衰老對2種獼猴血液基因表達的影響及與人類的對比
2022-01-26 06:47:18王鑫頎嚴超超王嬌蘭月范振鑫四川大學生命科學學院生物資源與生態環境教育部重點實驗室成都60065中國科學院成都生物研究所中國科學院山地生態恢復與生物資源利用重點實驗室生態恢復與生物多樣性保護四川省重點實驗室成都6004四川大學生命科學學院四川省瀕危野生動物保護生物學重點實驗室成都60065
四川動物 2022年1期
王鑫頎, 嚴超超, 王嬌, 蘭月, 3, 范振鑫*(. 四川大學生命科學學院, 生物資源與生態環境教育部重點實驗室, 成都60065; . 中國科學院成都生物研究所, 中國科學院山地生態恢復與生物資源利用重點實驗室, 生態恢復與生物多樣性保護四川省重點實驗室, 成都6004; 3. 四川大學生命科學學院, 四川省瀕危野生動物保護生物學重點實驗室, 成都60065)
衰老是一個系統的過程,表現為逐漸減慢的新陳代謝和細胞器官水平的衰老,并伴隨著許多生理指標的變化(Lópezotín.,2013)。年齡是心臟病、癌癥和中風等高死亡率疾病的主要風險因素,也是許多慢性病最重要的風險因素(van den Akker.,2014;Peters.,2015)。衰老主要受遺傳和環境因素的影響。目前對衰老遺傳機制的研究主要來自模式物種和長壽人群,例如和基因的突變被認為與中國漢族人的壽命有關(Li.,2009)。遺傳可能只在人類壽命中發揮15%~30%的決定作用,其余的影響則來自環境因素(Ruby.,2018b)。
獼猴屬是分布最廣的非人靈長類動物,也是人類疾病研究和臨床藥物開發領域中最重要的動物模型之一(Lan.,2020;Yan.,2020)。老年的獼猴屬動物會患上多種人類老年疾病,如心臟病和癌癥等(Justice.,2016);普通獼猴(CR)和人類在衰老過程中有許多相似之處,如毛發顏色變淺變薄、身體脂肪重新分布、皮膚和肌肉張力降低以及活力喪失等(Mattison.,2017)。此外,隨著年齡增長,CR還會出現糖尿病、腫瘤、肌肉萎縮、骨質流失、免疫功能改變和認知能力下降等癥狀(Ngwenya.,2015)。對CR的研究還發現,營養限制對延緩衰老的機制在靈長類動物中保守(Mattison.,2012),這種機制可能對人類也同樣適用(Colman.,2014;Mattison.,2017;Simpson.,2017)。藏酋猴(TM)是中國特有種,是個體最大的獼猴屬物種,也是亞洲最大的猴科Cercopithecidae動物之一(Smith.,2010),是潛在的良好的醫學模式動物,其肥胖個體易出現高脂血癥等疾病(楊鋒等,2010)。
轉錄組研究發現,基因調控和表達模式的變化在物種進化中起重要作用,而高通量測序技術為研究衰老機制提供了一種新的有效方法(Lan.,2020)。在不同的轉錄組衰老研究中,只有小部分涉及了與年齡相關的基因集的交叉比較,這無疑增加了構建統一的與年齡相關標記基因的難度。與年齡相關的基因集在不同組織和物種的功能水平上的表達有許多相似之處,這表明通過功能分類研究年齡相關的基因集比分析單個基因更有可能揭示衰老過程的基因表達變化(Zahn.,2006;Passtoors.,2008)。目前獼猴屬轉錄組比較研究沒有涉及2個不同的物種年齡相關轉錄組的對比,而是對比了不同組織間的差異,如Chaves等(2020)對食蟹猴的腦微血管系統、內皮連接介質轉錄組進行了比較。
本研究收集了本團隊之前研究的CRs和TMs隨年齡變化的轉錄組數據(Yan.,2020),系統比較了2種獼猴在年齡增長過程中基因表達的共性和異性,同時比較了這2種獼猴和人類年齡變化相關的血液轉錄組數據,分析了它們和人類在衰老過程中基因表達的異同,為認識靈長類物種在衰老過程中的基因表達變化特征提供數據。
2 材料和方法
2.1 數據采集與預處理
CRs和TMs血液組織轉錄組數據共52個樣本(7雌21雄CRs和12雌12雄TMs):CRs年齡為1.5~20歲:1.5~5歲7個、6~10歲10個、11~15歲6個、16~20歲5個;TMs年齡為3~18歲:3歲6個、5歲6個、8歲3個、10歲2個、11歲 1個、15歲4個、17歲1個、18歲1個。
下載與CR和TM的年齡相關差異表達基因(differential expressed genes,DEGs),其中,CR 2 138個(閾值:<0.05;933個上調和1 205個下調),TM 2 523個[假發現率(false positive rate,FDR)<0.05;1 358個上調和1 165個下調]。此外,從人類血液組織轉錄組研究(Peters.,2015)(共14 983個樣本)中下載得到了1 497個(600個上調和897個下調)與人類年齡相關的DEGs(閾值:<2.2E-5)。在上述3項研究的差異表達分析中,年齡均被視為連續變量。
2.2 年齡相關DEGs的種間比較
使用VennDiagram包對基因集進行比較并繪制Venn圖(Chen,2018)。由于同源基因在各物種中擁有的基因名不同引起了誤差的產生(Durinck.,2005),所以在對人類DEGs與2種獼猴進行比較時,將人類基因提交至Ensembl BioMart與獼猴基因進行一對一直系同源基因的同源篩選。
2.3 差異表達基因的富集分析
使用g:Profiler(Raudvere.,2019)對DEGs做GO功能和KEGG通路富集分析。在Significance threshold項選擇Benjamini-Hochberg FDR,其他選項為默認,并最終選擇FDR<0.05的GO和KEGG富集結果。使用R包ggplot2(Wickham,2011)繪制GO富集結果氣泡圖和KEGG富集結果條形圖。
3 結果
3.1 衰老對TMs和CRs基因表達的影響
341個共同上調和389個共同下調的DEGs與2種獼猴的年齡相關(圖1)。而3個DEGs(、、ENSMMUG00000030508)在TM中下調,在CR中上調。
對2種獼猴年齡相關DEGs分別做了GO和KEGG富集分析。TMs的上調DEGs富集在能量代謝途徑、自噬途徑等相關條目中,如GTPase活性的調控(GO:0036211)、利用自噬機制的過程(GO:0061919)(圖2:左)。而TMs的下調DEGs富集在翻譯和物質合成代謝的相關條目(圖2:右),如翻譯(GO:0006412)、肽生物合成過程(GO:0043043)、肽代謝過程(GO:0006518)。CRs的上調DEGs富集在天然免疫途徑和凝血等相關條目中,如炎癥反應(GO:0006954)、凝血(GO:0007596)等(圖3:左)。CRs的下調DEGs富集在翻譯、體液免疫等相關條目(圖3:右),如基因表達(GO:0010467)、B細胞活化(GO:0042113)。

圖1 普通獼猴和藏酋猴上、下調差異表達基因的Venn圖Fig. 1 Venn diagram of up- and down-regulated differentially expressed genes (DEGs) in Macacca mulatta and M. thibetana
2種獼猴與年齡相關的DEGs富集結果顯示出差異性。在凝血方面,CRs的上調DEGs富集在與凝血相關的生物過程(GO:BP)中,如凝血(GO:0007596),而TMs卻沒有富集到與凝血相關的GO條目,但TMs上調的DEGs富集到了與凝血相關的KEGG條目,如血小板活化(ko04611)。在能量代謝方面,CRs和TMs上調的DEGs都富集到了GTPase相關途徑,如調控GTPase活性(GO:0043087)(圖4:右)。但有所區別的是標注了所有CRs和TMs的DEGs的KEGG通路。CRs只在氧化磷酸化(ko00190)的一個調控節點[F-type ATPase (Eukaryotes) c]表現下調,大部分DEGs隨著年齡增長而上調。與CRs相反的是,大部分TMs的DEGs在氧化磷酸化途徑中隨年齡的增長而下調。
2種獼猴DEGs的富集結果也顯示出許多相似之處。比如,2種獼猴的下調DEGs都富集到了與蛋白質翻譯有關的GO條目,如基因表達(GO:0010467)(圖4:右)。這些富集到與蛋白質翻譯有關的條目中下調的DEGs表達會影響核糖體的整體功能,這與TMs中下調DEGs富集到的核糖體(ko03010)通路及蛋白質合成過程一致。
2種獼猴的上調DEGs均富集到與細胞自噬相關的GO通路,如囊泡介導的轉運(GO:0016192),這與在衰老過程中的細胞代謝減弱所導致的細胞自噬現象一致。此外,CRs上調DEGs富集到的唯一一條KEGG通路是溶酶體(ko04142),與抗衰老密切相關,它同時出現在TMs的上調DEGs富集到的KEGG通路中(圖5)。CRs下調基因富集到 2條KEGG通路,分別是原發性免疫缺陷(ko05340)和B細胞受體信號通路(ko04662),其中B細胞受體信號通路也出現在TMs的下調DEGs富集到的KEGG通路中。
在衰老過程中,2種獼猴都經歷了免疫上的變化,但也有一些差異。CRs的1 205個下調的DEGs富集在體液免疫相關的GO條目中,并且富集到了B細胞受體信號(ko04662)的KEGG通路。在TMs的B細胞受體信號通路中,則是富集了更多的上調DEGs。通過KEGG富集圖(圖6)可以發現, 2種獼猴在B細胞受體信號通路中基因表達水平的巨大分歧。大多數CRs的年齡相關DEGs隨年齡的增長而表達下調,只在5個節點(PIR-β、SOS、PI3K、CaN、IKKγ)表現為上調。而在TMs中,雖然B細胞信號受體通路的起始階段(CD72、Igα、Igβ、CD81、CD19等)的基因表達隨著年齡增長而有所下降,但由于復雜的原因,通路的中間節點(SOS、PKCβ等)和近末尾節點(CaN、ErK等)的基因表達量總體升高。

圖2 藏酋猴的差異表達基因富集到的生物過程GO功能條目(取校正后P值前20)Fig. 2 Top 20 enrichment of biological process-related GO terms of differentially expressed genes (DEGs) in Macacca thibetana
左. 上調DEGs, 右. 下調DEGs
Left. up-regulated DEGs, right. down-regulated DEGs

圖3 普通獼猴的差異表達基因富集到的生物過程GO功能條目(取校正后P值前20)Fig. 3 Top 20 enrichment of biological process-related GO terms of differentially expressed genes (DEGs) in Macacca mulatta
左. 上調DEGs, 右. 下調DEGs
Left. up-regulated DEGs, right. down-regulated DEGs

圖4 普通獼猴和藏酋猴的差異表達基因共同富集到的GO條目(取校正后P值前15)Fig. 4 Top 15 enrichment of GO terms by the common differentially expressed genes (DEGs) in Macacca mulattaand M. thibetana
左. 上調DEGs, 右. 下調DEGs; 灰色為普通獼猴, 黑色為藏酋猴
Left. up-regulated DEGs, right. down-regulated DEGs; gray presents, black represents
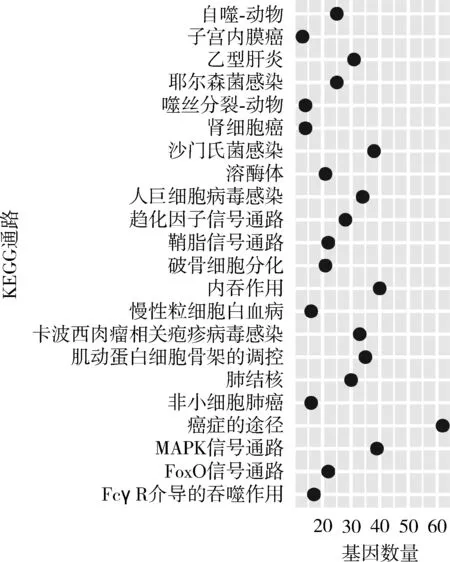
圖5 藏酋猴的上調差異表達基因富集到的KEGG通路Fig. 5 Enrichment of KEGG terms by the up-regulated differentially expressed genes of Macacca thibetana
3.2 TMs、CRs與人類年齡相關的DEGs的比較分析
3個物種的年齡相關DEGs一起比較發現,在1 497個人類年齡相關的DEGs中,有309個同時也與TMs的年齡相關,其中133個上調,150個下調。CRs與人類年齡相關的共有DEGs 310個,其中118個上調,174個下調(表1)。三者共有DEGs 127個,如、等,其中23個上調,38個下調。
衰老過程對保守基因表達的影響在不同物種間存在差異。在TMs中,與人類年齡相關DEGs表達方向相反的DEGs包括16個上調(在人類中下調)和10個下調(在人類中上調)。TMs與人類表達方向相反的26個DEGs富集在Th17細胞的分化(ko04659)中,該通路富集了3個與年齡相關的DEGs(、和),其中和的表達隨年齡增長而上調。起到抑制通路效果的則是隨著年齡增長而下調。在CRs和人類中具有相反趨勢的18個年齡相關DEGs并未富集到功能條目,但這些相反的基因也參與到了代謝和延緩衰老等過程,如凋亡調節因子1()、RNA結合基序蛋白11()、真核翻譯起始因子2亞基3()。

表1 人類、藏酋猴和普通獼猴的差異表達基因交集統計表Table 1 Intersection statistics of differentially expressed genes (DEGs) in human, Macacca thibetana and M. mulatta
3個物種共同的38個下調DEGs富集到了細胞免疫相關GO通路,比如髓系白細胞介導的免疫(GO:0002444)。3個物種共同的23個上調DEGs富集在與體液免疫密切相關的B細胞受體信號通路(ko04662)。為了更全面地了解3個物種衰老過程中的免疫差別,將所有年齡相關DEGs標注在與體液免疫密切相關的B細胞受體信號通路上。在B細胞受體信號通路中,3個物種在起始階段(Igα、Igβ、CD72、CD19)的基因表達隨著年齡增長下調,人類和TMs在中間節點和近末尾節點(人類:VAV、SHIP、Erk等;TMs:SOS、PI3K、PKCβ、BCL-10等)的基因表達總體上調,而CRs則是下調。從整個通路來說,CRs富集了更多下調DEGs。
4 討論
4.1 衰老對2種獼猴新陳代謝和免疫力的不同影響
在衰老過程中,CRs和TMs的基因表達的改變有很多相似的地方。2種獼猴的蛋白質翻譯功能相關基因表達均顯著下調,核糖體的形成過程如核糖體生物發生(GO:0042254)的基因也出現表達下調。這2個過程聯系緊密,并造成一系列連鎖反應。在核糖體的形成過程中,rRNA不足影響核糖體的合成量;核糖體合成量不足又反過來阻礙蛋白質的翻譯過程。翻譯是蛋白質代謝的環節,不足的蛋白質代謝影響嘌呤和嘧啶代謝(Rudolph,1994),從而阻礙DNA合成,進而減緩 mRNA 的表達和蛋白質自身產量,并最終表現為機體的代謝率減慢。

圖6 人類、藏酋猴和普通獼猴年齡相關差異表達基因富集的KEGG比較通路(B細胞受體信號通路)Fig. 6 Comparison of the B cell receptor signaling pathway enriched by the age-related differentially expressed genes (DEGs) in human, Macacca thibetana and M. mulatta
不同的顏色代表DEGs表達方向的不同; 左邊代表人類基因表達水平, 中間代表藏酋猴基因表達水平, 右邊代表普通獼猴基因表達水平, 紅色表示與年齡正相關DEGs的KEGG同源基因單元, 綠色表示與年齡負相關DEGs的KEGG同源基因單元
Different colors represent different expression directions of DEGs; left part represents the gene expression of human, middle part represents the gene expression of, and right part represents the gene expression of; red represents the KOs including positively age-associated DEGs, green represents the KOs including negatively age-associated DEGs
在2種獼猴隨著年齡增長而整體代謝逐漸衰減時,也有部分生物過程被強化,如凝血和自噬相關通路等。個體衰老過程往往伴隨著心血管相關疾病(如動脈粥樣硬化)的發生(Wilkerson & Sane. 2002)。巨噬細胞是動脈粥樣硬化的主要效應細胞(Moore.,2013),2種獼猴的上調DEGs富集到一些巨噬細胞相關的條目:巨噬細胞活化的調控(GO:0043030)、巨噬細胞自噬(GO:0016236,FDR<0.01)。而血栓通常發生在動脈粥樣硬化斑塊的侵蝕或破裂之后,故巨噬細胞的活躍會影響血栓的形成(Prandoni.,2003)。同時血小板活化通路(ko04611)中增強的凝血功能是血栓形成的風險因子之一,這個通路的功能在TMs中隨年齡的增長而增強。推測2種獼猴在衰老過程中血栓的發生率隨年齡的增長而增加,這與本團隊之前在CRs中的研究一致(Yan.,2020)。
在衰老過程中,2種獼猴的天然免疫能力都顯著增強,促進體內形成活躍的炎癥環境(Shaw.,2010)。天然免疫系統通過激烈的細胞自噬、吞噬作用等迅速吞噬老舊細胞、病菌等有害物質,是個體對抗衰老、維護健康的手段(Huynh.,2007)。CRs和TMs免疫能力改變的差異表現在以B細胞為主體的體液免疫上。B細胞受體信號通路會隨著年齡增長而功能減弱,最終導致B細胞合成減少,表現出免疫老化(Niiro & Clark,2002)。KEGG富集圖發現,TMs的B細胞受體信號通路隨著年齡的增長而變得更加活躍,表明體液免疫隨著年齡的增長而增強(Murphy & Weaver,2012),CRs的情況則相反。以上暗示TMs特異性免疫力下降可能沒有CRs明顯,CRs的免疫老化現象隨著年齡的增長可能更加突出(Asquith.,2012)。
2種獼猴在衰老過程中的基因表達總體相似,但也表現出一些差別,如TMs年齡相關的DEGs更多富集在減弱的代謝活動中,而CRs更顯著地表現出免疫老化。
4.2 獼猴與人類在衰老過程中的差異
機體代謝水平和疾病易感性的改變是衰老的重要特征之一(Kenyon,2010;Ruby.,2018a),這些改變也反映在3個物種的血液轉錄組中。3種靈長類動物共同的38個下調DEGs富集到了B細胞受體信號通路(ko04662),該通路通過激活B細胞來啟動下游的體液免疫。在標注了3個物種所有DEGs的B細胞受體信號通路富集圖中,3個物種存在4個相同下調的環節(CD72、CD19、Igα、Igβ)。CD72和CD19是免疫球蛋白基因超家族的成員,Igα和Igβ與表面免疫球蛋白共同構成B細胞抗原受體復合物,CD72、CD19、Igα和Igβ都是B細胞抗原受體表達和功能所必需的分子。衰老對3個物種的共同影響體現在B細胞受體信號通路的起始節點,衰老下調起始物質的表達,從而影響B細胞免疫。另外與人類相比,CRs富集了更多的下調基因,TMs富集了更多的上調基因。由此推斷年齡增長過程中,TMs整體的B細胞免疫可能比人類的更活躍,即擁有更強的體液免疫能力。在衰老過程中,CRs的B細胞數量顯著下降,導致其衰弱的體液免疫(Didier.,2012)。推斷在衰老過程中,體液免疫能力由強到弱依次是TMs、人類、CRs。這種能力的不同可能反映了3個物種免疫系統的差異。但這一推測還需要采用其他研究手段加以驗證。
另一個相似的現象是蛋白質的翻譯功能、核糖體的形成過程,這些過程的相關基因在這3種靈長類動物中都是隨著年齡增長而下調。人類衰老過程中參與蛋白質翻譯的基因顯著下調(Jung.,2015)。蛋白質合成速率降低是衰老過程中的一個重要環節,會導致代謝衰減。但代謝衰減是衰老的原因,還是衰老過程產生的結果尚不明確(Anisimova.,2018)。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
哈哈畫報(2022年4期)2022-04-19 11:11:50
大科技·百科新說(2021年6期)2021-09-12 02:37:27
英語文摘(2021年2期)2021-07-22 07:56:54
好孩子畫報(2020年5期)2020-06-27 14:08:05
意林·全彩Color(2019年6期)2019-07-24 08:13:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
