對根系表型結構參數的影響 由表2可以看出,外源硫化氫可在一定程度上促進成齡桃樹根系的生長。與CK相比,外源硫化氫處理成齡桃樹的根系總長度、根系總表面積、根系總體積、根尖數、分枝數和交叉數分別提高了12.79%、19.26%、12.86%、12.96%、10.83%和25.28%,差異顯著;根系平均直徑降低了17.46%,差異顯著,反映了硫化氫處理促進了成齡桃樹細根的發生。并且,硫化氫處理顯著提高了成齡桃樹的根系活力(圖1)。
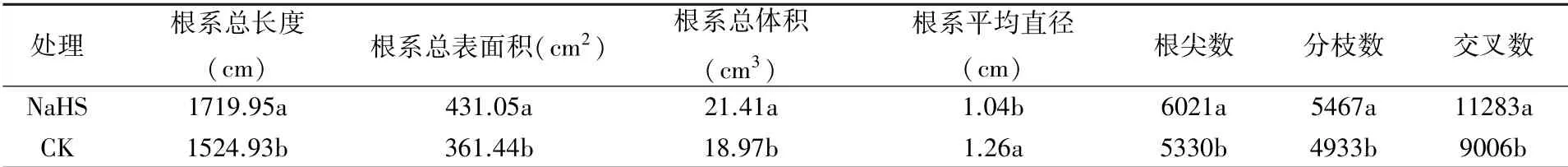
表2 外源硫化氫對成齡桃樹根系生長的影響

圖1 外源硫化氫對成齡桃樹根系生長活力的影響
2.1.2 對不同粗度根表型性狀的影響 與對照相比,外源硫化氫處理顯著提高成齡桃樹直徑0~2.0 mm的細根長度、表面積、體積,分別提高了15.54%、27.88%、33.52%,差異顯著,但對直徑2.0~4.0 mm的中粗根和>4.0 mm的粗根長度、表面積、體積影響不顯著(圖2~圖4)。

圖2 外源硫化氫對成齡桃樹不同直徑根系長度的影響
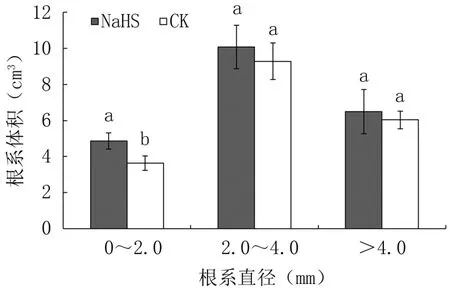
圖4 外源硫化氫對成齡桃樹不同直徑根系體積的影響
2.1.3 對根系分形維數的影響 外源硫化氫處理后成齡桃樹根系分形維數較對照提高了2.89%,差異顯著(圖5)。表明外源硫化氫處理可使成齡桃樹根系復雜程度變大。

圖3 外源硫化氫對成齡桃樹不同直徑根系表面積的影響

圖5 外源硫化氫對成齡桃樹根系分形維數的影響
2.2 外源硫化氫對成齡桃樹根系干物質積累量的影響
由圖6可以看出,外源硫化氫處理成齡桃樹直徑0~2.0 mm的細根干物質積累量與對照相比顯著提高36.22%;但對直徑2.0~4.0 mm的中粗根和>4.0 mm的粗根的干物質積累量影響不顯著。可見,在成齡桃樹上施用外源硫化氫當年主要促進直徑0~2.0 mm細根的生長。
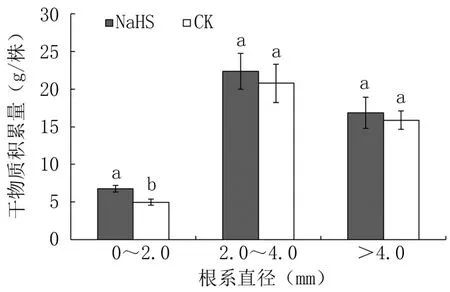
圖6 外源硫化氫對成齡桃樹根系干物質積累量的影響
2.3 外源硫化氫對桃根系生長素合成及轉運編碼基因表達的影響
外源硫化氫處理桃實生苗2 h后,根系生長素合成編碼基因YUC1和YUC2上調表達(圖7);根系生長素轉運編碼基因AUX1、PIN1、PIN2、PIN3的表達量也上調,尤其PIN1上調表達幅度較高(圖8)。表明外源硫化氫可以通過影響根系生長素合成和轉運編碼基因的表達來影響桃實生苗根系的生長。

圖7 外源硫化氫對桃根系生長素合成編碼基因表達水平的影響

圖8 外源硫化氫對桃根系生長素轉運編碼基因表達水平的影響
3 討論與結論
李東波等[4]發現用低濃度外源硫化氫處理豌豆能夠使根尖組織可溶性蛋白含量升高,根尖邊緣細胞存活率上升,粘膠層相對面積變小,從而促進豌豆胚根生長。本研究結果也表明,施用0.2 mmol·L-1NaHS可顯著提高大田條件下7年生‘瑞光33/毛桃’成齡桃樹根系總長度、根系總表面積、根系總體積、根尖數、分枝樹和交叉數,與對照相比分別提高12.79%、19.26%、12.86%、12.96%、10.83%和25.28%,根系平均直徑降低了17.46%,差異均達顯著水平;尤其對細根生長促進明顯,能使直徑0~2.0 mm細根的長度、表面積、體積和干物質積累量分別顯著提高15.54%、27.88%、33.52%和36.22%;外源硫化氫處理后成齡桃樹根系分形維數較對照顯著提高2.89%。由于本試驗只進行了一季,時間較短,對根系的長期影響效果可能還未完全顯現,且大田條件下成齡桃樹根系生長對NaHS的敏感性要小于實驗室條件,因此今后有必要對外源硫化氫影響成齡桃樹根系生長的長期效果進行深入研究。
Zhang等[12]研究發現NaHS處理能夠誘導切根番薯的不定根發生,而加入生長素轉運抑制劑和NO清除劑能夠逆轉IAA和INO對不定根的誘導作用,推測硫化氫可能通過IAA和NO來誘導植物不定根的發生。PIN蛋白是生長素運輸所必需的,PIN2蛋白是IAA向基運輸的關鍵調節因子,PIN1蛋白調控IAA的向頂運輸。PIN1和PIN2都對根系構型的形成具有重要作用[13]。在PIN系列中,PIN1、PIN3、PIN4和PIN7調節植物側根的形成[13]。pin1突變體和pin3pin4pin7三重突變體表現出側根彎曲程度明顯減弱[14,15]。有研究表明,當擬南芥根系內源性硫化氫水平升高時,PIN1、PIN2和PIN7的表達增強[10]。另外,控制細胞周期進展的下游分子有助于擬南芥的側根發生[16,17]。在水稻中,細胞周期調節基因ORC3突變破壞細胞周期進程并阻斷主根上側根的發生[18]。并且生長素可通過調節根系外周細胞中細胞周期調控基因如CYCA2;1、CYCA3;1、CYCD3;1和CDKA1的轉錄調控側根的形成[19,20]。
本研究發現,外源硫化氫處理桃實生苗后,根系中生長素合成編碼基因YUC1、YUC2及生長素轉運編碼基因AUX1、PIN1、PIN2、PIN3均上調表達,表明外源硫化氫可能通過影響桃根系中生長素的合成和轉運來調控根系生長,但其具體調控機制還有待進一步研究。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
