轉(zhuǎn)多基因107楊對目標害蟲的抗性檢測*
2022-01-18 06:56:48韓昆瑾郭娟娟呂梓晴李玉言王世杰楊敏生王進茂
林業(yè)科學(xué) 2021年11期
關(guān)鍵詞:檢測
韓昆瑾 郭娟娟 呂梓晴 李玉言 王世杰 楊敏生 王進茂
(河北農(nóng)業(yè)大學(xué)林學(xué)院 河北省林木種質(zhì)資源與森林保護重點實驗室 保定 071001)
楊樹(Populus)是世界上分布范圍最廣、用途最多、適應(yīng)性最強的樹種之一。在我國,楊樹主要應(yīng)用于木材生產(chǎn)、生態(tài)防護和通道綠化等方面,由于其具有高生長率和易繁殖等特點而受到廣泛重視。然而,楊樹在栽培種植過程中易受到蟲害威脅,食葉害蟲中鱗翅目(Lepidoptera)昆蟲美國白蛾(Hyphantriacunea)及鞘翅目(Coleoptera)昆蟲柳藍葉甲(Plagioderaversicolora)等危害較為嚴重。蟲害除影響楊樹生長和木材質(zhì)量外,還會給林業(yè)經(jīng)濟效益和生態(tài)效益帶來巨大損失。據(jù)調(diào)查數(shù)據(jù)顯示,我國每年因病蟲害損失約 880億元,其中直接經(jīng)濟損失145億元,生態(tài)服務(wù)價值損失 735億元(蘇宏鈞等,2004),同時蟲害造成的葉損失會使森林由碳匯變?yōu)樘荚?(Kurzetal.,2008)。利用植物基因工程解決楊樹蟲害問題是一種較為有效的途徑。楊樹作為林木基因工程中的一種模式植物,其在抗蟲轉(zhuǎn)基因研究中也取得了許多突破性進展(Zuoetal.,2018;Zhangetal.,2016),目前,蘇云金芽孢桿菌(Bacillusthuringiensis)晶體蛋白基因(Bt)、蛋白酶抑制劑類基因(PI)和昆蟲毒素基因等已應(yīng)用于楊樹抗蟲轉(zhuǎn)基因研究(睢韡等,2019)。但這些外源基因能否在受體植物中穩(wěn)定表達,還需要進行長期觀測。Hawkins等(2003)檢測了楊樹的44個不同轉(zhuǎn)基因株系,發(fā)現(xiàn)1個沉默株系,推測在木本植物中基因沉默相對較少,且在轉(zhuǎn)錄開始便已沉默。胡建軍等(2007)觀察并檢測了7年生的轉(zhuǎn)Bt基因歐洲黑楊(Populusnigra),調(diào)查結(jié)果表明Bt基因能夠穩(wěn)定表達,且未發(fā)現(xiàn)害蟲對轉(zhuǎn)基因植株產(chǎn)生耐受性。轉(zhuǎn)基因楊樹中外源基因穩(wěn)定性與環(huán)境也存在一定的相關(guān)性,樹木的生長周期較長,基因沉默的可能性增加(魏冰等,2009)。
本研究以河北農(nóng)業(yè)大學(xué)林木遺傳育種實驗室培育的轉(zhuǎn)多基因歐美楊107楊(Populus×euramericana‘Neva’)為植物材料,對6個轉(zhuǎn)基因株系2年生田間苗通過PCR技術(shù)鑒定外源基因是否穩(wěn)定存在,研究Bt基因表達情況以及生長季的毒蛋白含量,以美國白蛾和柳藍葉甲為測試昆蟲檢測轉(zhuǎn)多基因107楊的抗蟲性。通過分析轉(zhuǎn)多基因107楊的外源基因表達情況及表達效率等問題,為轉(zhuǎn)多基因楊樹的品種培育和推廣提供依據(jù)。
1 材料和方法
1.1 試驗材料
1.1.1 植物材料 試驗材料為轉(zhuǎn)Cry1Ac-Cry3A-BADH基因107楊(簡稱V1)株系ECAA1、ECAA2、ECAA3及轉(zhuǎn)Cry1Ac-Cry3A-NTHK1基因107楊(簡稱V2)株系ECAB1、ECAB2、ECAB3。其中,V1載體(圖1a)攜帶抗蟲的Bt基因Cry1Ac和Cry3A,同時攜帶有甜菜堿醛脫氫酶基因BADH,基因Cry1Ac和Cry3A的啟動子為CAMV35S啟動子,Cry1Ac基因接近載體右臂,Cry3A接近載體左臂;V2載體(圖1b)同樣攜帶抗蟲的Bt基因Cry1Ac和Cry3A,同時攜帶有煙草乙烯受體基因NTHK1,基因Cry1Ac和Cry3A的啟動子及位置與V1載體相同。以未轉(zhuǎn)基因107楊為對照。轉(zhuǎn)基因植株為栽植在河北農(nóng)業(yè)大學(xué)西校區(qū)林學(xué)院專用苗圃中的2年生幼樹,正常管理,整個生長過程中未施用任何農(nóng)藥。
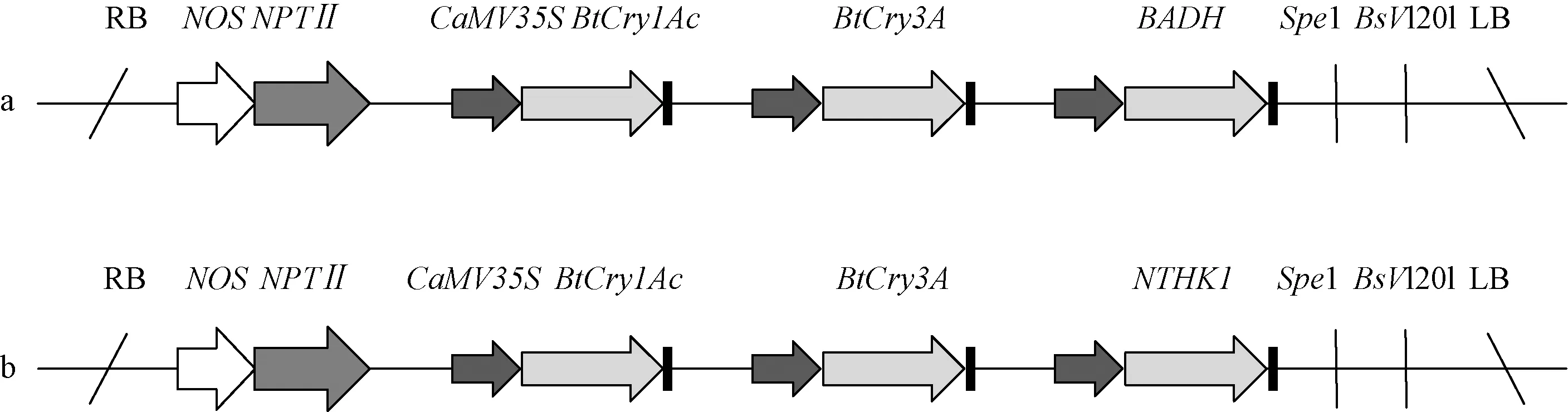
圖1 載體結(jié)構(gòu)Fig.1 The structure of vectora:載體V1結(jié)構(gòu)圖;b:載體V2結(jié)構(gòu)圖。a:The structure of vector V1;b:The structure of vector V2.
1.1.2 測試昆蟲 美國白蛾屬昆蟲綱(Insecta)鱗翅目燈蛾科(Arctiidae),幼蟲卵塊由北京林業(yè)大學(xué)陳敏教授提供(2019年7月)。柳藍葉甲屬昆蟲綱鞘翅目葉甲科(Chrysomelidae),成蟲、幼蟲或卵塊取自保定滿城和河北農(nóng)業(yè)大學(xué)苗圃(2019年7月底至8月初)。
1.2 轉(zhuǎn)基因植株的PCR檢測
采集2年生轉(zhuǎn)基因苗木當年生的長枝葉片,采用改良的 CTAB法提取各轉(zhuǎn)基因株系及對照植株的基因組DNA。以基因組DNA為模板,分別用Cry1Ac、Cry3A基因的正反引物進行PCR檢測,正反引物的DNA序列及目的基因擴增片段見表1。PCR檢測采用20 μL的反應(yīng)體系[ddH2O:12.8 μL;10×PCR Buffer:2 μL;引物R(10 pmol·L-1):1 μL;引物F(10 pmol·L-1):1 μL;DNTP:2 μL;2DNA:1 μL],以含有Cry1Ac和Cry3A基因的質(zhì)粒DNA為陽性對照,以未轉(zhuǎn)基因的107楊的DNA作為陰性對照。PCR反應(yīng)程序為:94 ℃預(yù)變性10 min;94 ℃變性30 s,50 ℃復(fù)性40 s,72 ℃延伸90 s,共30個循環(huán);72 ℃退火5 min。PCR儀購于新加坡Bio-Rad公司,型號為T1000。

表1 引物信息Tab.1 Primer information
1.3 Cry1Ac和Cry3A毒蛋白的ELISA檢測
8月下旬于河北農(nóng)業(yè)大學(xué)西校區(qū)苗圃采集各轉(zhuǎn)基因株系及未轉(zhuǎn)基因歐美楊107楊2年生幼樹頂端完全展開的第3、4葉位的葉片,用于Cry1Ac、Cry3A毒蛋白檢測。采用美國Agdia公司的Bt-Cry1Ab/1Ac和Bt-Cry3A ELISA試劑盒,分別進行檢測,檢測方法參照試劑盒說明書進行。檢測毒蛋白的陽性對照為試劑盒自帶,陰性對照為對照組的107楊,檢測結(jié)果用 BioRad 550型酶標儀進行測定。蛋白濃度以每g新鮮葉片所含毒蛋白質(zhì)量(ng)計算。每個處理進行3次生物學(xué)重復(fù)。
1.4 抗蟲性檢測
美國白蛾的1齡、2齡幼蟲用于檢測Cry1Ac毒蛋白的抗蟲性。1齡供試幼蟲為室內(nèi)孵化1~2天的初孵幼蟲,2齡幼蟲在未進行試驗前以107楊新鮮葉片飼養(yǎng),其后用轉(zhuǎn)基因葉片進行抗蟲性檢測,均設(shè)置對照組,飼喂至少量幼蟲結(jié)繭為止,歷時45天。
柳藍葉甲的1齡、2齡幼蟲及成蟲用于檢測Cry3A毒蛋白的抗蟲性,其幼蟲飼喂試驗方法與美國白蛾大致相同,增加2齡供試幼蟲與1齡幼蟲進行混齡抗蟲性試驗,同時增加成蟲抗性試驗。各試驗均以未轉(zhuǎn)基因107楊作為對照組,飼喂至試驗幼蟲發(fā)育成蛹或成蟲出現(xiàn)大面積死亡為止,歷時20天。
采集各轉(zhuǎn)基因株系和對照組當年生枝條新鮮葉片,將測試昆蟲均勻放置于葉片表面,盡量減少對試驗昆蟲造成損傷,然后將葉片(插于花泥保鮮)安放在透明塑料盒中,盒蓋扎上小孔,保證透氣性。
試驗中,美國白蛾初孵幼蟲各株系設(shè)置2~4個重復(fù),每處理20~30頭;美國白蛾2齡幼蟲各株系設(shè)置3個重復(fù),每處理10頭。柳藍葉甲初孵幼蟲各株系設(shè)置4個重復(fù),每處理30頭;其余蟲期各株系設(shè)置3個重復(fù),1齡、2齡混齡幼蟲每處理20頭,成蟲每處理15頭。
計算公式如下:
總死亡率=Σ(各重復(fù)昆蟲死亡數(shù)/該重復(fù)內(nèi)測試昆蟲總頭數(shù))/重復(fù)數(shù);
校正死亡率=(轉(zhuǎn)基因株系死亡率-對照死亡率)/(1-對照死亡率);
總增長率=(最后一天的體質(zhì)量/體長-第1天的體質(zhì)量/體長)/天數(shù);
取食葉面積率=取食面積/葉片總面積。
2 結(jié)果與分析
2.1 轉(zhuǎn)基因植株的PCR檢測
利用Cry1Ac和Cry3A基因的特異性引物對6個轉(zhuǎn)基因株系2年生田間苗DNA進行PCR 擴增,以帶有目的片段的質(zhì)粒為陽性對照,未轉(zhuǎn)基因107楊為陰性對照。PCR擴增結(jié)果顯示,在6個轉(zhuǎn)基因107楊株系DNA中,均能檢測到Cry1Ac基因(圖2a,特異性條帶為546 bp)和Cry3A基因(圖2b,特異性條帶為667 bp),而對照組均未檢測到特異性條帶,說明轉(zhuǎn)多基因107楊在生長發(fā)育過程中,未發(fā)生外源目的基因的丟失。
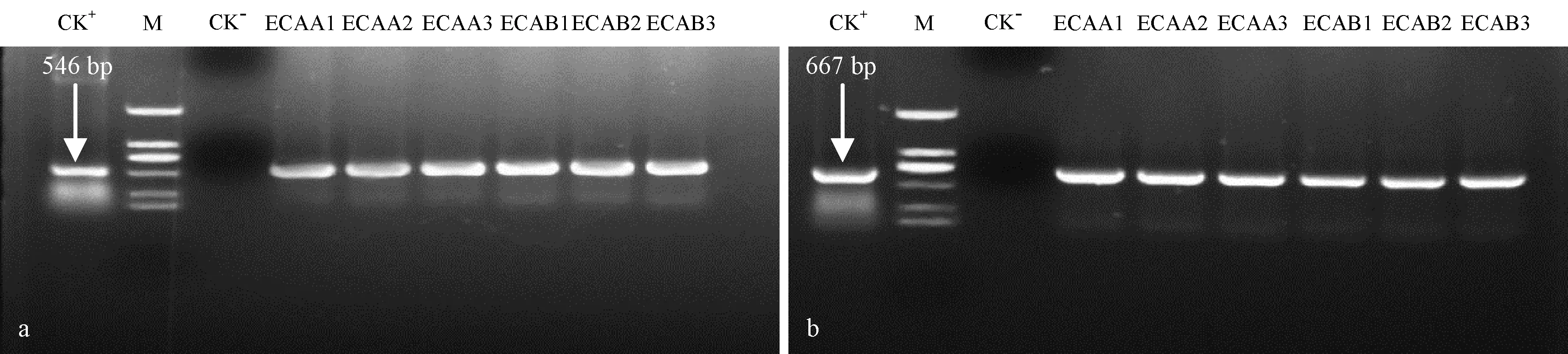
圖2 外源基因的PCR檢測Fig.2 PCR amplification of the exogenous genes of various transgenic linesa:Cry1Ac基因的PCR檢測;b:Cry3A基因PCR檢測。CK+:質(zhì)粒p09687199;CK-:陰性對照(未轉(zhuǎn)基因植株);M:DL2000 DNA Marker(自上而下為2 000,1 000,750,500,250,100 bp)。ECAA1-3:轉(zhuǎn)Cry1Ac-Cry3A-BADH基因的3個株系;ECAB1-3:轉(zhuǎn)Cry1Ac-Cry3A-NTHK1的3個株系。a:PCR detection of Cry1Ac;b:PCR detection of Cry3A. CK+:Plasmid p09687199;CK-:Control (Untransformed line);M:DL2000 DNA Marker (From up to down,these are 2 000,1 000,750,500,250,100 bp).ECAA1-3:Three lines transformed with Cry1Ac-Cry3A-BADH gene;ECAB1-3:Three lines transformed with Cry1Ac-Cry3A-NTHK1 gene.
2.2 轉(zhuǎn)基因植株的Bt毒蛋白檢測
8月下旬采用ELISA法對轉(zhuǎn)多基因107楊葉片中Cry1Ac和Cry3A毒蛋白進行檢測,結(jié)果(圖3)表明,轉(zhuǎn)基因株系均含有2種Bt毒蛋白,但對照組均未檢測到。各轉(zhuǎn)基因株系Cry1Ac毒蛋白含量(3.60~20.76 ng·g-1)均明顯低于Cry3A毒蛋白含量(3 479.66~7 230.29 ng·g-1),二者相差1 000倍左右。各株系間Cry1Ac毒蛋白含量存在顯著差異,株系ECAA1和ECAB3中含量較高,而株系ECAA3和ECAB2中含量較少。顯著低于其他株系。各轉(zhuǎn)基因株系中Cry3A毒蛋白含量均較高,且各株系間也存在顯著差異,株系ECAA1中毒蛋白含量最高,株系ECAA3與ECAB1中毒蛋白含量較少。在各株系中ECAA1的Cry1Ac和Cry3A毒蛋白含量均是最高的。

圖3 轉(zhuǎn)基因107楊不同株系的Cry1Ac和Cry3A毒蛋白含量Fig.3 Content of Cry1Ac and Cry3A toxin protein in different lines of transgenic 107 poplar
2.3 轉(zhuǎn)基因株系的抗蟲性檢測
2.3.1 對美國白蛾幼蟲的抗性 用轉(zhuǎn)多基因107楊飼養(yǎng)美國白蛾1齡幼蟲14天,結(jié)果如表2所示。各株系之間美國白蛾的致死效果存在顯著差異,株系ECAA2、ECAA3和ECAB1對1齡幼蟲的致死效果較好,其中ECAB1抗性最高,其校正死亡率為100%。美國白蛾幼蟲在前6天死亡個數(shù)較多,隨后死亡率降低,但在12~14天,幼蟲的死亡個數(shù)突然增加,這可能與Cry1Ac毒蛋白的積累有關(guān)。
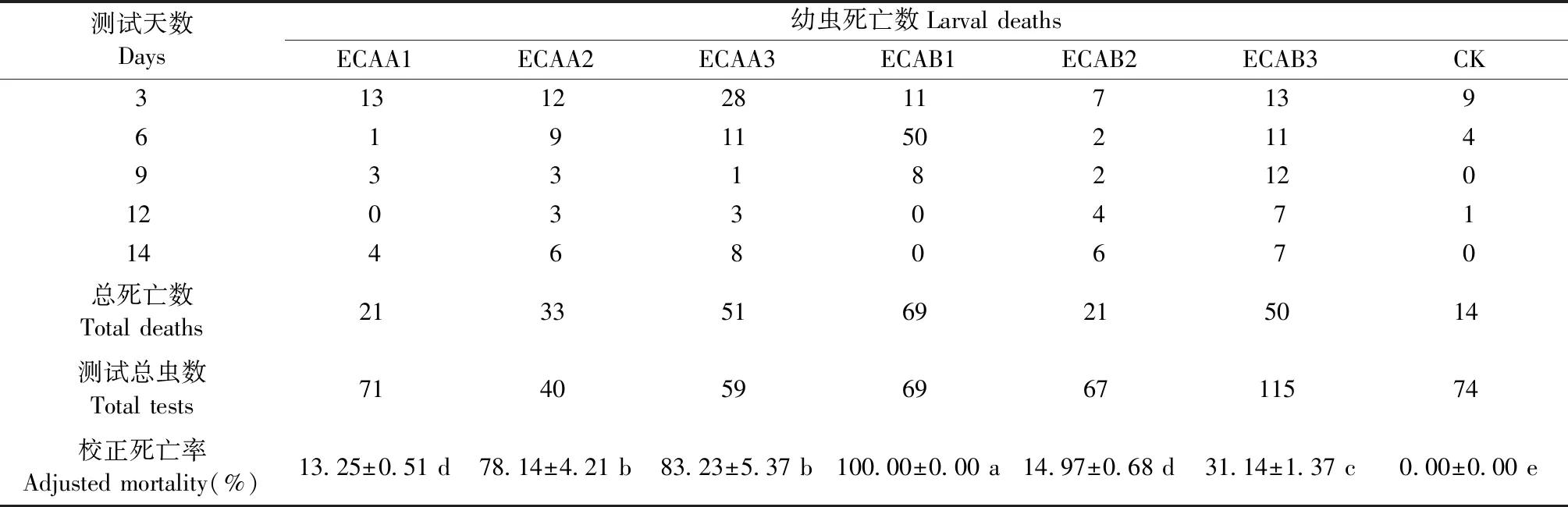
表2 轉(zhuǎn)基因株系對美國白蛾1齡幼蟲的致死作用①Tab.2 Lethal effect of transgenic lines on the 1st instar larvae of Hyphantria cunea
2.3.2 對美國白蛾的滯育效應(yīng) 用各轉(zhuǎn)基因株系與對照組107楊飼喂美國白蛾1齡幼蟲,圖4為飼喂不同天數(shù)時美國白蛾的各齡級分布。從圖可以明顯看出,美國白蛾的發(fā)育進程,對照組植株在7天時已全部進入2齡以上,而轉(zhuǎn)基因株系ECAA1、ECAA2和ECAA3中,仍存在很多1齡幼蟲,其中株系ECAA2的滯育表現(xiàn)最好。其他株系在7天時均出現(xiàn)3齡幼蟲,僅ECAB1的幼蟲全部處于2齡階段。到達13天時,對照組已大部分進入4、5齡,此時轉(zhuǎn)基因株系ECAB1的幼蟲已全部死亡,轉(zhuǎn)基因株系ECAA2仍存在1齡幼蟲,ECAA1、ECAA3有較多的2齡幼蟲。由此可見,轉(zhuǎn)基因株系ECAA1、ECAA2、ECAA3和ECAB1對美國白蛾1齡幼蟲有較為明顯的抑制其生長發(fā)育的作用,即延緩其發(fā)育進度;而另外2個株系對美國白蛾生長發(fā)育無明顯抑制作用。
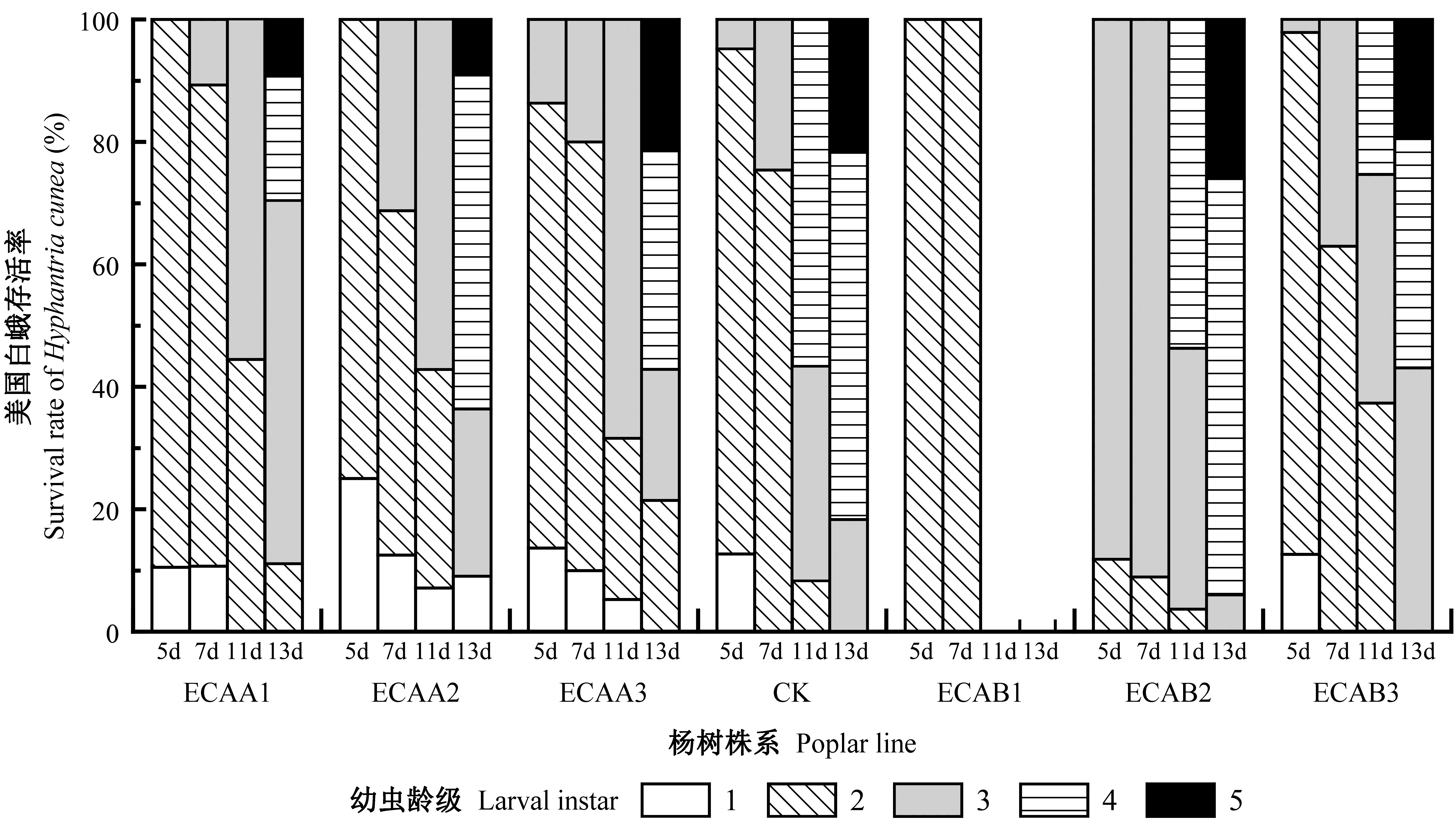
圖4 美國白蛾初孵幼蟲發(fā)育進度Fig.4 Developmental progress of the first hatched larvae of Hyphantria cunea
用轉(zhuǎn)基因株系和對照飼喂的2齡幼蟲,結(jié)果見表3、圖5和圖6,可以看出各轉(zhuǎn)基因株系取食率皆低于對照組107楊。在8月6日至7日和8月14日至15日期間對照組美國白蛾的采食率、體質(zhì)量及體長均高于轉(zhuǎn)基因各株系,特別是株系ECAA1、ECAA3、ECAB1、ECAB2均明顯抑制了美國白蛾2齡幼蟲的取食活動,進而影響其生長發(fā)育,且各株系間統(tǒng)計學(xué)差異顯著。由表3可看出,在8月6日至7日時間段內(nèi),對照組107楊取食面積最高,而從圖6也可以看出轉(zhuǎn)基因107楊株系ECAA3、ECAB1、ECAB2的取食面積較低,由此表明目的基因在受體107楊中得到較好表達。
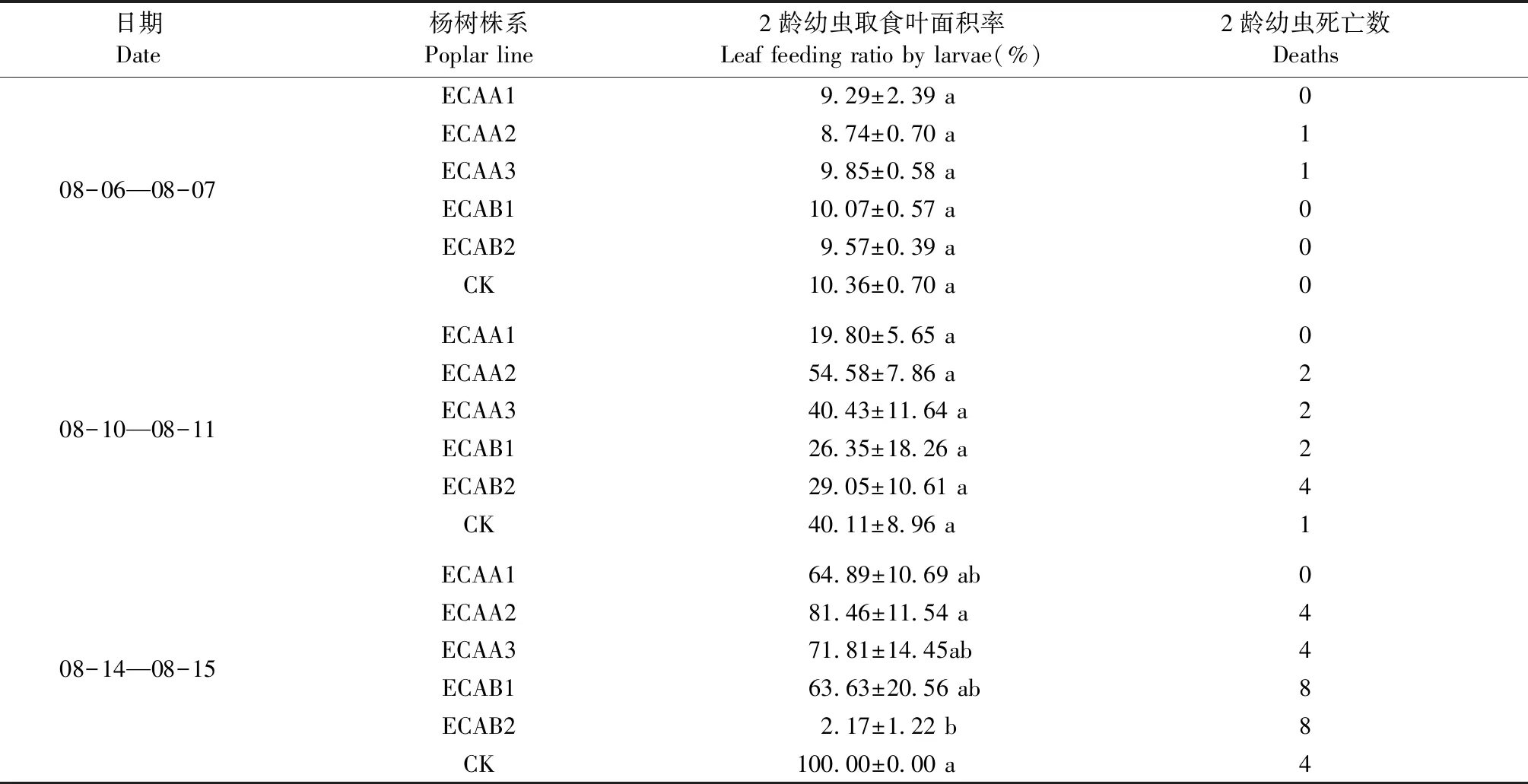
表3 美國白蛾2齡幼蟲平均采食率與死亡數(shù)①Tab.3 Average feeding rate and mortality of the 2nd instar larvae of Hyphantria cunea

圖5 美國白蛾2齡幼蟲平均體長、質(zhì)量Fig.5 Average body length or weight of the 2nd instar larvae of Hyphantria cunea
2.3.3 轉(zhuǎn)基因株系對柳藍葉甲的抗性檢測 圖7結(jié)果顯示,轉(zhuǎn)基因株系ECAA2對柳藍葉甲無論幼蟲還是成蟲致死率皆為最低,表現(xiàn)中低抗蟲性。除株系ECAA2外,其他轉(zhuǎn)基因株系對柳藍葉甲1齡幼蟲的致死率均達到100%。對1、2齡混齡幼蟲的致死率均在85%~100%之間,其中株系ECAB2 和ECAB3的致死率為100%。對于成蟲,株系ECAA3、ECAB1、ECAB2和ECAB3的致死效果較好,致死率在90%以上,且6個轉(zhuǎn)基因株系之間存在顯著性差異。圖8可以看出轉(zhuǎn)基因株系明顯抑制了柳藍葉甲成蟲的取食。
3 討論
植物的大多數(shù)性狀和生理功能都是通過多基因的協(xié)調(diào)表達實現(xiàn)的,而轉(zhuǎn)多基因植株可通過協(xié)調(diào)控制多種外源基因的表達來綜合改善植物的性狀(初易洋等,2016)。多基因轉(zhuǎn)化指將2個及以上的外源基因轉(zhuǎn)入受體植株中,且轉(zhuǎn)化的多基因可以是不同的抗性基因或是改良植物性狀的基因。楊夢悅(2006)將DREB(調(diào)節(jié)逆境脅迫)、IRT1(提高耐缺鐵性)和rolC(毛狀根誘導(dǎo))融合三價基因 (Rirol)轉(zhuǎn)入八棱海棠(Malusrobusta),其轉(zhuǎn)基因植株明顯表現(xiàn)出耐鹽、矮化和生根力強等性狀。王建革等(2006)發(fā)現(xiàn)轉(zhuǎn)SacB、vgb、BtCry3A+OC-Ⅰ、調(diào)節(jié)基因JERF36及報告基因NPTⅡ等基因的庫安托楊(Populus×euramericana‘Guariento’)具有抗鹽堿能力的同時,又具有Cry3A基因的抗蟲性。另外在對Bt基因的研究中發(fā)現(xiàn),當2個Bt基因同時存在時,楊樹表現(xiàn)為雙抗性,從而擴大抗蟲譜,提高抗蟲效果(張超等,2019)。
本研究中,采用2年生轉(zhuǎn)多基因107楊株系的葉片進行外源基因表達檢測和抗蟲試驗,顯示外源基因仍存在于基因組中,2種Bt毒蛋白均存在,且對美國白蛾和柳藍葉甲表現(xiàn)雙抗性,說明2年生轉(zhuǎn)基因楊樹的外源基因依舊穩(wěn)定表達。李秀芬(2008)在轉(zhuǎn)Bt基因的南林895楊(Populus×euramericana‘Nanlin895’)研究中證明其繼代培養(yǎng)4年后仍然攜帶外源目的基因;陳虞超等(2014)在對9年生抗蟲轉(zhuǎn)基因銀河楊(Populusalba×P.hopeiensis)的抗蟲性檢測中發(fā)現(xiàn),該轉(zhuǎn)基因銀河楊的外源抗蟲基因仍然表達正常。充分證明轉(zhuǎn)多基因植物的外源基因能夠保持長時間穩(wěn)定存在和表達。
本試驗中,8月末各轉(zhuǎn)基因株系中Cry1Ac毒蛋白在葉片中含量遠遠低于Cry3A毒蛋白在葉片中的含量,Cry3A毒蛋白在葉片中含量約是 Cry1Ac毒蛋白的103~104倍。張超等(2019)對1年生轉(zhuǎn)Bt基因107楊毒蛋白的時空表達進行檢測,發(fā)現(xiàn)Cry1Ac毒蛋白含量在9月份達到最高,Cry3A毒蛋白含量在8月份達到最高;徐麗娜等(2015)認為蛋白質(zhì)含量的季節(jié)性和時空變化是植物體內(nèi)毒蛋白的積累和其代謝過程所造成的,理論上蛋白含量應(yīng)增加。但經(jīng)試驗發(fā)現(xiàn)轉(zhuǎn)抗蟲基因三倍體毛白楊的抗蟲性雖表達穩(wěn)定,但隨著樹齡的增大Bt基因的表達強度會減弱(李立,2009),具體原因有待研究。
本試驗的轉(zhuǎn)化載體V1及V2的啟動子均為CaMV35S啟動子,4個外源基因分別為Cry3A、Cry1Ac、BADH和NTHK1,其中,Cry1Ac基因處于上游,Cry3A處于下游。試驗結(jié)果顯示,Cry1Ac毒蛋白含量明顯低于Cry3A。Xu等(2016)認為在載體中上游Bt基因的表達水平較低;而張冰玉等(2006)認為是由于Cry1Ac基因和Cry3A基因的同源性高,易出現(xiàn)共抑制狀況。轉(zhuǎn)多基因工程中存在的基因沉默等問題,成為了影響受體植物外源基因表達穩(wěn)定性和高效性的巨大障礙,而引起基因沉默的原因大致分為3類:第1種為位置效應(yīng)引起的基因沉默,可以通過調(diào)整目的基因在載體中的排列位置與方向,或是在外源基因的兩側(cè)加上MAR結(jié)構(gòu),顯著減少共抑制狀況,提高外源基因表達水平的穩(wěn)定性和效率,如Qiu等(2017)發(fā)現(xiàn)2個Bt基因處于相反方向排列時,對2個目的基因的表達均有較強的促進作用;第2種為轉(zhuǎn)錄水平的基因沉默,主要是由啟動子甲基化引起,Giho等(2012)研究發(fā)現(xiàn)轉(zhuǎn)入受體植物本氏煙草(Nicotianabenthamiana)的花椰菜病毒,因35S啟動子發(fā)生甲基化導(dǎo)致其基因沉默,啟動子的甲基化對轉(zhuǎn)基因的遺傳穩(wěn)定性產(chǎn)生負面影響,可使用去甲基化試劑、修飾外源基因的序列或是選取有效促動劑等方法解決轉(zhuǎn)錄水平的基因沉默問題;第3種轉(zhuǎn)錄后水平的基因沉默,應(yīng)通過盡量避免或是減少基因之間的同源性等方法解決(李寶健等,2005)。
本試驗通過統(tǒng)計幼蟲、成蟲的致死率、發(fā)育進度、蟲長和蟲質(zhì)量,篩選出具中高雙抗性株系ECAA3和ECAB1,但發(fā)現(xiàn)這2個株系的Cry1Ac毒蛋白和Cry3A毒蛋白含量與抗蟲性之間無明顯規(guī)律性,毒蛋白含量與抗蟲性關(guān)系還有待進一步研究,同時再次驗證轉(zhuǎn)基因楊樹能夠顯著毒殺靶標害蟲。另外,轉(zhuǎn)基因株系對低齡幼蟲有較強致死效果,因低齡幼蟲對Bt毒蛋白較為敏感,其作用機理為Bt毒蛋白進入幼蟲體內(nèi)轉(zhuǎn)化成活化了的毒素與腸道上皮的特異受體結(jié)合,形成穿孔導(dǎo)致幼蟲死亡(Batesetal.,2005);而對高齡幼蟲的滯育效果較明顯,可能是隨著蟲齡的增長,蟲體的解毒酶活力增加,而抗蟲毒蛋白被降解,故死亡率降低,但可以有效地抑制幼蟲的體長與體質(zhì)量的增長和發(fā)育速率(田亞坤等,2014)。轉(zhuǎn)多基因庫安托楊具有抗旱、抗?jié)场⒖果}堿和抗鞘翅目昆蟲的特點(李環(huán),2008),這種轉(zhuǎn)入的多個基因均能在楊樹中穩(wěn)定表達,與本次試驗中所選出的高抗株系中多基因穩(wěn)定表達的結(jié)果相似。
抗蟲基因轉(zhuǎn)化有諸多優(yōu)點,不僅可以實現(xiàn)定向改造植株的目標,而且可以擴大林木的抗蟲譜,達到一種林木具多抗蟲性的目的。但仍存在轉(zhuǎn)化效率低、重復(fù)性差和在受體中不能穩(wěn)定表達等問題,特別是外源基因能否穩(wěn)定保留在基因組中的問題(Zhangetal.,2016)仍需深入研究。因此,為了使外源基因安全穩(wěn)定表達、提高林木品質(zhì)仍需進一步的研究和探討。同時,本試驗中的毒蛋白含量與抗蟲性關(guān)系還待進一步研究,且未對BADH和NTHK1這2種耐鹽基因進行檢測和表達分析,未來需要進一步探究。
4 結(jié)論
轉(zhuǎn)多基因107楊2年生田間苗各株系的外源基因仍然存在且穩(wěn)定表達,不同轉(zhuǎn)基因株系間抗蟲性差異顯著,株系ECAA3對美國白蛾1齡幼蟲具有明顯抑制其生長發(fā)育的作用,株系ECAB1對美國白蛾的致死率最高,ECAA3和ECAB1株系對柳藍葉甲和美國白蛾均有較高抗性。
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
