仙琴蛙對不同類型聲音的感知差異研究
2022-01-26 06:47:16鈕文俊李照沈迪張保衛方光戰安徽大學生命科學學院合肥230601湖南農業大學生物科學技術學院長沙410128中國科學院成都生物研究所成都610041中國科學院大學北京100049
四川動物 2022年1期
關鍵詞:信號
鈕文俊, 李照, 沈迪, 張保衛, 方光戰*(1. 安徽大學生命科學學院,合肥230601; 2. 湖南農業大學生物科學技術學院,長沙410128;3.中國科學院成都生物研究所,成都610041; 4. 中國科學院大學,北京100049)
聽覺是大腦感知外界信息的重要功能之一,通過聽覺系統感知不同的聲音信息,并整合其他模態輸入(如視覺和嗅覺等)的處理結果,最終決定動物捕食、逃逸和交配等行為,對維系個體與種群的生存具有極其重要的意義(Schrode.,2014)。聽覺感知包括對聲音信息的覺察、感覺、注意和知覺等多個認知過程,既往研究對這些過程進行了深入研究(Janik,2000;Charrier.,2001;Vlgnal.,2004;Elie & Theunissen,2018),但是大腦如何對復雜的聲音信號進行解碼和處理依然是神經生物學的核心問題之一。例如,大腦在處理不同類型的復雜聲音信號時,大腦活動的動態特征依然不清楚;對這一問題進行研究有助于深入理解大腦功能、聽覺感知和聽覺目標識別的相關機理。
哺乳類(包括人)、鳥類和爬行類的聽覺行為往往受多模信息影響,視覺、聽覺和嗅覺等信息的時空交互給聽覺感知的研究帶來一定困難。由于大腦功能的保守性,脊椎動物在一定程度上共享著基于對信號的選擇性處理而指導行為響應的基本環路(Krauzlis.,2018),因此可選擇單模通訊動物(比如主要依賴聲音通訊的夜行性蛙類)作為模型對聽覺感知進行研究。對蛙類而言,雄性廣告鳴叫(復雜聲音信號)傳遞著物種、個體、繁殖狀態、所處位置、資源占有、體型等復雜信息,即聲學成分編碼了物種/個體識別所需的信息;雌性根據廣告鳴叫識別物種、亞種或個體,并選擇最適雄性作為配偶。雄性仙琴蛙在繁殖季節能產生2種廣告鳴叫(產生于巢穴內的洞內鳴叫和巢穴外的洞外鳴叫),雌雄兩性均偏好響應洞內鳴叫(Cui.,2012;Fang.,2014);而在非繁殖季節,仙琴蛙通常將自己埋于泥土之中,進入或準備進入冬眠狀態,此時很少聽到外界聲音,尤其不可能聽到同種廣告鳴叫(Yang.,2018)。行為學研究發現,雌雄兩性對同種鳴叫和非同種鳴叫(如白噪聲)的行為響應不同(Cui.,2012;Fang.,2014),而且仙琴蛙的腦網絡特征受聲音信號的生物學意義調制(Xue.,2018),說明仙琴蛙能分辨不同類型的聲音信號。但仙琴蛙在感知不同類型的聲音時其大腦活動的動態特征尚不清楚。
腦電(electroencephalogram,EEG)是神經元群電活動在大腦皮層的總體反映,是腦功能信息傳輸的內部載體,蘊藏豐富、動態的生理信息。EEG功率譜反映了同步放電的神經元數量(Klimesch,1999),即功率譜增加表明參與當前任務的神經元數量增加。通常將EEG劃分為delta、theta、alpha、beta和gamma 5個頻段,各頻段對聽覺認知均具有重要作用(Klimesch.,2007;Klimesch,2012;Wang.,2016),各頻段不同功率譜與特定的大腦狀態相關聯,反映了與注意力、學習、記憶和決策相關的感覺和認知過程(Klimesch,1999,2012;Engel,2001;Thut & Miniussi,2009)。Delta頻段與最古老的信息傳遞系統有關,需要內部專注的任務往往伴隨著delta出現,它抑制干擾任務解決的其他過程(Harmony,2013);其抑制作用可能是由較高的系統抑制較低的系統,即下行抑制機制(Knyazev & Slobodskaya,2003)。Theta頻段在認知過程中起著重要作用,包括感知、注意、學習、空間導航、情景記憶和工作記憶(Kahana,2006;Soltani Zangbar.,2020)。Alpha頻段參與聽覺注意的調制,皮層中alpha頻段振蕩是選擇性注意的關鍵標志(Klimesch,2012)。Beta頻段的活動與刺激加工相關,參與對新奇刺激的處理(Haenschel.,2000)。Gamma頻段在動態選擇信息過程中起重要作用,還與語言的加工和運動協調有關(Uhlhaas.,2011)。但仙琴蛙僅有delta、theta、alpha和beta頻段,缺少gamma頻段,這可能與兩棲動物大腦缺少新皮質有關(Fang.,2012)。由于大腦功能具有保守性,脊椎動物之間的EEG節律沒有本質的系統發育差異(Segura & De Juan,1966),所以EEG是研究聽覺感知的理想信號。
由于處于繁殖期的仙琴蛙能對聲音信號產生相應的行為響應;而處于非繁殖期的仙琴蛙通常埋于泥土之中,很少聽到外界聲音,尤其不可能聽到同種廣告鳴叫。因此,選擇繁殖期的仙琴蛙作為研究對象,更接近于其自然條件下的生存狀態,也更能揭示其自然條件下的聽覺感知特征。為此,本研究以繁殖期的仙琴蛙為對象,采用等概率實驗范式,在隨機播放白噪聲和洞內鳴叫2種不同類型的聲音刺激時,實時采集仙琴蛙相關腦區的EEG信號;分析和探討聽覺感知過程中EEG各頻段的動態特征,相關結果將有助于深入理解蛙類在進行同種鳴聲識別時各EEG頻段的功能。
1 實驗材料和方法
1.1 實驗動物
在繁殖季節,從四川省峨眉山地區采集16只仙琴蛙(雌雄各半)。按性別分裝于2只不透明塑料箱子里(長54 cm×寬40 cm×高33 cm),箱內有適量的泥和水。在室內飼養條件下,雄性往往會發出廣告鳴叫,而雌性通常有卵,保證雌雄兩性在實驗過程中的激素水平維持在一個相對較高的水平。飼養箱置于 12L∶12D 光照周期(08∶00 開燈)的飼養房中,室溫23 ℃±1 ℃。每隔3 d喂食新鮮活蟋蟀一次。手術時,動物平均體質量為9.96 g±1.64 g,平均體長為4.65 cm±0.24 cm。所有實驗操作均遵守中國科學院成都生物研究所動物福利相關規定。
1.2 手術過程
將仙琴蛙浸入0.15%的魚安定(MS-222)溶液中麻醉(保持頭部外露,避免眼部不適),通過夾趾反應判斷麻醉程度,無夾趾反應時停止麻醉。用濕潤紗布包裹動物身體,留出頭部手術區域,依次使用碘伏、75%醫用酒精及消炎藥對手術區域進行處理,避免術后感染。去除手術區皮膚,暴露頭骨,將7個不銹鋼電極(=0.8 mm)植入顱骨內直至接觸硬腦膜為止(通過平均顱骨厚度、不銹鋼螺絲的螺距及電極植入時眼部變化共同確定),其中6個電極分別位于端腦(left telencephalon,LT;right telencephalon,RT)、間腦(left diencephalon,LD;right diencephalon,RD)和中腦(left mesencephalon,LM;right mesencephalon,RM)的左右兩側,參考電極(cerebellum,C)位于小腦上方(圖1)。電極埋植完成后,用牙托水泥對電極進行封固;用自封膜對接插件進行防水密封。動物術后單獨置于裝有適量純凈水的飼養盒中,恢復7 d后開始實驗。實驗結束后,對動物進行安樂死,并在電極相應位點注射蘇木精染料,檢查電極位置是否與預期一致,排除與目標位置不一致的數據。
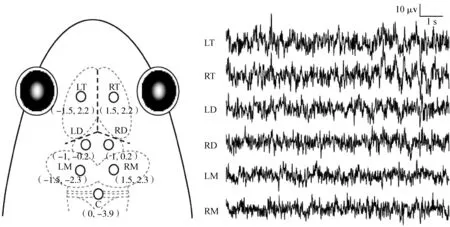
圖1 電極位置分布及相應的10 s EEG特征波形Fig. 1 Electrode placements and 10 s typical EEG tracings for each brain area
3條粗體虛線表示顱骨骨縫; LT、RT、LD、RD、LM、RM分別表示端腦、間腦和中腦的左右兩側; C表示植入小腦上方的參考電極
Three dashed lines in bold denote the intersection among the suture lines in the frog skull; LT, RT, LD, RD, LM, RM denote the left and right sides of telencephalon, diencephalon and mesencephalon, respectively; while C denotes the reference electrode implanted above the cerebellum
1.3 聲音刺激
實驗使用白噪聲和洞內鳴叫2種刺激。雄蛙聽覺敏感的頻率范圍通常比雌性更寬(Liu.,2014),為了更好地檢測聽覺感知的性別差異,需要采用帶寬比同種鳴叫更寬的聲音刺激。由于理想的白噪聲具有無限帶寬(實際使用的白噪聲帶寬與聲音播放設備的頻率響應帶寬相等),同時白噪聲在整個頻域范圍內的所有頻率分量具有相等的功率譜密度,所以白噪聲適合用于對包括神經系統在內的生理系統的刺激響應進行識別和分析(Marmarelis & Marmarelis,1978);并且生理系統對白噪聲的刺激響應在很大程度上不受出現在該系統中的其他類型噪聲影響。為避免偽重復,選用 4只非實驗對象的同種雄性個體的、包含 5個音節的洞內廣告鳴叫,每個洞內鳴叫分別與白噪聲配對,組成4組刺激對(圖2)。白噪聲上升/下降沿均為7.5 ms(正弦調制),時長等于 4個洞內鳴叫的平均時長(約1.28 s)。隨機選取1組刺激對用于隨機選取的 4只仙琴蛙(雌雄各半)。聲音刺激通過2個便攜式揚聲器(SME-AFS;Saul Mineroff Electronics,Elmont,New York,USA)播放,揚聲器置于實驗箱兩端,并與實驗箱等距。聲音強度通過聲壓計(AWA6291,愛華,杭州)調整為65 dB±0.5 dB SPL(在實驗箱中心測得,C計權)。
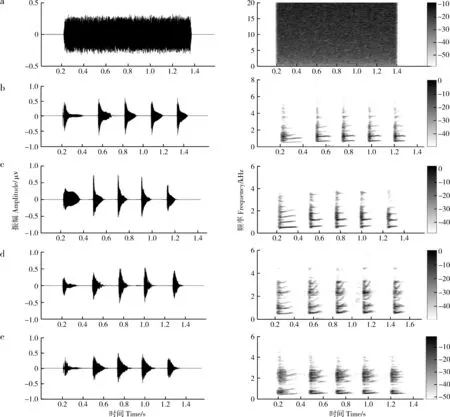
圖2 聲音刺激的波形圖和頻譜圖 Fig. 2 Waveforms and spectrograms of the five stimuli
a. 白噪聲, b~e. 來自4只同種個體的洞內鳴叫
a. white noise, b-e. 4 advertisement calls acquired from four conspecific individuals
1.4 數據采集
實驗在隔音的電磁屏蔽室(背景噪聲23.0 dB±1.7 dB)進行。恢復7 d后,動物被放入裝有泥和水的不透明實驗箱(長80 cm×寬60 cm×高55 cm),并連接至信號采集系統(RM6280C,成儀,成都),適應24 h;實驗箱上方裝有紅外攝像機,監測被試行為。采用隨機、等概率方式回放白噪聲與洞內鳴叫,刺激呈現順序隨機(即下一個刺激是什么不可預測)、刺激間隔在2 s、2.5 s、3 s、3.5 s和4 s中隨機(下一個刺激什么時候出現不可預測;由于在自然條件下雄性廣告鳴叫之間的平均間隔為 3.3 s,且雄性對時間間隔為3 s的聲音回放的預測準確率最高(Fang.,2014),所以選取以3 s為中心的5個時長為刺激間隔),最大化消除有意注意影響,實驗共隨機回放了40次聲音刺激(白噪聲回放 20次,洞內鳴叫回放20次)。刺激播放起始時刻,播放程序向信號采集系統發送TTL電平用于數據同步。采集動物的EEG信號和行為數據,帶通濾波設置為0.05~200 Hz,采樣頻率為1 000 Hz。實驗前一天喂食,實驗過程中不進食,以保證信號受營養代謝的影響最小。
1.5 數據處理
EEG信號經過50 Hz陷波和0.5~45 Hz帶通濾波后,分別取刺激起始點前后1 s且無偽跡的原始數據進行分析。利用Welch算法(漢明窗,頻譜分辨率為0.5 Hz)計算每段數據的delta(0.5~5.5 Hz)、theta(5.5~8.5 Hz)、alpha(8.5~17 Hz)、beta(17~45 Hz)的絕對功率譜(Fang.,2012)。得到每個EEG頻段、每種刺激(白噪聲和洞內鳴叫)、每個腦區(左右端腦、左右間腦和左右中腦)在不同時間(刺激前和刺激后)的絕對功率譜。
1.6 統計分析
對絕對功率譜進行正態分布檢驗(Shapiro-Wilk檢驗)和方差同質檢驗(Levene’s檢驗)。利用5個因素(刺激組別、性別、刺激類型、時間和腦區)重復測量ANOVA對數據進行分析,發現在刺激組別和性別這2個因素上均無顯著差異,說明本實驗已排除偽重復可能造成的不利影響。進一步采用三因素(刺激類型、時間和腦區)重復測量ANOVA。若交互效應顯著,進行簡單或簡單-簡單效應分析。事后檢驗采用Bonferroni檢驗;若有必要,進行Greenhouse-Geisser矯正;分別用Partial和Cohen’s d檢測方差分析和檢驗的效應度(0.20為低效應度,0.50為中效應度,0.80為高效應度)。統計分析使用SPSS 26。顯著水平定義為α=0.05。
2 結果
2.1 Delta頻段
刺激類型(=13.140,partial=0.467,=0.002)、時間(=43.385,partial=0.752,<0.0001)和腦區(=11.590,partial=0.436,=0.001)的主效應均顯著;且三交互效應顯著(=3.061,partial=0.169,=0.039;表1)。簡單-簡單效應分析和簡單效應分析結果顯示,對不同刺激而言:左右間腦和左右中腦在白噪聲刺激下的絕對功率譜均顯著小于洞內鳴叫刺激;對刺激前后而言:白噪聲和洞內鳴叫均表現出刺激前的絕對功率譜顯著小于刺激后;對不同腦區而言:腦區間的差異主要表現為端腦最大、間腦次之、中腦最小,但左右端腦、左右間腦、左右中腦之間均無顯著差異(表2;圖3)。
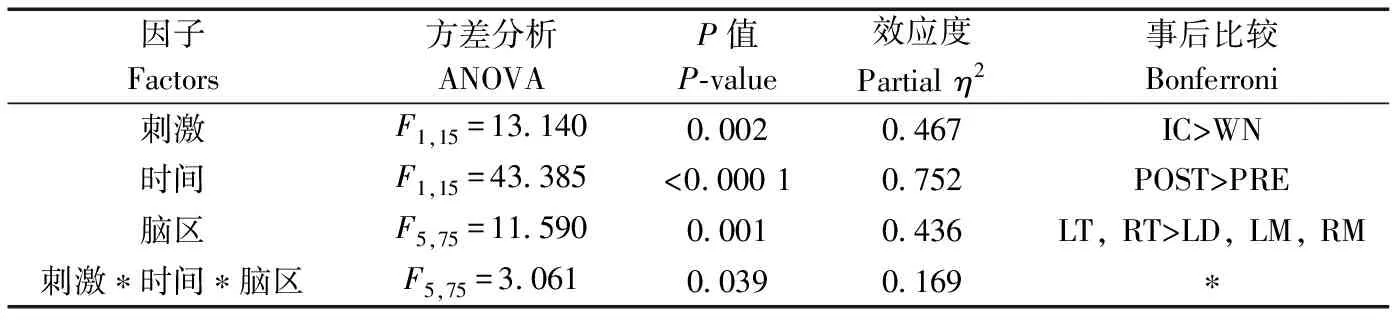
表1 Delta頻段絕對功率譜的ANOVA統計結果Table 1 Results of ANOVA for absolute power spectra of delta band
注: “>”表示左側條件下的絕對功率譜大于右側, 同側間無顯著差異; WN. 白噪聲, IC. 洞內鳴叫, PRE. 刺激前, POST. 刺激后, LT, RT. 左右端腦, LD, RD. 左右間腦, LM, RM. 左右中腦, NA. 不適用; * 結果詳見后表; 下同
Notes: the symbol “>” denotes that the absolute power spectra of the four bands associated with various conditions on the left side of “ > ” are significantly larger than those on the right side, and no significant difference exists among the corresponding conditions on the same side of “ > ” for each case; WN. white noise, IC. inside call, PRE. pre stimulation, POST. post stimulation, LT, RT. the left and right telencephalon, LD, RD. the left and right diencephalon, LM, RM. the left and right mesencephalon, NA. not applicable; * see the table below; the same below

表2 Delta頻段絕對功率譜的簡單-簡單效應分析結果Table 2 Results of simple-simple effects analysis for the absolute power spectra of delta band
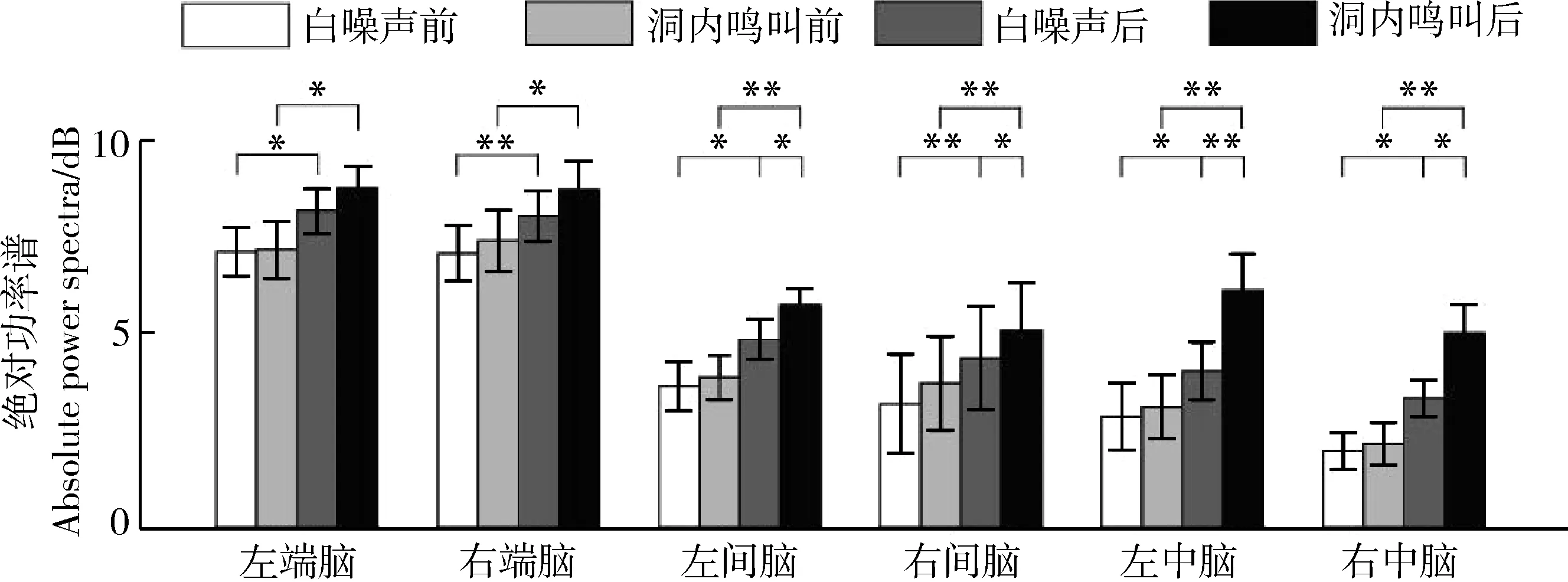
圖3 白噪聲和洞內鳴叫刺激下不同腦區、不同時間(刺激前后)對應的delta頻段的絕對功率譜Fig. 3 Absolute power spectra of delta band elicited in different brain regions before and after playbacks of white noise and inside call
*<0.05, **<0.001; 下同, the same below
2.2 Theta頻段
時間(=98.923,partial=0.868,<0.000 1)和腦區(=12.987,partial=0.464,<0.000 1)的主效應均顯著;且三交互效應顯著(=5.267,partial=0.260,=0.006;表3)。簡單-簡單效應分析和簡單效應分析結果顯示,對不同刺激而言:左中腦和右中腦在白噪聲刺激下的絕對功率譜均顯著小于洞內鳴叫;對刺激前后而言:白噪聲和洞內鳴叫刺激前的絕對功率譜均顯著小于刺激后;對不同腦區而言:腦區之間的差異主要表現為端腦最大、間腦次之、中腦最小;但左右端腦、左右間腦、左右中腦之間均無顯著差異(表4;圖4)。

表3 Theta頻段絕對功率譜的ANOVA統計結果Table 3 Results of ANOVA for absolute power spectra of theta band
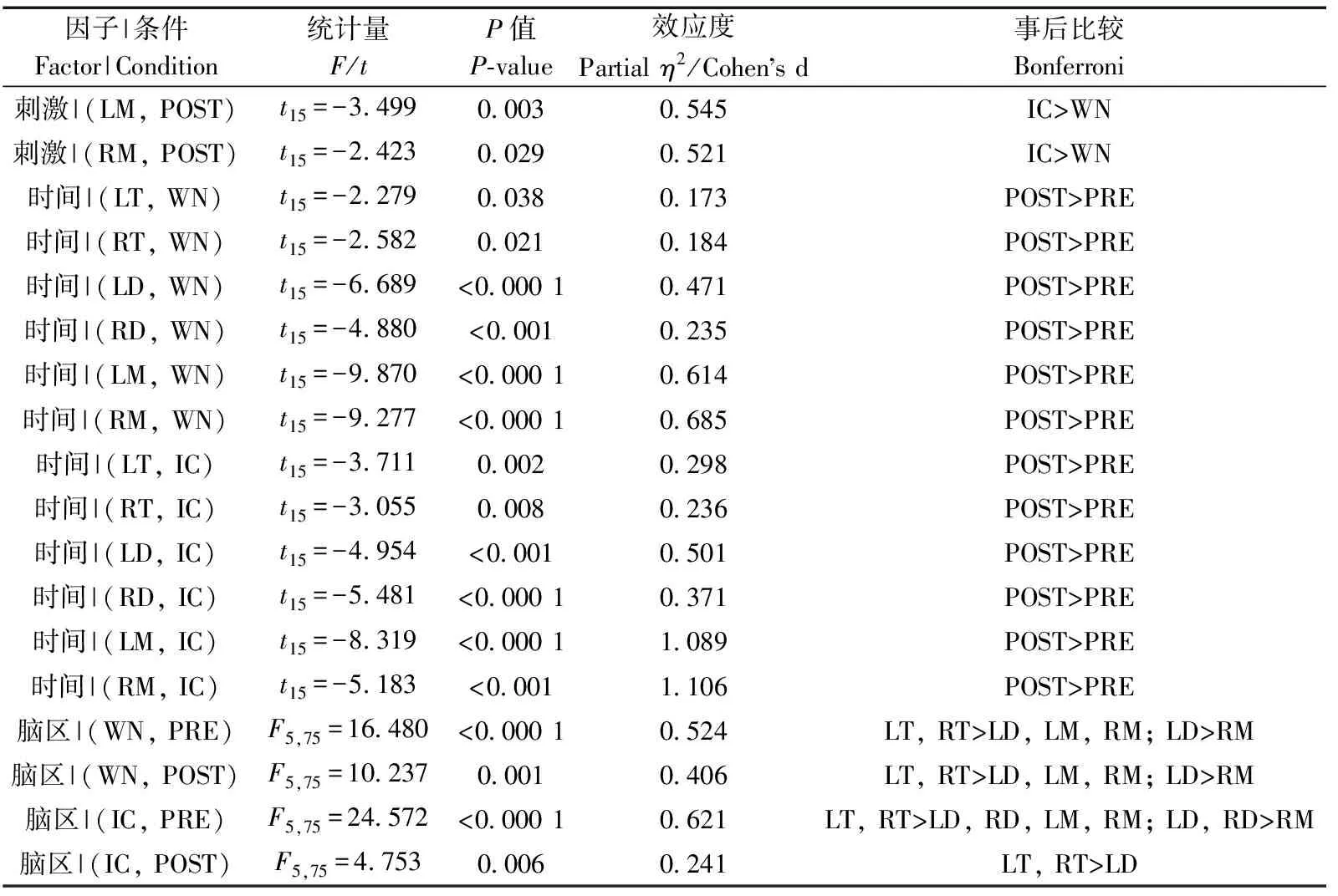
表4 Theta頻段絕對功率譜的簡單-簡單效應分析結果Table 4 Results of simple-simple effects analysis for the absolute power spectra of theta band
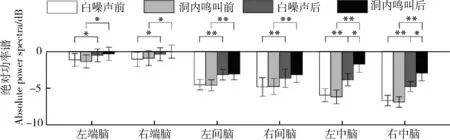
圖4 白噪聲和洞內鳴叫刺激下不同腦區、不同時間(刺激前后)對應的theta頻段的絕對功率譜Fig. 4 Absolute power spectra of theta band elicited in different brain regions before and after playbacks of white noise and inside call
2.3 Alpha頻段
刺激類型(=8.751,partial=0.368,=0.010)、時間(=36.185,partial=0.707,<0.000 1)和腦區(=13.863,partial=0.480,<0.001)的主效應均顯著;且三交互效應顯著(=5.016,partial=0.251,=0.011;表5)。簡單-簡單效應分析和簡單效應分析結果顯示,對不同刺激而言:左端腦、左中腦和右中腦在白噪聲刺激下的絕對功率譜均顯著小于洞內鳴叫;對刺激前后而言:絕大部分腦區在白噪聲和洞內鳴叫刺激前的絕對功率譜顯著小于刺激后;對不同腦區而言:整體而言,端腦最大、間腦次之、中腦最小;但左右端腦、左右間腦、左右中腦之間均無顯著差異(表6;圖5)。
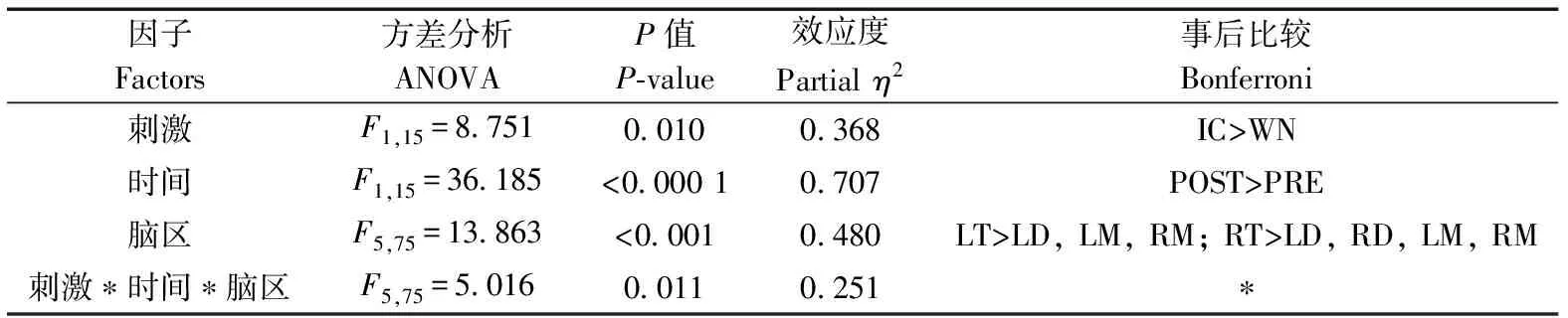
表5 Alpha頻段絕對功率譜的ANOVA統計結果Table 5 Results of ANOVA for absolute power spectra of alpha band
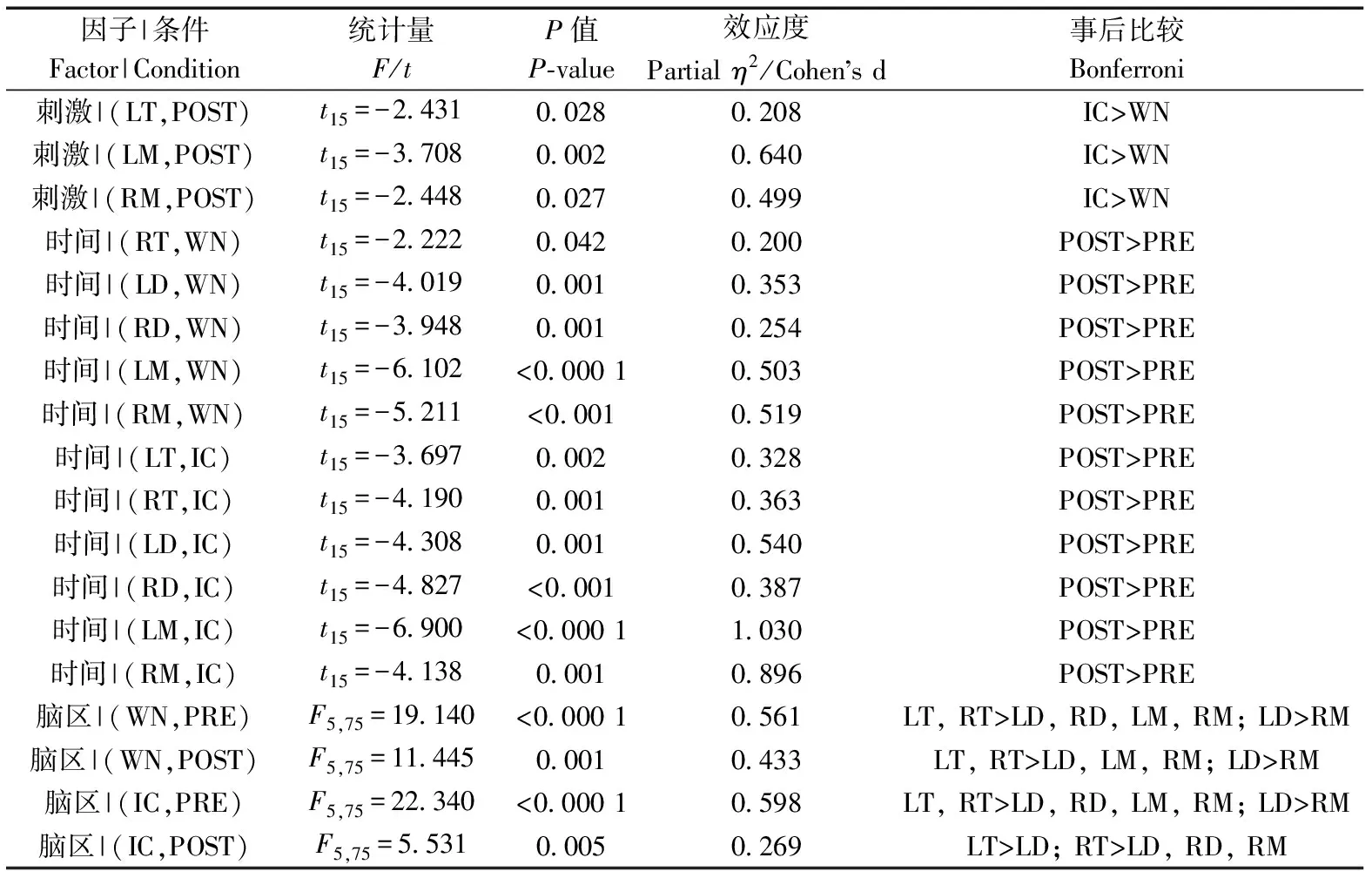
表6 Alpha頻段絕對功率譜的簡單-簡單效應分析結果Table 6 Results of simple-simple effects analysis for the absolute power spectra of alpha band
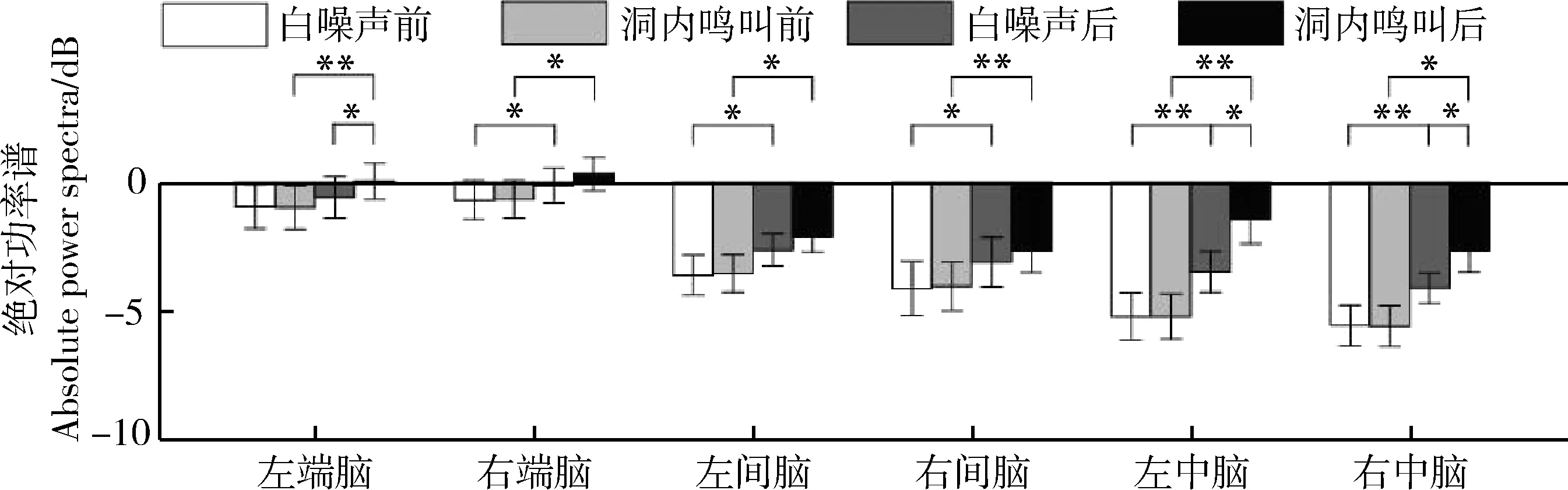
圖5 白噪聲和洞內鳴叫刺激下不同腦區、不同時間(刺激前后)對應的alpha頻段的絕對功率譜Fig. 5 Absolute power spectra of alpha band elicited in different brain regions before and after playbacks of white noise and inside call
2.4 Beta頻段
刺激類型(=5.761,partial=0.277,=0.030)、時間(=18.979,partial=0.559,=0.001)和腦區(=16.324,partial=0.521,<0.000 1)的主效應均顯著;刺激與時間的交互作用顯著(=8.632,partial=0.365,=0.010)、時間與腦區的交互作用顯著(=8.305, partial=0.356,=0.001;表7)。簡單效應分析結果顯示,對不同刺激而言:白噪聲刺激后的絕對功率譜顯著小于洞內鳴叫;對刺激前后而言:白噪聲和洞內鳴叫刺激前的絕對功率譜顯著小于刺激后;對不同腦區而言:腦區之間的差異具體表現為端腦最大、間腦次之、中腦最小;但左右端腦、左右間腦、左右中腦之間均無顯著差異(表8;圖6)。
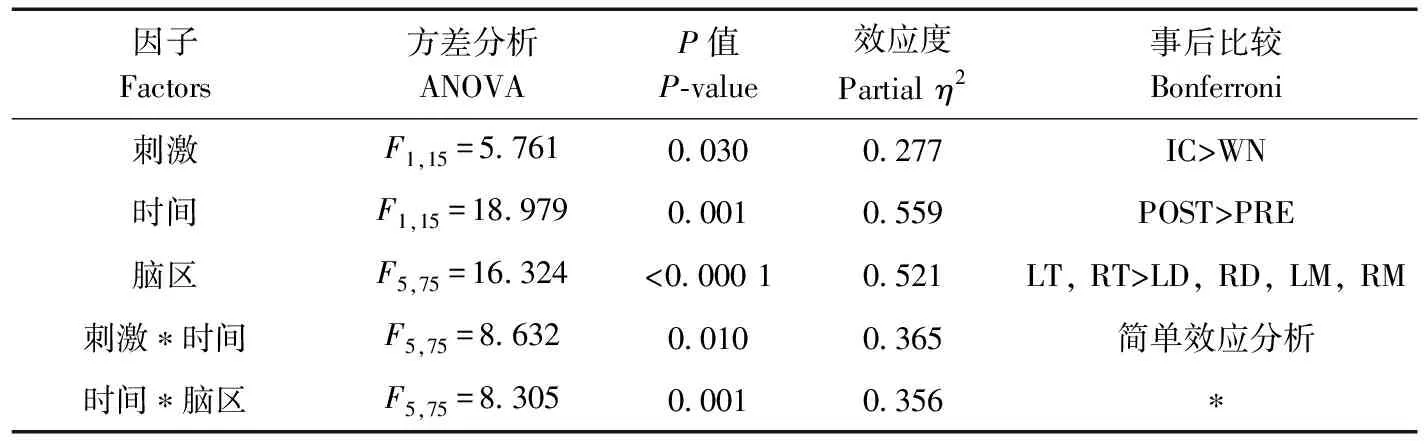
表7 Beta頻段絕對功率譜的ANOVA統計結果Table 7 Results of ANOVA for absolute power spectra of beta band

表8 Beta頻段絕對功率譜的簡單效應分析結果Table 8 Results of simple effects analysis for the absolute power spectra of beta band

圖6 白噪聲和洞內鳴叫刺激下不同腦區、不同時間(刺激前后)對應的beta頻段的絕對功率譜Fig. 6 Absolute power spectra of beta band elicited in different brain regions before and after playbacks of white noise and inside call
3 討論
本研究發現,在播放白噪聲和洞內鳴叫時,仙琴蛙EEG各頻段活動均表現出刺激后大于刺激前,說明各頻段共同參與了仙琴蛙的聽覺感知過程;不同腦區對應的各頻段活動模式主要表現為端腦最大、間腦次之、中腦最小,說明仙琴蛙聽覺感知需全腦共同參與;洞內鳴叫刺激下EEG各頻段的絕對功率譜強于白噪聲刺激,說明EEG各頻段活動強度受到聲音刺激的生物學意義調制。
3.1 聲音刺激使EEG各頻段的活動增加
注意、知覺、學習和記憶等認知過程的基礎是EEG振蕩,這些認知過程并非相互獨立,而是緊密相關的(Baar,2005)。因此,討論各頻段的認知功能對于闡明聽覺感知的神經機制具有重要意義。本研究發現仙琴蛙EEG各頻段在給予聲音刺激后均出現絕對功率譜增大的現象,這與EEG各頻段的認知功能密切相關。
Delta頻段參與注意和刺激顯著性檢測等認知過程(Knyazev,2007;Knyazev.,2009;Güntekin & Basar,2016),在低等脊椎動物的大腦活動中占主導地位(Knyazev & Slobodskaya,2003)。本研究發現聲音刺激引起delta頻段活動增加,說明其參與了仙琴蛙的聽覺感知過程。蛙類通過工作記憶保持對聲音信號的注意(Akre & Ryan,2010),而記憶信息存儲在分布式theta網絡之中,并與大腦后部區域的傳入感覺痕跡相匹配(Sauseng.,2008)。同時,theta頻段與多種感覺共享注意和注意分配,如視聽整合,這也是theta頻段具有認知控制功能的原因之一(Wang.,2016;Keller.,2017)。聲音刺激引起仙琴蛙theta頻段活動增加,說明仙琴蛙可能與其他蛙類一樣,運用工作記憶保持對聲音信號的注意(Akre & Ryan,2010)。傳統認為alpha頻段表征大腦的空閑態,但最近研究發現,alpha頻段與注意的2個基本功能(抑制和選擇)密切相關,前者通過事件相關同步實現,后者通過事件相關去同步解除抑制實現(Klimesch.,2007;Klimesch,2012);即alpha頻段表征大腦通過抑制與當前任務不相關的認知過程,提高大腦相關認知任務的信號處理的皮層信噪比。本研究發現刺激后alpha功率增大可能也體現了alpha的抑制作用。Beta頻段活動與刺激加工(Siniscalchi.,2013)和注意調節相關(Wrobel.,2007),本研究發現beta頻段活動增加,體現其頻段參與仙琴蛙的聽覺感知過程。
綜上可知,聲音刺激引起的EEG各頻段的絕對功率譜增大(即相關頻段的活動增強),與刺激呈現時仙琴蛙需要調用注意等認知資源處理相關信息匹配;同時與“同一頻段可能參與多種大腦功能的實現,同一大腦功能亦需多個頻段的共同參與”的結論一致(Sauseng & Klimesch,2008;Kim & Davis,2021)。
3.2 EEG各頻段的活動強度受聲音信號的生物學意義調制
洞內鳴叫刺激下的端腦、間腦、中腦的絕對功率譜比白噪聲刺激更大,反映了處于繁殖期的仙琴蛙在處理不同類型的聲音信息時會投入不同的認知資源。在繁殖季節,雄性仙琴蛙通常發出廣告鳴叫與同種其他雄性個體競爭并吸引雌性,雌性則根據雄性發出的廣告鳴叫,選擇最適雄性作為配偶(Cui.,2012),因此廣告鳴叫對雌雄兩性的成功繁殖至關重要,這類繁殖相關信息需投入更多的認知處理資源;而白噪聲在自然界中并不存在,不包含特定的生物學信息,仙琴蛙投入的認知資源就會相對較少。本研究結果亦說明仙琴蛙能辨別 2類聲音,且EEG各頻段活動強度受聲音信號的生物學意義調制,這與Xia等(2018)發現的仙琴蛙腦網絡受聲音信號的生物學意義調制的結論一致。無尾類的中腦存在響應同種鳴叫中特定聲學特征的特異性神經元(Feng.,1990;Gooler & Feng,1992;Edwards.,2008)和可能與調節聽覺處理有關的類固醇受體(Forlano.,2010),而雌二醇可增強對聲音的聽覺反應(Yovanof & Feng,1983)及同種鳴叫和其他聲音之間的聽覺差異性(Chakraborty & Burmeister,2015)。本研究采用繁殖期的仙琴蛙,其激素水平相對穩定,中腦特異性神經元的存在可解釋其繁殖期如何對繁殖信息投入更多的認知資源。本研究并未發現大腦活動的性別差異,說明在電生理層面,雌雄兩性對不同類型聲音(同種鳴叫和白噪聲)的處理過程存在相似性。
本研究發現EEG各頻段的絕對功率譜由端腦到間腦再到中腦依次減小;這可能與聽覺感知或電極布局相關。一方面,端腦、間腦和中腦共同參與了無尾類的聽覺感知。比如損毀紋狀體(位于端腦)和丘腦的淺層或深層結構(位于間腦)損害無尾類的鳴叫識別,而中腦(包含半環隆枕)是無尾類聽覺感知的中樞,也是無尾類聽覺系統中首個基于聲音信號的時頻域信息進行復雜特征檢測的腦區(Narins.,2006);聲音信號中的“what”和“where”信息在半環隆枕中整合(Fang.,2015),損毀半環隆枕嚴重影響無尾類的趨聲性行為,說明聽覺感覺階段(比如簡單刺激特征的識別和聲源定位)在中腦水平就基本完成(Wilczynski & Endepols,2006);但對復雜聲音信號的處理(如同種鳴叫的識別與感知)需端腦參與(Yang.,2018)。本研究發現EEG各頻段的絕對功率譜表現為端腦最大、間腦次之、中腦最小,這可能在一定程度上也反映了仙琴蛙的聽覺感知需要全腦共同參與。另一方面,當前結果可能與電極布局有關,本研究的參考電極位于小腦上方,端腦電極離參考電極最遠,中腦電極離參考電極最近;由于EEG信號隨距離增加而衰減,因此對差分放大器而言,相同的信號在端腦采集會比在間腦和中腦采集時更大(由于電極數較少,不便于用平均參考或重參考)。同時,本研究通過分析仙琴蛙EEG各頻段絕對功率譜的變化,探討各頻段在仙琴蛙聽覺感知過程中的功能和大腦活動在不同類型聲音之間的差異,但由于研究方法和技術條件的限制,未能揭示各頻段在聽覺感知過程中的確切功能及其對辨別不同類型聲音的具體作用,未來可以使用空間密度更大的電極采集聽覺感知相關核團的局部場電位信號或采用事件相關電位中的Oddball實驗范式,并結合行為學進一步探討仙琴蛙聽覺感知過程中EEG各頻段的活動特征與功能。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
