甘藍型油菜初花期不同漬水時長對產量及產量因子的影響
2022-02-08 08:19:38郭一鳴王同華劉新紅
湖南農業(yè)科學 2022年12期
關鍵詞:產量
郭一鳴,王同華,劉新紅,李 寶,周 興,李 莓
(湖南省作物研究所,油菜雜種優(yōu)勢利用湖南省工程研究中心,湖南 長沙 410125)
漬澇是一類全球發(fā)生的環(huán)境脅迫,全球氣候變化導致越來越頻繁和嚴重的漬澇災害[1]。近5 a以來,我國每年因漬水危害引起的農業(yè)損失僅次于干旱,導致農作物減產高達15%~80%。油菜作為我國重要的油料作物之一,常年種植面積733.3萬hm2,為我國提供了約55%的食用植物油[2]。然而,在長江流域主產區(qū)普遍采用水稻—油菜水旱輪作和稻茬免耕種植模式,由于土壤本身黏重、透氣性差,春秋季節(jié)多降水等氣候特點,再加上近10 a來極端降水現(xiàn)象明顯增多,油菜種植期間極易發(fā)生漬澇災害,尤其是低洼地塊。漬澇災害易導致根系長期缺氧,對油菜生長發(fā)育影響很大,一般可造成17.0%~42.4%甚至更大的產量損失[3]。
大田出現(xiàn)漬澇災害時,水稻等耐受型植物在乙烯因子、脫落酸、赤霉素等激素應激響應下,根系上部可通過啟動細胞程序性死亡不斷生成通氣組織,并迅速擴張使氧氣擴散阻力下降,同時積極生成高位的不定根或替代根來保障長期淹水條件下根系氧氣的獲得,從而有效提高了其耐漬澇的能力[4-5]。而大豆、油菜等水分敏感型作物在形成通氣組織的同時,主要解決辦法是啟動應急通道,包括快速調動抗氧化酶等相關基因產生抗氧化因子,盡量減少活性氧等成分的大量生成、堆積而造成毒害作用,同時通過減少基本生理需求,包括淀粉代謝管理,降低呼吸作用、光合作用等來保存能量[6]。因此,耐漬能力主要體現(xiàn)在毒性物質的耐受力以及能量保障上。油菜對漬澇的耐受性遺傳變異廣泛,不同類型和不同地域的油菜種質資源存在顯著差異。湖南省地處長江中游地區(qū),春季多降水,花期遭受漬澇的概率大,相應的遺傳變異豐富,有利于耐漬型油菜種質資源的篩選及其遺傳資源的挖掘探索。前人對油菜漬澇的研究以苗期為主,并建立了以存活率、葉綠素含量、光合效率、可溶性糖及脯氨酸等相應的方法和指標[7-9]。然而,相對于恢復期和補償期長的種子萌發(fā)和苗期階段,開花期對漬水脅迫更加敏感,并直接影響一次分枝數(shù)、有效角果數(shù)、千粒重等產量構成因子,最終影響產量[10]。花期油菜葉片不同光譜區(qū)域對漬水脅迫等響應不同,因此可利用光譜指數(shù)曲線等變化對耐受型材料進行鑒別[11]。耐受型材料在花期遭遇漬水后相對于敏感型材料產量損失顯著降低[12-13],其在脅迫過程中通過應激感應調控并作用到植物生長和產量因子上的效應更顯著,且不同脅迫時間和時長對于耐受型材料的鑒定和后期相關主效基因的挖掘也有影響。為了探究花期淹水脅迫下不同甘藍型油菜材料的調節(jié)機制,筆者以湖南省主推甘藍型油菜品種的親本為材料,研究了初花期不同時長漬水處理對產量及產量因子的影響,以期為耐漬型油菜品種的選育提供參考。
1 材料與方法
1.1 供試材料
選用11份湖南省主推油菜品種的親本材料為研究對象,各供試材料的編號和代碼見表1。
表1 11份親本材料的實驗編號和材料代碼
育苗基質由湖南百威生物科技股份有限公司提供。供試盆栽規(guī)格為盆高22 cm,盆口直徑25 cm,盆底直徑17 cm。
1.2 試驗方法
試驗在湖南省作物研究所玻璃大棚進行。首先將大田黏土與育苗基質按照體積比1∶1混合,然后裝入花盆中進行育苗。各親本材料分別于2020年10月中旬和11月上旬播種。播種后置于有遮擋、能通風的玻璃大棚中,按時澆水、定期施肥,確保親本材料健康生長。出苗后開始間苗,每盆留苗5棵,3~4葉期定苗至每盆留苗2棵。于次年2月開花時選擇生長一致性好的材料進行漬水處理。漬水處理于親本材料群體開花后第2天開始,以水沒過土壤表面1 cm為標準。漬水時長設7和5 d 2個處理,以不漬水為對照(CK),CK每2 d補充1次水分。每處理設10次重復,按隨機區(qū)組排列。漬水處理結束后按常規(guī)方法進行水肥管理,直至收獲。
1.3 測定指標與數(shù)據(jù)分析
油菜成熟后測定各處理的株高、單株有效角果數(shù)、角果長度、每角果粒數(shù)、單株產量,并按公式(1)計算各指標的相對值。
各指標相對值(%)=各指標處理值/對照值×100 (1)
1.4 數(shù)據(jù)統(tǒng)計分析
采用Excel 2010軟件和R軟件v3.10進行數(shù)據(jù)統(tǒng)計分析。
2 結果與分析
2.1 花期漬水對油菜產量及其相關因子的影響
花期漬水處理7 d后,11份親本材料的株高比CK下降24.7%~40.0%,差異均達到顯著或極顯著水平;單株有效角果數(shù)比CK下降63.6%~92.9%,差異也均達到顯著或極顯著水平;角果長比CK下降6.3%~41.2%,除YS01、YS03、YS06、YS10和YS11外其他親本的差異均達到極顯著水平;每角果粒數(shù)比CK下降41.0%~83.8%,差異均達到顯著或極顯著水平;單株產量比CK下降69.1%~99.0%,差異均達到極顯著水平,其中YS08下降最少、YS09下降最多(表2)。
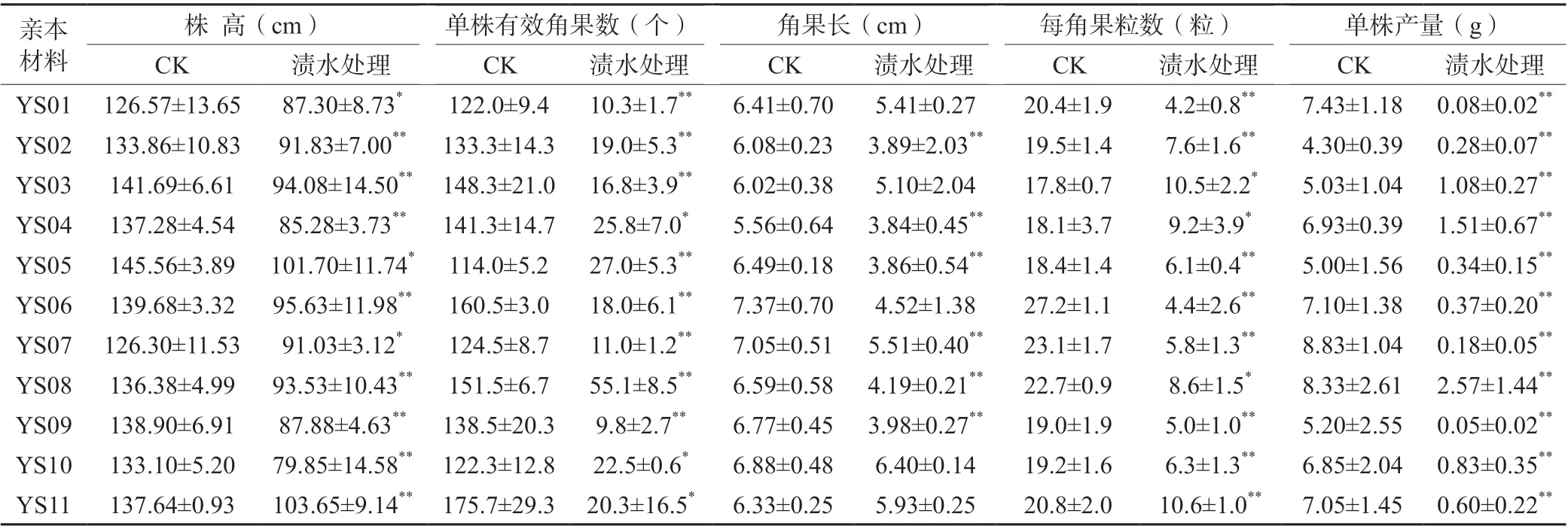
表2 花期漬水7 d對各親本材料產量及其相關因子的影響
花期漬水處理5 d后,11份親本材料的株高比CK下降18.2%~40.8%,差異均達到顯著或極顯著水平;單株有效角果數(shù)比CK下降31.2%~90.9%,除YS10外差異均達到顯著或極顯著水平;除YS11漬水處理角果長比CK增加了13.4%外,其他材料漬水處理的角果長比CK下降0~28.0%,但差異均未達顯著水平;每角果粒數(shù)比CK下降8.3%~46.6%,除YS03、YS05、YS08外其他親本材料的差異均達到顯著或極顯著水平;單株產量比CK下降26.7%~89.9%,除YS08和YS10外差異均達到顯著或極顯著水平,其中YS10下降最少、YS09下降最多(表3)。
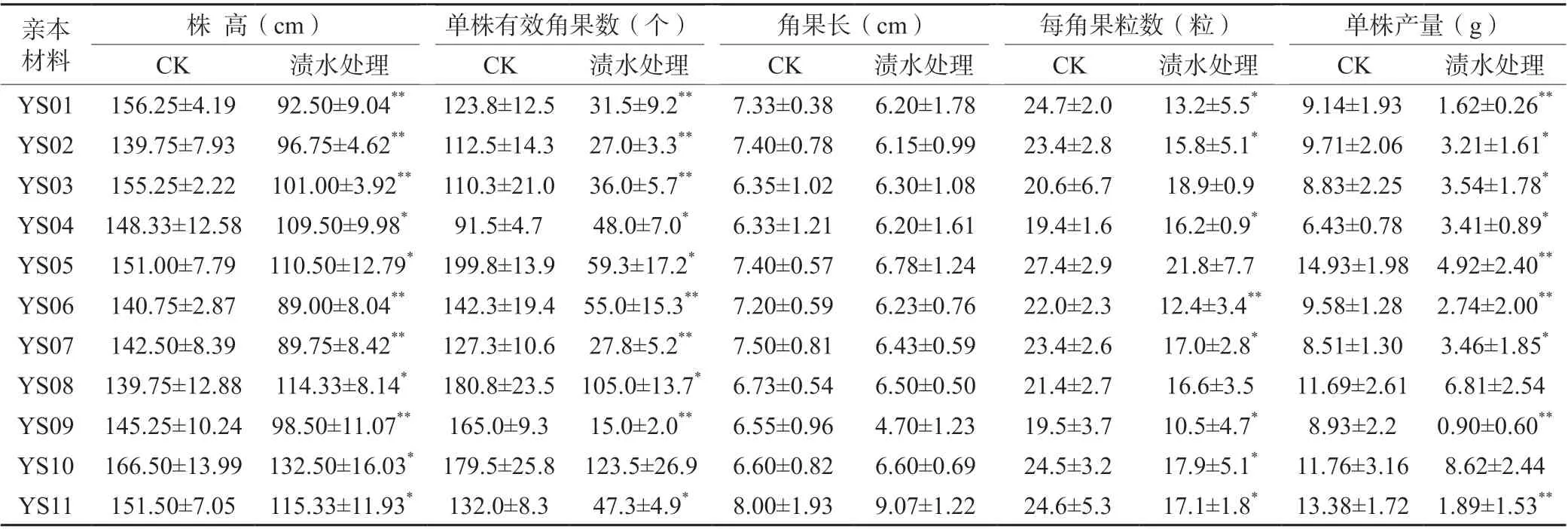
表3 花期漬水5 d對各親本材料產量及其相關因子的影響
2.2 各親本材料花期漬水處理后產量及其相關因子的比較
花期漬水處理7 d對產量及各相關因子的影響比處理5 d顯著。花期漬水處理7和5 d后,各親本材料的平均株高分別下降32.4%和29.8%,各親本材料的2個處理間均沒有顯著性差異(圖1);平均單株有效角果數(shù)分別下降84.6%和64.0%,其中親本材料YS03、YS04、YS10和YS11的2個處理間差異達到顯著或極顯著水平(圖2);平均角果長分別下降26.3%和8.1%,其中親本材料YS08的2個處理間差異達到極顯著水平(圖3);平均每角果粒數(shù)分別下降64.4%和29.2%,其中親本材料YS05、YS06、YS07、YS09和YS10的2個處理間差異達到顯著或極顯著水平(圖4);平均單株產量分別下降89.3%和63.5%,其中親本材料YS04、YS07和YS10的2個處理間差異達到顯著水平(圖5)。
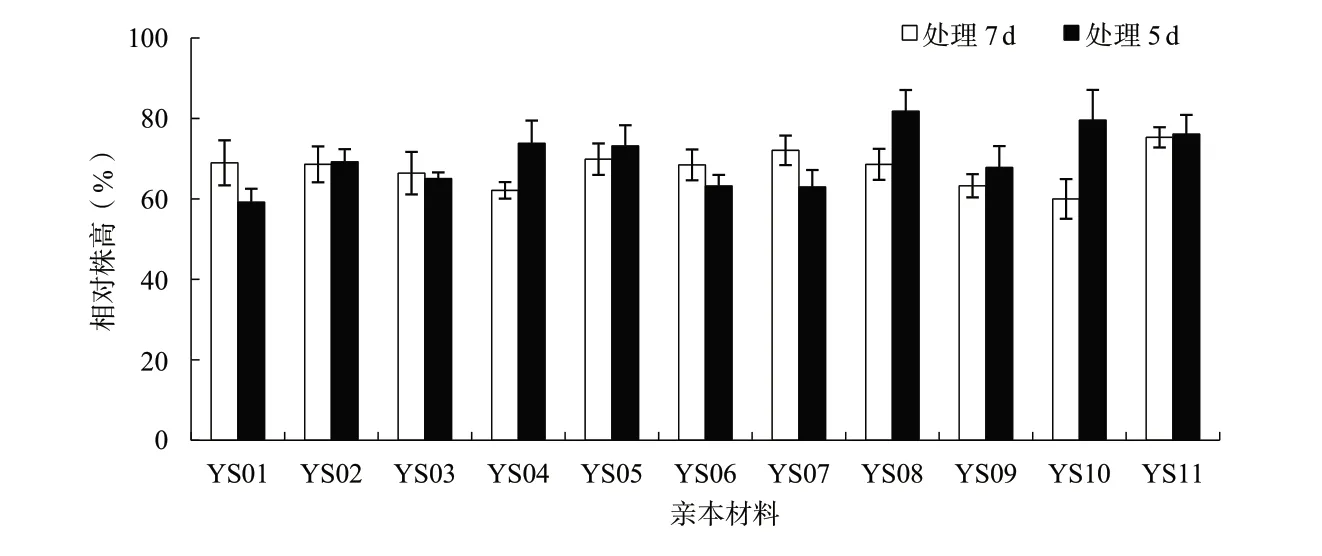
圖1 各親本材料花期漬水處理7和5 d后相對株高的比較
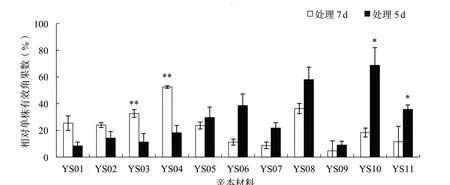
圖2 各親本材料花期漬水處理7和5 d后相對單株有效角果數(shù)的比較
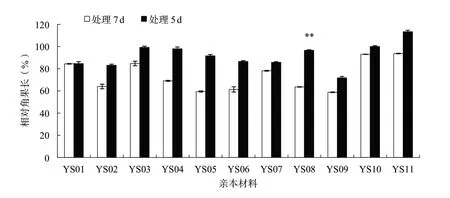
圖3 各親本材料花期漬水處理7和5 d后相對角果長的比較
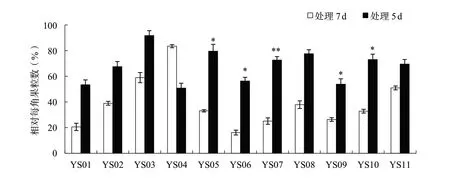
圖4 各親本材料花期漬水處理7和5 d后相對每角果粒數(shù)的比較
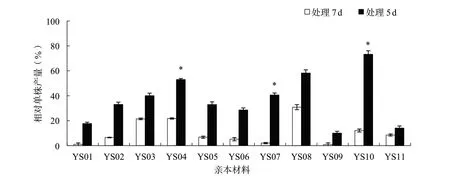
圖5 各親本材料花期漬水處理7和5 d后相對單株產量的比較
2.3 各親本材料花期不同漬水時長的產量及其相關因子方差分析
如表4所示,花期漬水處理7和5 d的親本材料間和漬水處理間的單株產量及產量相關因子(株高、單株有效角果數(shù)、角果長、每角果粒數(shù))均存在顯著或極顯著差異。
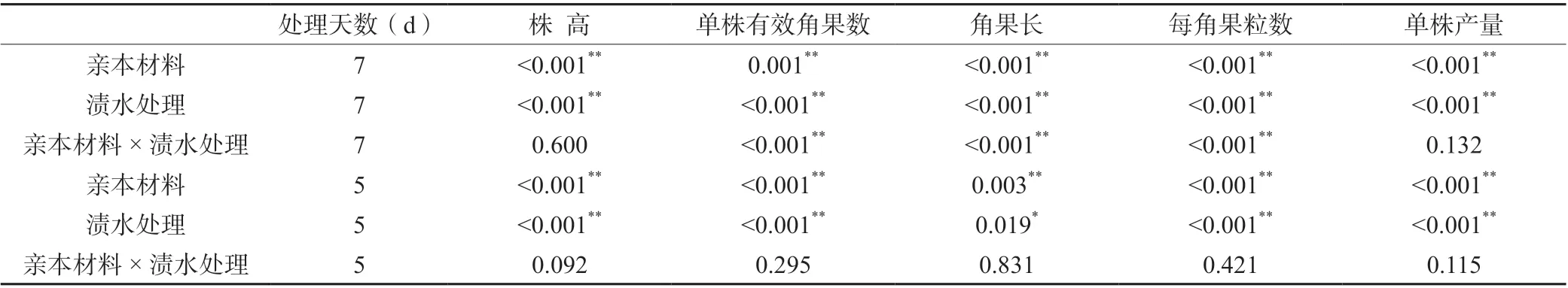
表4 各親本材料花期不同漬水時長的產量及相關因子方差分析
2.4 花期漬水處理7和5 d后的產量及其相關因子的相關性分析
如表5所示,花期漬水處理7 d后,單株產量與單株有效角果數(shù)和每角果粒數(shù)呈極顯著正相關,但與其他因子的相關性不顯著;而花期漬水處理5 d后,單株產量與株高、單株有效角果數(shù)和每角果粒數(shù)均呈顯著或極顯著正相關,同時單株有效角果數(shù)與株高和角果長、角果長與每角果粒數(shù)也存在顯著正相關。

表 5 各親本材料花期漬水處理7和5 d后的產量及其相關因子的相關性
3 討 論
油菜作為我國南方稻田的主要輪作作物,其生產在花期遭受漬澇脅迫的概率較大,篩選花期耐漬水的種質資源是選育油菜花期耐漬水品種的基礎。相比于油菜苗期漬水處理7 d以上后期生長的反彈能力[9-10],筆者的試驗結果表明,花期處理5 d已足以對角果長以外的單株產量及其相關因子造成顯著或極顯著影響,花期漬水處理5 d后各親本材料的平均產量減產63.5%;而處理7 d后的平均產量減產達89.3%,甚至個別處理的植株出現(xiàn)死亡,說明漬水7 d已臨近閾值。同時,花期漬水處理5 d后的產量與各產量因子之間均有顯著的相關性,而處理7 d后的產量與各產量因子間的相關系數(shù)下降,這可能是由于漬水處理時間過長所致,因此盆栽漬水處理的時長為5 d時比較理想。此外,油菜在花期遭遇非生物逆境脅迫應答后會在轉錄、蛋白和代謝水平發(fā)生一系列復雜變化,這些代謝反應及其調控并不是孤立的,往往由不同基因形成復雜的通路和網(wǎng)絡,為增強逆境耐受能力,生殖器官發(fā)育相關基因的表達可能受阻[14]。因此,油菜花期漬水脅迫后期可以考察生殖器官的基因表達與調控路徑等,為進一步挖掘油菜花期相關基因提供參考。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(yè)(2021年14期)2021-11-25 23:57:29
今日農業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(yè)(2020年20期)2020-11-26 06:09:10
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
