嫁接于3個棗園病砧的抗病接穗棗瘋植原體檢測及分子變異*
2022-01-18 00:20:08張文鑫于少帥田國忠任爭光王圣潔孔德治林彩麗
林業科學 2021年11期
張文鑫 于少帥 田國忠 王 合 任爭光 王圣潔 孔德治 李 永 林彩麗
(1.中國林業科學研究院森林生態環境與保護研究所 國家林業和草原局森林保護學重點實驗室 北京 100091; 2.中國熱帶農業科學院椰子研究所 文昌 571339;3.北京市林業保護站 北京 100029;4.北京農學院植物科學技術學院 北京 102206)
由植原體(phytoplasma)引起的棗瘋病(jujube witches’-broom,JWB)是威脅我國棗樹(Ziziphusjujuba)栽培和紅棗生產的最嚴重病害之一。該病害傳染性強、分布范圍廣、危害嚴重;病樹結棗少、品質下降或不結棗,小樹1~2年、大樹5~6年即全株死亡,造成巨大經濟損失(田國忠,2002)。棗瘋病的防治措施主要有選用無病繁殖材料、推廣抗病或耐病品種、清除病株和野生酸棗毒源、殺滅或趨避傳病媒介、病樹抗生素治療及水肥管理等栽培和生態控制等(王焯等,1988;田硯亭等,1993;田國忠等,2009)。
我國是棗樹原產地,栽培歷史悠久,種質資源豐富,其中一些品系表現出對棗瘋病的高度感病,而另一些品系則表現出不同程度的自然抗病能力(曲澤洲等,1991;田國忠等,1999)。有學者通過人工接種-嫁接手段鑒定和篩選出抗病品系(種)(溫秀軍等,2005;趙錦等,2006;趙京芬等,2011)。任爭光等(2015)采用分子檢測手段對抗病與感病品系染病后病原菌濃度的比較研究初步揭示了某些抗病品系對病原菌生長與繁殖的抑制作用。病原菌因環境和寄主的選擇壓力作用存在廣泛的遺傳變異與多樣性,因此,充分了解不同地區病原菌變異不僅有助于預測病害的發生和流行規律,而且及時監測病原菌的致病性變異對合理利用抗病遺傳資源、預防病害爆發性流行、控制棗瘋病等均具有重要意義(馬占鴻,2010;于少帥等,2016)。
植原體難于人工培養,從技術上很難獲得單克隆菌株,因此病菌變異分析的難度較大。王海妮等(2002)利用16SrDNA和tuf基因比較發現,陜西、河北、山東的棗瘋病病原一致。Bu等(2016)對我國北方4省棗瘋植原體株系16S rDNA進行克隆測序,測得其同源性超過99.8%,推測4省棗瘋植原體廣泛傳播與其高度保守性特征相關。徐啟聰等(2009)以16S rDNA、16S-23S間區序列(SR)和secY基因片段為靶標研究我國不同地區棗瘋病分離物的分子變異,發現不同地區、不同品種上的棗瘋植原體間存在較為豐富的遺傳變異。目前,對于寄主抗病反應、癥狀表現、環境適合度相關的植原體群體的生長繁殖規律、分子變異特性等仍不明確。本研究在對我國主要棗產區抗病資源調查與鑒定的基礎上,選擇有代表性的抗病品系在3個不同背景已感染棗瘋病的棗園進行嫁接傳病試驗,采集三地不同品種、不同抗性和不同病級棗樹接穗和砧木樣品作為試材,采用直接PCR、巢式PCR和環介導等溫擴增(LAMP)技術對嫁接接種后的砧木和接穗感染植原體狀況進行檢測;同時,利用16S rDNA、SR、secA基因、tuf基因、fusA-tuf基因間區、himA及其下游假定蛋白基因序列等基因片段進行分析,揭示病原遺傳變異規律和系統發育關系,以期為抗病品系合理利用與病害防治提供依據。
1 材料與方法
1.1 供試樣品
嫁接傳病試驗地為北京玉泉山棗樹資源圃(YQ)、北京昌平王家園(CP)和河北唐縣軍城鎮(JC)3個棗園。選取棗園內自然發病IV-V級的棗樹作為砧木,當年3—4月采集不同品種的健康接穗,于當年4月底采用插皮接方式嫁接到上述發病砧木上。昌平王家園嫁接用發病砧木品種為冬棗(Z.jujuba‘Dongzao’);河北唐縣軍城鎮棗園發病砧木品種為當地感病婆棗(Z.jujuba‘Pozao’);玉泉山有9株嫁接樹,其中2株棗樹砧木嫁接多接穗,苗圃地T19砧木上嫁接4種接穗,分別為T27(2016和2017年都有成活嫁接枝)、T30、Z18、50號;中國林業科學研究院(CAF)內1株未知品系砧木(2009年嫁接時砧木無癥,2016年發病)嫁接3種接穗,分別為T27、M11、JK。同時在玉泉山苗圃用健康接穗嫁接到健康砧木上作為對照處理。接穗及砧木抗性情況見表1。

表1 不同棗樹品種對棗瘋病抗性情況Tab.1 The resistance information of different jujube cultivars to jujube witches’broom
2017年8—10月,采集北京玉泉山棗樹資源圃、北京昌平王家園和河北唐縣軍城鎮棗園三地棗樹材料樣品57份。同時選取國內幾個地區棗瘋病樣品9份,分別為江蘇泗洪大棗4份(JWB-JSSH-QC1、JWB-JSSH-QC2、JWB-JSSH-QC3、JWB-JSSH-FJT)、山東泰安JWB-SDTA、河北唐縣軍城鎮梁家溝JWB-HBTX-LJG、河北唐縣軍城鎮史家溝JWB-HBTX-SJG、山東泰安長寧縣葛石鎮東長紅棗(Z.jujuba‘Changhongzao’)JWB-SDTA-GSZD、北京通州區瑞正園農莊JWB-BJTZ-RZY。
1.2 試驗方法
1.2.1 樣品采集與DNA提取 采集3個棗園不同材料中有典型棗瘋病癥的棗樹組織,無癥樣品選取1年生枝條梢部棗吊葉片,同時進行綜合病級劃分(I-V級)(任爭光等,2015),單純黃葉癥狀分級方法為i~iii,其中葉緣或葉面斑駁輕微發黃褪綠為黃葉i級,ii級為葉面2/3發黃或僅主葉脈未褪綠,iii級黃葉為全葉發黃變脆。將采集的棗樹樣品分別裝袋并標記,干燥備用。植物總DNA提取參照艾德萊生物科技有限公司CTAB法植物基因組DNA快速提取試劑盒說明書。
1.2.2 植原體分子檢測 利用通用引物對P1/P7(Dickinsonetal.,2013)和R16 mF2/R16 mR1(Leeetal.,1993)對棗樹樣品DNA進行PCR和Nested-PCR。LAMP檢測參考廣州迪澳生物科技有限公司恒溫擴增試劑盒的說明書。采用顯色法對提取的總DNA進行63 ℃水浴恒溫擴增(王圣潔等,2017)。
1.2.3 棗瘋植原體基因與基因間區的擴增測序與序列分析 將棗樹DNA樣品進行16S rDNA、SR(Changetal.,2004)、secA基因(Hodgettsetal.,2008)、tuf基因(Dickinsonetal.,2013)、himA及其下游假定蛋白基因(設計引物對himA-F 5′-TAGAAATACCTGCGAAAACC-3′/himA-R 5′-AAGGA AGATTGTTCATTAATTG-3′擴增,退火溫度54 ℃)、fusA-tuf基因間區(于少帥等,2016)PCR擴增。將具有陽性結果的PCR產物送北京博邁德基因技術有限公司測序。利用DNAMAN6.0對測序結果進行分析和手工校驗,用DNAman6.0和MEGA5.0對所得序列進行校驗、拼接、比對和分子進化遺傳分析。
1.2.4 構建系統發育樹 將3個棗園不同樣品各基因的測序結果分別與GenBank中已報道的植原體不同組的序列構建系統發育樹,建樹采用NJ法,Bootstrap 值為1 000。
2 結果與分析
2.1 3個棗園棗樹嫁接傳毒癥狀表現與樣品的直接PCR、巢式PCR和LAMP檢測
將抗性程度不同品系的休眠接穗嫁接到發病嚴重的感病砧木(一般為IV-V級)上,接穗上的萌芽表現出的癥狀包括小葉、叢枝、花梗延長、花變葉、脈間褪綠、葉片黃化、褐化、卷葉等,其中感病的砧木,包括昌平棗園的冬棗、唐縣軍城的婆棗、玉泉山棗資源圃的泗洪大棗、襄汾圓棗、晉棗、酸棗等發病砧木及感病接穗濮陽冬棗主要表現為叢枝、小葉、花變葉等典型癥狀。抗病品系接穗T30和T27在嫁接早期常出現花梗延長現象,但多未進一步發展成花變葉,而大都能正常開花結果;在正常光照生長條件下二者也未產生叢枝癥狀,T27多表現出典型的葉片黃化癥狀,T30則很少產生葉片黃化癥狀;但在遮蔭或光照不足的條件下,這2個品系皆產生了小葉叢枝癥狀(表2)。
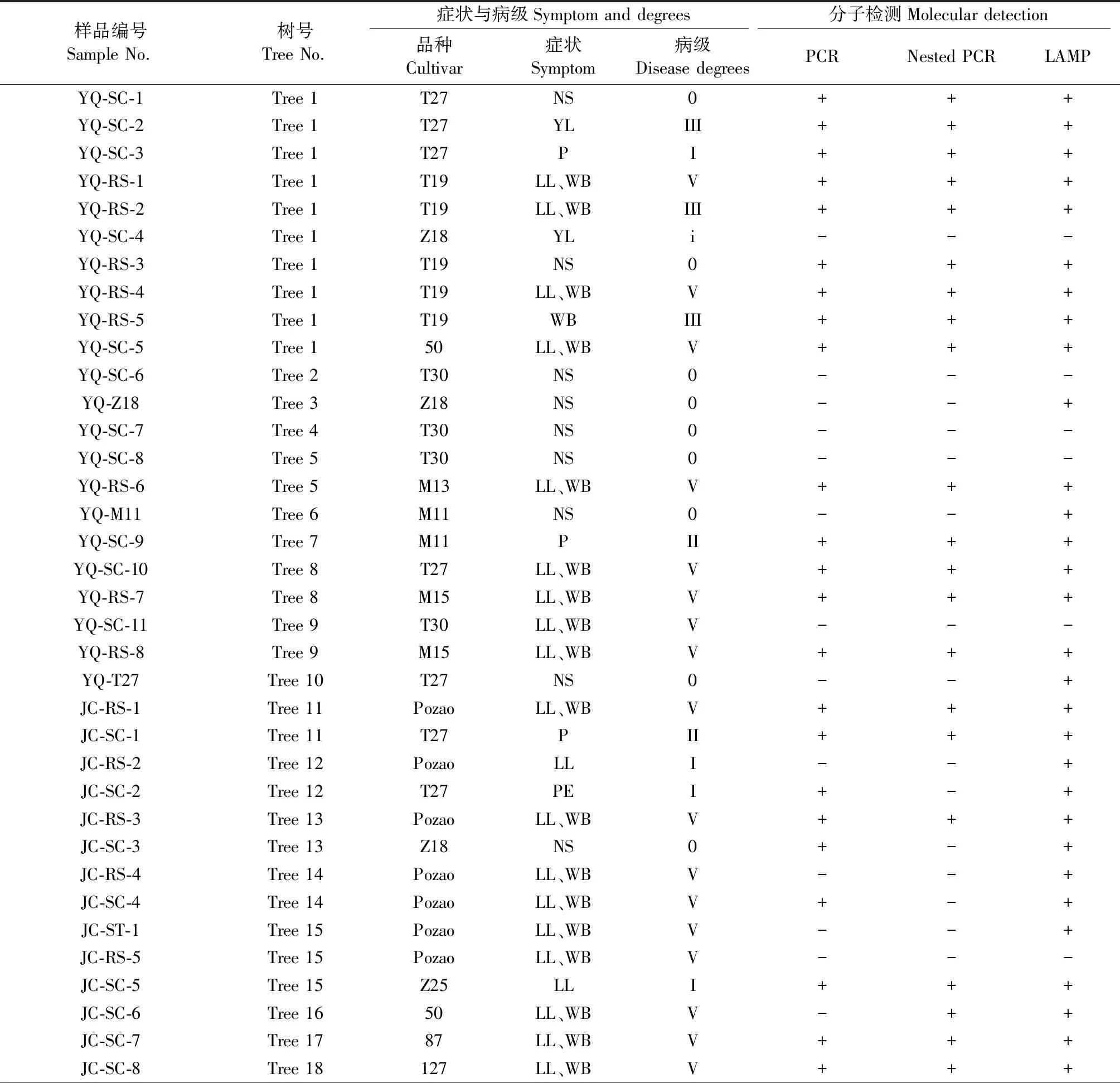
表2 3個棗園不同品種接穗嫁接后癥狀表現及植原體檢測結果①Tab.2 Symptom and phytoplasma detection results of different cultivars after grafting transmission in three jujube orchards
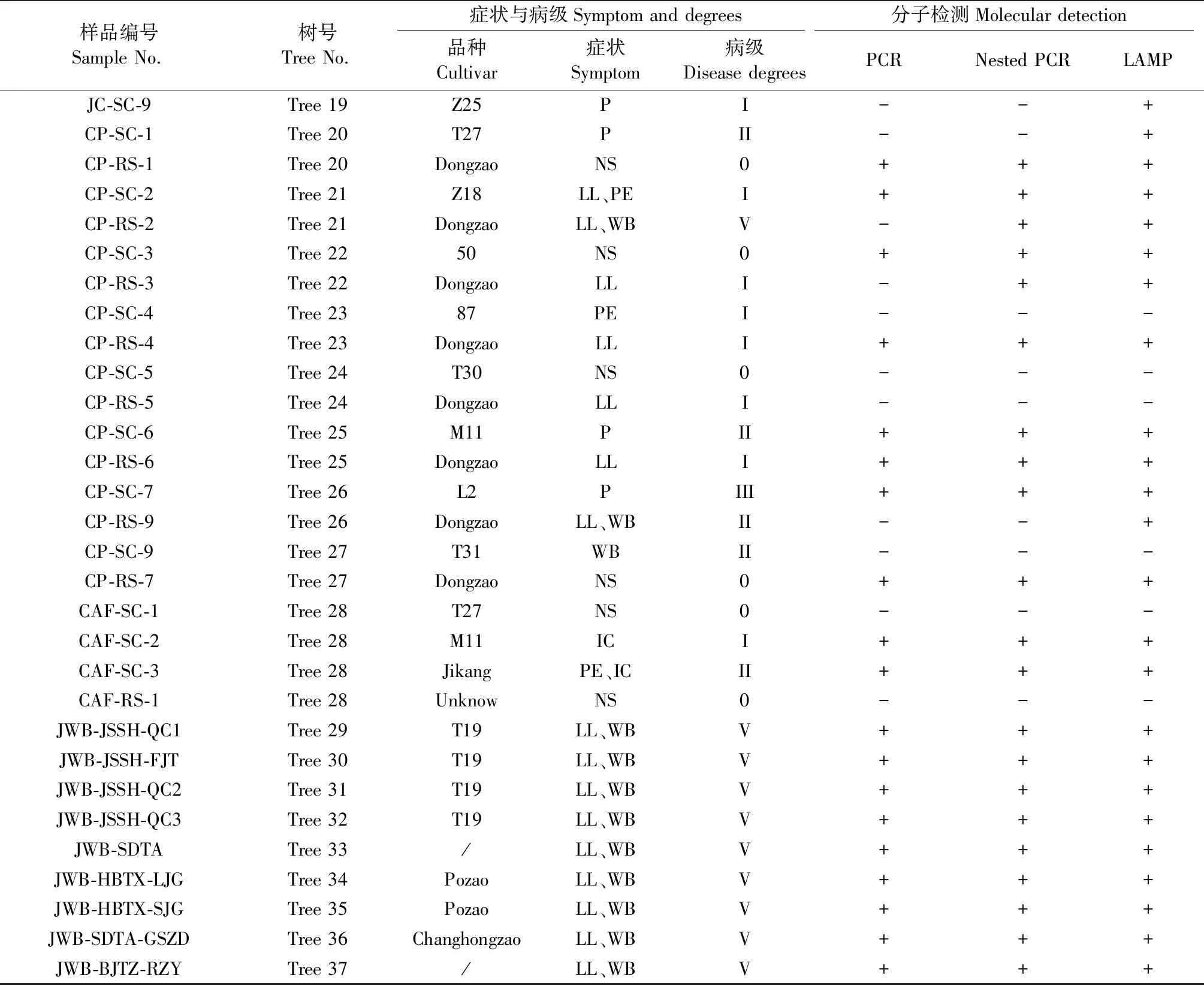
續表2 Continued
中抗品系蜂蜜罐M11接穗嫁接接種后的表現復雜多樣,初期表現出葉片脈間褪綠、花梗延長,進一步表現葉片黃化、卷葉(上卷)、部分葉片局部褐化壞死、部分新梢出現小葉、叢枝、花變葉等癥狀,后期部分變態葉又能逆轉抽出近正常形狀和葉色的葉片。
待鑒定品系Z18接穗嫁接初期也觀察到部分花梗延長癥狀,但后期開花均正常發育且當年結果,枝葉生長正常,表現出對植原體的高度抗性(HR);而且其嫁接砧木枝條的叢枝癥狀也明顯減輕、個別萌條叢枝癥狀消失。
從北京古棗樹篩選的抗病品系50號在三地的癥狀反應有明顯差異。在玉泉山與Z18嫁接同一母株不同分支砧木上的50號接穗當年癥狀嚴重,其下部砧木也發病嚴重(皆為V級);昌平接穗則表現為無癥;而在唐縣軍城的50號是嫁接在無癥的酸棗砧木上,但當年也感染發病嚴重。
巢式PCR檢測結果表明,來源于玉泉山苗圃的棗樹抗病品系的采穗樹Z18、T27、T30、M11均未檢測到植原體;但用LAMP的復核檢測驗證結果顯示,除T30外,Z18、M11、T27無癥采穗母樹皆檢測陽性。而嫁接到發病砧木上的一直未表現癥狀的T30接穗用直接和巢式PCR及LAMP均檢測不到植原體的感染,即使在遮蔭條件下表現出小葉叢枝的樣品中也未檢測到植原體,表現出高度抗感染能力。Z18接穗嫁接到北京玉泉山資源圃和河北唐縣軍城的發病砧木上皆表現為健康無癥或個別黃葉癥狀,其中玉泉山黃葉未檢測到植原體,而軍城無癥樣品卻檢測到植原體。抗病接穗T27黃葉癥狀部位同樣檢測到植原體,而對花變葉部位的檢測結果則是玉泉山資源圃的樣品用直接和巢式PCR皆檢測到植原體,而昌平棗園嫁接的T27和Z25及感病的L2接穗花變葉部位未檢測到植原體,但以上樣品用LAMP卻檢測到植原體。對中度抗病的M11接穗不同癥狀樣品的檢測結果顯示,脈間褪綠、花梗延長、叢枝、小葉和花變葉部位皆易于檢測到植原體的感染。對昌平嫁接的50號無癥接穗用3種方法都能檢測到植原體感染。表現花變葉、小葉叢枝等癥狀乃至無癥的感病接穗與砧木樣品包括冬棗、泗洪大棗、婆棗等品系、酸棗及不同地區采集的典型棗瘋病樣品大多用直接PCR即檢測到植原體,LAMP復核驗證結果也與PCR結果完全吻合,表明這些樣品植原體濃度相對較高(表2)。
2.2 棗瘋植原體株系變異分析
2.2.1 不同靶標基因和基因間區序列的PCR擴增和測序 對采集樣品進行的植原體其他基因與基因間區的PCR擴增和直接測序,與序列比對(BLAST)結果顯示,42條16S rDNA序列、42條SR基因序列、31條secA基因序列和32條tuf基因序列各自同源性為100%,而himA及其下游假定蛋白基因序列以及fusA-tuf基因間區,序列變異率分別為9.6%和18.9%(表3)。且這些序列擴增及比對結果也表明棗瘋植原體株系的均一性,即皆屬于16SrV-B亞組的候選種Candidatusphytoplasmaziziphi;也進一步驗證了直接PCR、巢式PCR和LAMP檢測植原體的結果,從而保障測定結果的準確性與可靠性。
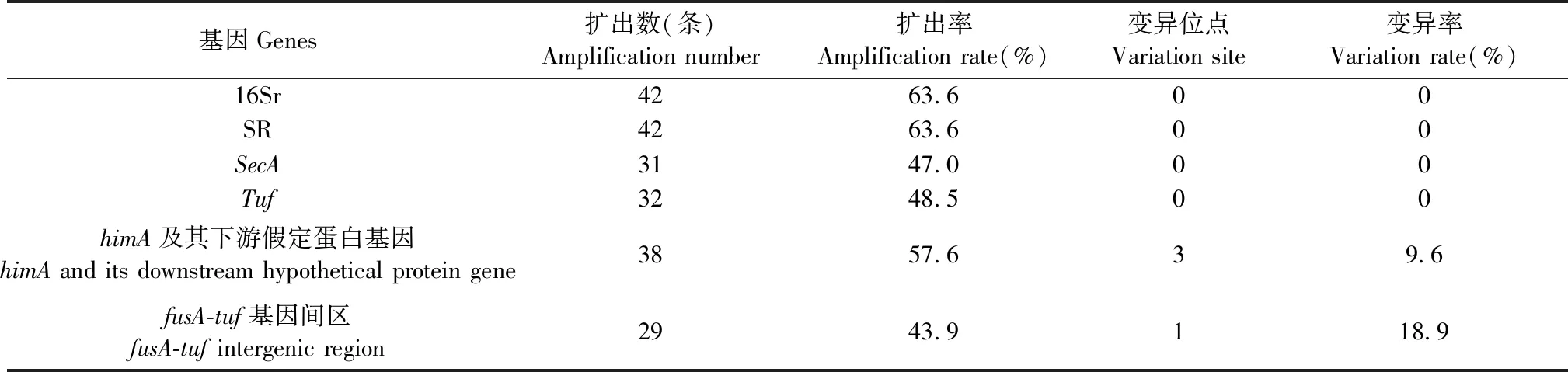
表3 棗瘋植原體基因擴增測序及各基因序列變異情況Tab.3 Sequences from gene amplification and variations of gene sequences of jujube switches’broom phytoplasma
2.2.2himA及下游假定蛋白(HP)基因序列分析himA基因編碼整合宿主因子亞基,在 DNA 修復和復制蛋白方面起作用(王景麗,2016)。Wang等(2018)在JWB全基因組中公布了himA和下游假定蛋白基因序列。通過序列比對在OY-M、AYWB基因組中也發現了同源性序列(圖1)。從38份樣品中擴增himA及其下游假定蛋白基因序列并測序,得到長度約523 bp的himA及其下游假定蛋白基因序列。分析其中313 bp的假定蛋白基因序列片段,在117 bp位點,有堿基的顛換;在154 bp位點,河北唐縣軍城鎮砧木3(嫁接接穗為Z18)有1個堿基A的插入;在190 bp位點,有堿基的顛換。根據3個位置堿基變化情況,將所測得38份樣品及洋蔥黃化和翠菊黃化植原體的himA及其下游假定蛋白基因序列,大體分為3種序列型,C-C型、C-A型和T-A型(表4)。

表4 38份樣品棗瘋植原體himA及其下游假定蛋白基因序列變異位點Tab.4 Variations of himA and its downstream hypothetical protein gene from 38 samples in three jujube orchards
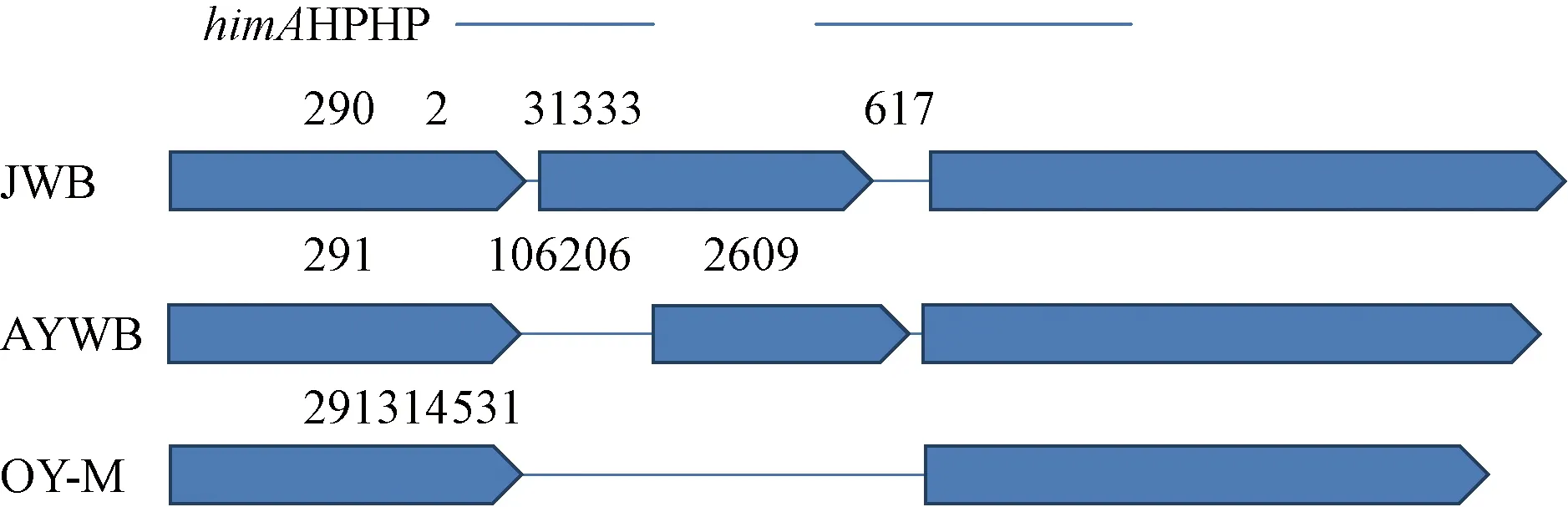
圖1 OY-M、AYWB和JWB植原體himA及其下游假定蛋白基因位置和結構Fig.1 Gene structure diagram of himA and its downstream hypothetical protein gene from OY-M,AYWB and JWB
系統發育關系分析結果顯示,測序菌株大致可聚于這3種序列型。其中軍城鎮樣品序列聚于T-A型,玉泉山樣品與河北唐縣梁家溝棗瘋序列聚于C-A型,而昌平樣品和泗洪、泰安、通州棗瘋序列同聚于C-C型。以上結果顯示出明顯的地域性分化特點(圖2)。
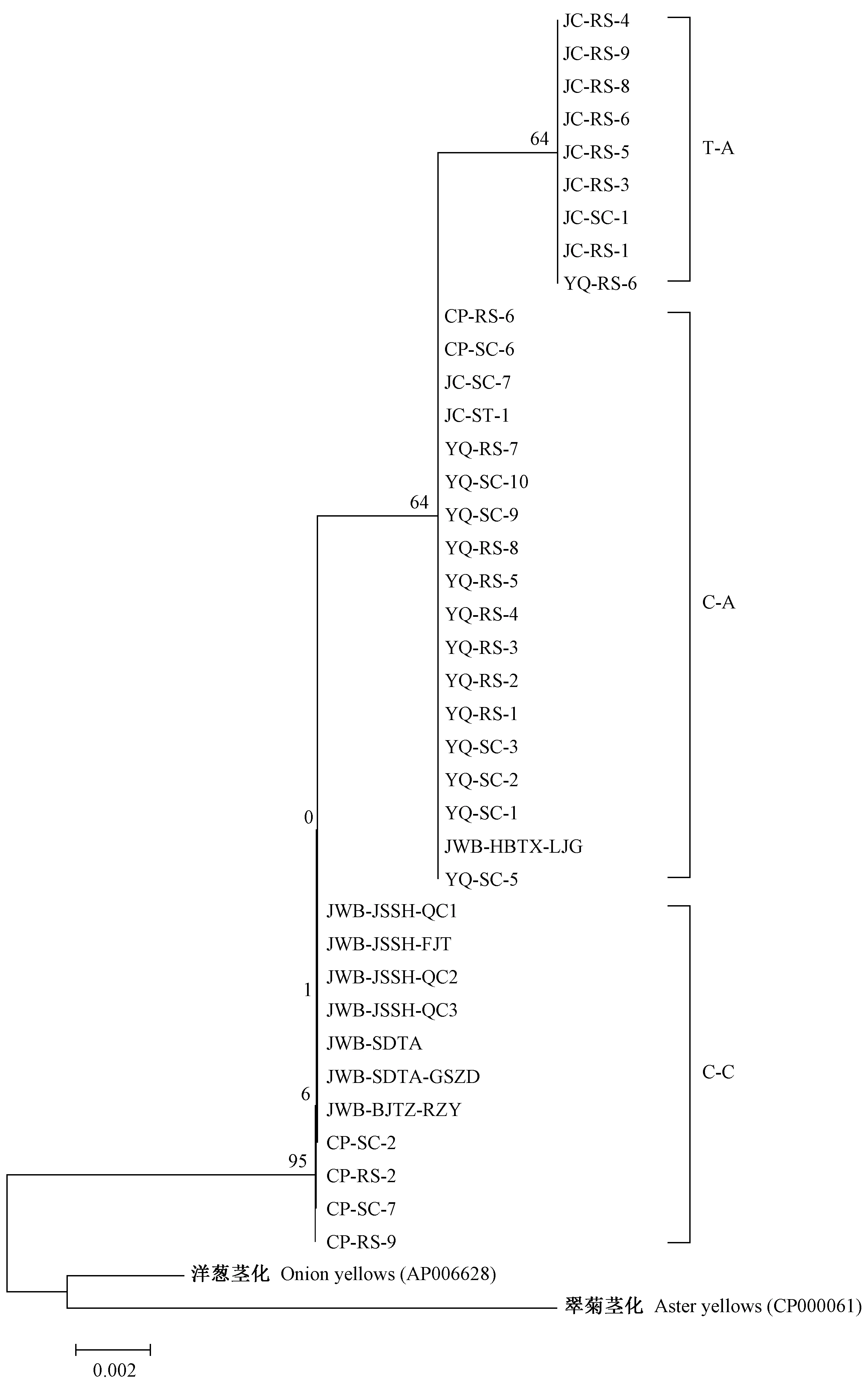
圖2 38份棗瘋病材料基于himA及其下游假定蛋白基因序列構建的系統發育樹(MEGA 5)Fig.2 Phylogenetic tree based on himA and its downstream hypothetical protein gene from 38 samples in three jujube orchards constructed with MEGA 5
2.2.3fusA-tuf基因間區的序列分析 經擴增和測序,得到長度為338 bp的片段序列共29份,其中fusA-tuf基因間區序列長度為53 bp,在間區39 bp位置有堿基A插入/缺失。于少帥等(2016)將16SrV組不同植原體株系fusA-tuf基因間區序列分為2種變異類型,變異類型TJ1(A-)和TJ2(A+)。其中TJ1型包括棗瘋病江蘇南京、江蘇鎮江、北京海淀、遼寧大連3個株系、遼寧葫蘆島2個株系、遼寧建昌、河南周口、河南洛陽、安徽合肥、陜西渭南3個株系和重慶武隆2個株系,以及四川西昌櫻桃致死黃化植原體株系和安徽合肥重陽木叢枝植原體株系。TJ2型僅包括河南濮陽棗瘋植原體株系和北京海淀2個黃金槐叢枝植原體株系。
玉泉山和軍城鎮樣品大都屬于TJ2型,國內其他地區各株系和昌平樣品大都為TJ1型。而YQ-RS-6、CP-SC-6以及CAF-SC-3是各地比較特殊序列株系,即與各地優勢株系的類型不同。這3個株系在himA及其下游假定蛋白基因序列中也屬于特殊株系,即玉泉山砧木(接穗為T30)屬于軍城為主的T-A型,昌平M11接穗(砧木6)屬于玉泉山為主的C-A型。50號古樹接穗樣品的昌平株系屬于TJ1型,而玉泉山和軍城鎮株系屬于TJ2型(圖3)。
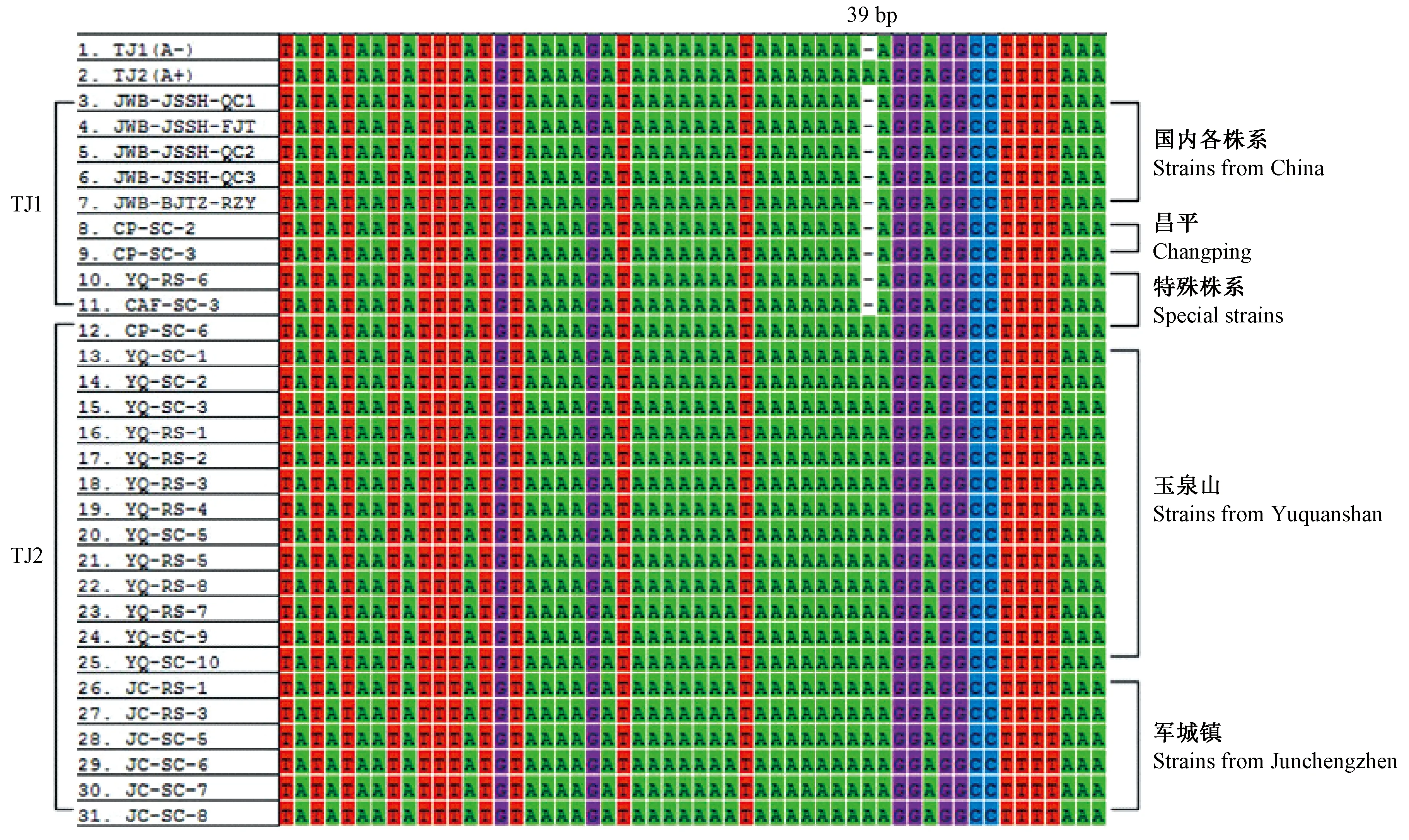
圖3 29份樣品與不同棗瘋植原體fusA-tuf基因間區變異位點Fig.3 Variations of fusA-tuf intergenic region from 29 samples in three jujube orchardsTJ1(A-)表示堿基A缺失,TJ1(A+)表示堿基A插入,TJ3-TJ31為已測定的29個樣品。TJ1(A-)means the deletion of A base.TJ1(A+)means the insertion of A base.TJ3-TJ31 means fusA-tuf genes of 29 samples.
3 討論
3.1 棗樹品種的抗病性
嫁接接后,抗病棗樹接穗的癥狀多數輕于對應的感病砧木,甚至有部分抗性品種接穗表現無癥,如T27、T30、Z18等。同品種接穗在不同棗園有較明顯的癥狀表現差異,如北京古樹50號接穗在昌平王家園棗園表現抗性較好,而在玉泉山苗圃和唐縣軍城鎮棗園則發生V級嚴重小葉叢枝癥狀。這可能與地域環境差異有關,而三地棗瘋植原體株系的致病性分化也是潛在因素。中抗品系T30接穗的抗棗瘋病反應較為特殊,較少產生抗病品種T27、Z17、M11等品種接穗的黃葉、卷葉等抗病表征;也無感病品種接穗的叢枝、花變葉等典型癥狀;在極端遮蔭條件下的接毒接穗僅產生小葉、矮縮和輕微腋芽萌生;接毒后無癥接穗與健康對照接近,能正常開花結果,且無畸形病果。
本研究首次對山西農科院園藝所提供的‘駿優2號’優良品系進行了抗棗瘋病能力測定。依據接穗嫁接于IV-V級病砧木上3個月后,接穗發病癥狀、病級和病情指數劃分棗樹品種抗病/感病類型(田國忠等,2013),對3個棗園不同棗樹品種接穗棗瘋病情指數分析,Z18品種的9枝接穗(嫁接于軍城鎮2株婆棗和玉泉山1株泗洪大棗)均未發病,病情指數為0,暫定其為高度抗性品種(HR)。
3.2 棗瘋植原體himA及其下游假定蛋白基因和fusA-tuf基因間區的序列差異
本研究首次檢測到在這3個棗園內棗瘋植原體株系的himA及其下游假定蛋白基因序列與fusA-tuf基因間區存在明顯的變異規律,即不同變異類型株系與地域差異有關,而與棗樹抗病性強弱明顯無相關性。通過對himA及其下游假定蛋白基因序列比對,可以將所測序3個棗園樣品和國內幾個地區棗瘋樣品較清晰地分為3個類型。第1類為昌平棗園和國內幾個地區菌株,第2類為玉泉山棗園菌株,第3類為軍城鎮棗園菌株。而fusA-tuf基因間區序列分析也支持了這個結論,尤其在幾個特殊序列株系玉泉山砧木(接穗為T30)、昌平M11接穗以及玉泉山林科院冀抗接穗的植原體地域變異上,himA及其下游假定蛋白基因序列與fusA-tuf基因間區序列的分析結果相吻合。而在tuf基因、secA基因、secY基因序列上并未發現明顯變異,一方面表明引起棗瘋病的植原體都屬于16SrV-B亞組植原體,并未發現與該組其他亞組植原體的混合感染,更未發現其他組植原體引致的棗瘋病(徐啟聰等,2009),同時也表明這些保守持家基因的相對穩定性。
關于玉泉山棗園棗瘋病的發生原因,特別是植原體來源,也是值得探討的問題。最早推測為移栽泗洪大棗過程中誤傳入病原,2016年對江蘇泗洪進行了棗瘋病調查采樣,本研究對該株系himA及其下游假定蛋白基因序列與fusA-tuf基因間區2個序列變異分析并未證實這種推論。江蘇的泗洪大棗棗瘋樣品序列與玉泉山的優勢株系序列明顯不同,反而與昌平棗園優勢株系序列有一定關聯,故判斷玉泉山病原可能是當地媒介昆蟲傳播感染或者通過其他來源的無癥帶菌接穗或種苗而傳入該資源圃。因嫁接繁殖接穗來源多樣,更進一步的溯源尚需要對所有引種地的相應樣品的比較分析。而上述himA及其下游假定蛋白基因序列與fusA-tuf基因間區可以用作首選的靶標序列。
3.3 棗樹抗病性鑒定要綜合評價,尤其注意抗病穩定性
品種的遺傳和抗病特性、病原變異、砧木種類與癥狀嚴重度、棗樹生長和環境條件都會對抗病性鑒定的準確性和客觀性產生影響,在抗病資源評價和開發利用時應統籌考慮。從北京古棗樹篩選的50號品系為普通抗性類型,在強致病菌株存在或自身生長勢較弱的情況下可能抗性表現較弱或喪失抗性;雖然已發現昌平菌株與玉泉山菌株的明顯的分子差異,但是否昌平菌株致病性低于玉泉山菌株、且與所揭示的分子變異有關系尚待進一步研究證實(王合等,2018)。
高抗品種T27和中抗品種T30接穗在玉泉山棗園一處林蔭位置表現癥狀為V級小葉叢枝癥狀,推測是林蔭位置光照、濕度和通風條件較差,影響棗樹有機物和抗性物質積累,從而發病嚴重。棗樹是喜光樹種,遮蔭會嚴重影響接穗的光合作用、抑制抗病基因的表達和抗病物質的合成(田國忠等,2013)。Liu等(2016)比較了植原體感染對高抗品種“星光”和感病婆棗光合色素和活性的變化,認為抗性品種的光合響應模式有助于其產生對植原體感染的更強免疫力。而對同一抗性品種接穗在不同光照條件的反應,本研究也做了初步探討。因此,采用適當的栽培管理方式、合理密植、適時修剪等可能會減輕棗瘋病害發生和流行。
4 結論
本研究對嫁接傳毒試驗后3個棗園不同棗樹材料進行癥狀觀察、分子檢測和分子變異分析,首次測定山西園藝所供試的駿優2號(Z18)為高抗品系;發現棗瘋植原體株系差異與地域有關,而與棗樹品種抗性強弱無關;初步排除了玉泉山棗園病原來源于江蘇的泗洪大棗移栽傳病。
