增氧灌溉對平邑甜茶生長及根區土壤的影響*
2021-12-15 03:40:40尹伊君毛云飛張璐璐胡艷麗毛志泉陳學森
林業科學 2021年10期
尹伊君 毛云飛 楊 露 張璐璐 胡艷麗 毛志泉 陳學森 沈 向
(山東農業大學園藝科學與工程學院 國家蘋果工程技術研究中心 泰安 271018)
灌溉可以使根部吸收充足的水分,保證植物進行正常的生長發育,而灌溉水中氧含量對植物發育具有重要作用。植物根系在生長發育過程中需要消耗充足的氧氣來進行呼吸作用,從而促進植株對土壤中養分和水分的吸收,因此,植物根際氧環境中的氧氣含量是影響植物根系進行正常生長代謝的主導因子(劉磊等, 2012; 劉學等, 2009)。目前,我國傳統果園灌溉的主要方式為大水漫灌,灌溉水會將土壤中氣體趕出,使土壤出現暫時性缺氧環境,而土壤氧氣的供應直接關系著土壤肥力、根際微生物群落結構和根系呼吸速率等(喬建磊等, 2017)。土壤中氧氣供應不足會導致根系呼吸作用減弱,引起土壤還原性物質和有毒物質的積累,減少水分、養分的吸收及向地上部分的運輸,引起作物水分和養分利用效率下降,導致作物生物量下降(Lietal., 2016; Bhattaraietal., 2009; 雷宏軍等, 2017),進而影響果樹的產量及品質。增氧灌溉作為一種緩解缺氧條件的方法,將通過地下滴灌將曝氣水直接輸送至根部(Bhattaraietal., 2006),氣態和溶解在水中的大量氧氣可以輸送至根部區域。因此,增氧灌溉可以同時提供水和氧氣,抵消傳統灌溉方式的負面影響,并改善缺氧癥狀。
增氧灌溉使作物根區的O2濃度和土壤呼吸分別增加2.4%~32.6%和42%~100%(Chenetal., 2010),甜椒(Capsicumannum)增氧灌溉生物量和果實產量分別提高16%和18%(Ityleetal., 2014)。因此,增氧灌溉是提高作物產量的有效灌溉方法(Cuietal., 2020),不僅可以提高水資源的利用率,緩解當前農業用水緊缺的問題,還可以解決因植物根際缺氧導致的產量、品質低劣等問題(Shahienetal., 2014; Duetal., 2018)。
本試驗以蘋果(Maluspumila)常用砧木1、2年生平邑甜茶(Malushupehensis)為試驗對象,采用增氧氣泵通過橡膠管對灌溉水進行增氧,并用便攜式溶解氧測定儀監測灌溉水的氧含量的變化,實現對不同處理的植株進行增氧灌溉。通過不同的增氧灌溉處理生理指標分析,研究該方法對平邑甜茶生長及根區土壤環境的影響,為探明增氧灌溉對大田模式栽培下果樹的影響提供參考。
1 材料與方法
1.1 試驗地概況與試驗設計
山東省泰安市泰山區(36°05′N,117°03′E),山東農業大學園藝科學與工程學院果樹根系實驗室和作物生物學國家重點實驗室,暖溫帶半濕潤季風氣候,四季分明,春季干燥多風,夏季高溫多雨,秋季天高氣爽,冬季寒冷少雪,平均氣溫13.2 ℃,平均降水量687.7 mm,極端最低氣溫-27.5 ℃,極端最高氣溫41 ℃,全年無霜期近200天。
挑選長勢基本相同、沒有病蟲害的1年生、2年生平邑甜茶作為盆栽苗,栽入試驗盆(上內徑25 cm、下內徑18 cm、高18 cm)內。盆土為園土、有機質按照4∶1比例均勻混合而成: 盆土堿解氮97.8 mg·kg-1、速效磷27.9 mg·kg-1、速效鉀82.1 mg·kg-1,pH6.8。每盆定植3株,每個處理定植10盆,并進行正常肥水管理。移栽盆內1個月后,于6月初進行加氧灌溉處理,灌溉水采用普通地下水,其含氧量在(1±0.1)mg·L-1。試驗設3種處理(表1)。采用電磁式充氣泵(型號:ACO—002)對灌溉水進行通氣增氧,在101 kPa、25 ℃條件下,對灌溉水進行氣泵通氣增氧,4 L灌溉水中的含氧量(y)與通氣時間(x)的變化關系為:y=-0.002 5x2+ 0.207 2x+ 1.854 4(R2=0.902 5),并采用便攜溶解氧測定儀(型號:BDO-7500)對灌溉水的氧含量進行監測。
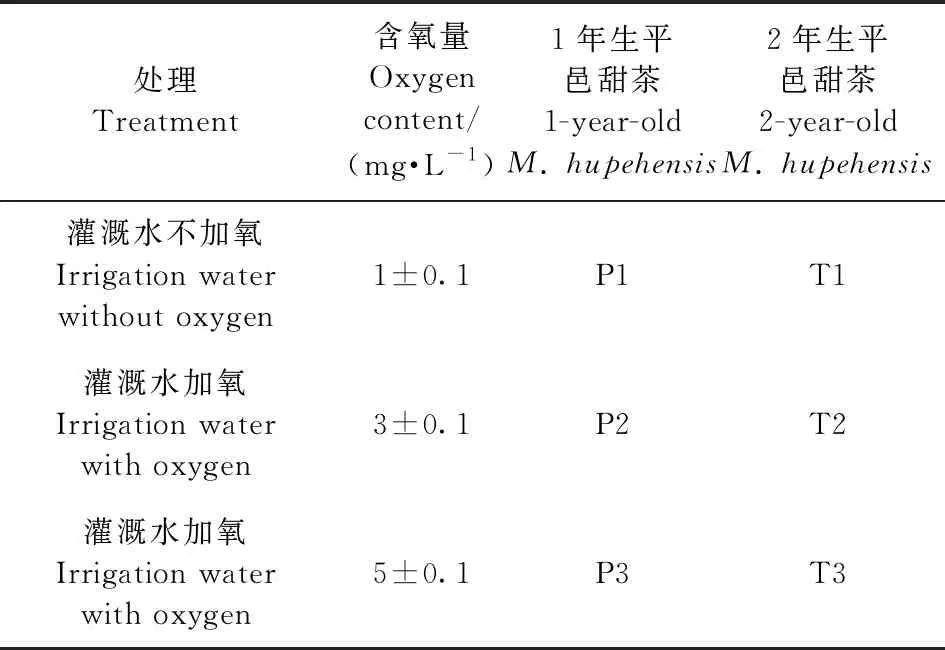
表1 不同處理灌溉水含氧量
1.2 試驗方法
1.2.1 生物量的測定 加氧處理后,選健壯幼苗做好標記,用米尺定期測定植株高度,用游標卡尺測定其地徑,每個處理測10棵。 將試驗植株從盆中取出帶入實驗室清洗,將植株鮮葉和莖放入烘箱中105 ℃殺青30 min,再調溫度至80 ℃烘24 h,稱量地上部干質量。
1.2.2 葉片葉綠素和光合熒光指標的測定 葉綠素相對含量(SPAD)用SPAD-502便攜式葉綠素儀測定,光合指標使用CIRAS-2型(PP-Systems)便攜式光合作用測定系統測定,熒光指標使用Junior-PAM(德國WAIZ)便攜式熒光儀測定。于7月14日、8月17 日、9月16日上午 10:00(晴朗的天氣),隨機對各個處理 3 株平邑甜茶進行測定,每株幼苗選擇頂端往下3~5 片舒展的生長良好的成齡葉,對葉片進行固定然后做好標記,測定時避開葉脈、葉片邊緣、病斑等部位以減少誤差。
1.2.3 葉片礦質營養的測定 2種年限平邑甜茶幼苗分別設置3個重復,每個重復采摘5片,共采摘15片,置于冰盒內帶回實驗室,對采集的葉片用去離子水沖洗干凈,再用吸水紙擦干葉片表面的水分后裝入信封,放入烘箱中105 ℃殺青20 min,80 ℃烘干后進行磨碎,過 60 目篩,裝入密封袋保存到干燥容器中,備用。
礦質元素的測定采用H2SO4-H2O2聯合消煮法。消煮后葉片全氮采用multiN/C3100測定; 葉片全磷采用鉬藍比色法測定; 葉片全鉀采用火焰光度法測定; 葉片鈣、鎂、鐵、鋅的含量采用原子吸收分光光度法測定(楊露等, 2020)。
1.2.4 根系抗氧化酶的測定 用氮藍四唑(NBT)光還原法對超氧化物歧化酶(SOD)的活性進行測定(陳貽竹等, 1988); 過氧化物酶(POD)的活性按照Omran(1980)的方法進行測定; 過氧化氫酶(CAT)的活性按照Singh(2010)的方法進行測定。
1.2.5 根系結構和活力的測定 將植株從花盆里小心取出,清水沖洗干凈,放置于測定盤上,打開Scan Makeri800 plus掃描儀,掃描獲取根系構型掃描圖像,用萬深LA-S系列植物圖像分析儀測定根系長度、表面積、根尖數等參數。植株根系活力采用TTC法(趙世杰等, 2002)測定。
1.2.6 土壤微細菌、微真菌、放線菌數量的測定 于2019 年7月14日、8月17日、9月16日取每個處理的土壤,參考華菊玲等(2012)的方法。細菌在37 ℃的培養箱內恒溫培養,放線菌和真菌培養箱溫度為28 ℃。通常細菌24 h后計數,放線菌5天后計數,真菌48 h后計數,用稀釋平板計數法進行測定(程麗娟等, 2000)。
1.2.7 土壤酶活性的測定 土壤蔗糖酶的測定采3,5-二硝基水楊酸比法;土壤脲酶的測定采用靛酚藍比色法;土壤磷酸酶的測定采用磷酸苯二鈉比色法(關松蔭, 1986)。
1.3 數據處理
采用 Excel 2003 處理試驗數據。通過 SPSS 19.0進行方差分析,采用鄧肯氏新復極差法進行差異顯著性檢測。
2 結果與分析
2.1 氣泵增氧對灌溉水中含氧量變化的影響
在101 kPa、25 ℃條件下,隨時間的增加,水中氧含量呈增加趨勢,且10 min內為快速增加時段,至30 min呈飽和狀態。灌溉水增氧效果較穩定(R2=0.902 5(圖1),灌溉水中的含氧量與通氣時間的關系可以表示為:y= -0.002 5x2+ 0.207 2x+ 1.854 4。
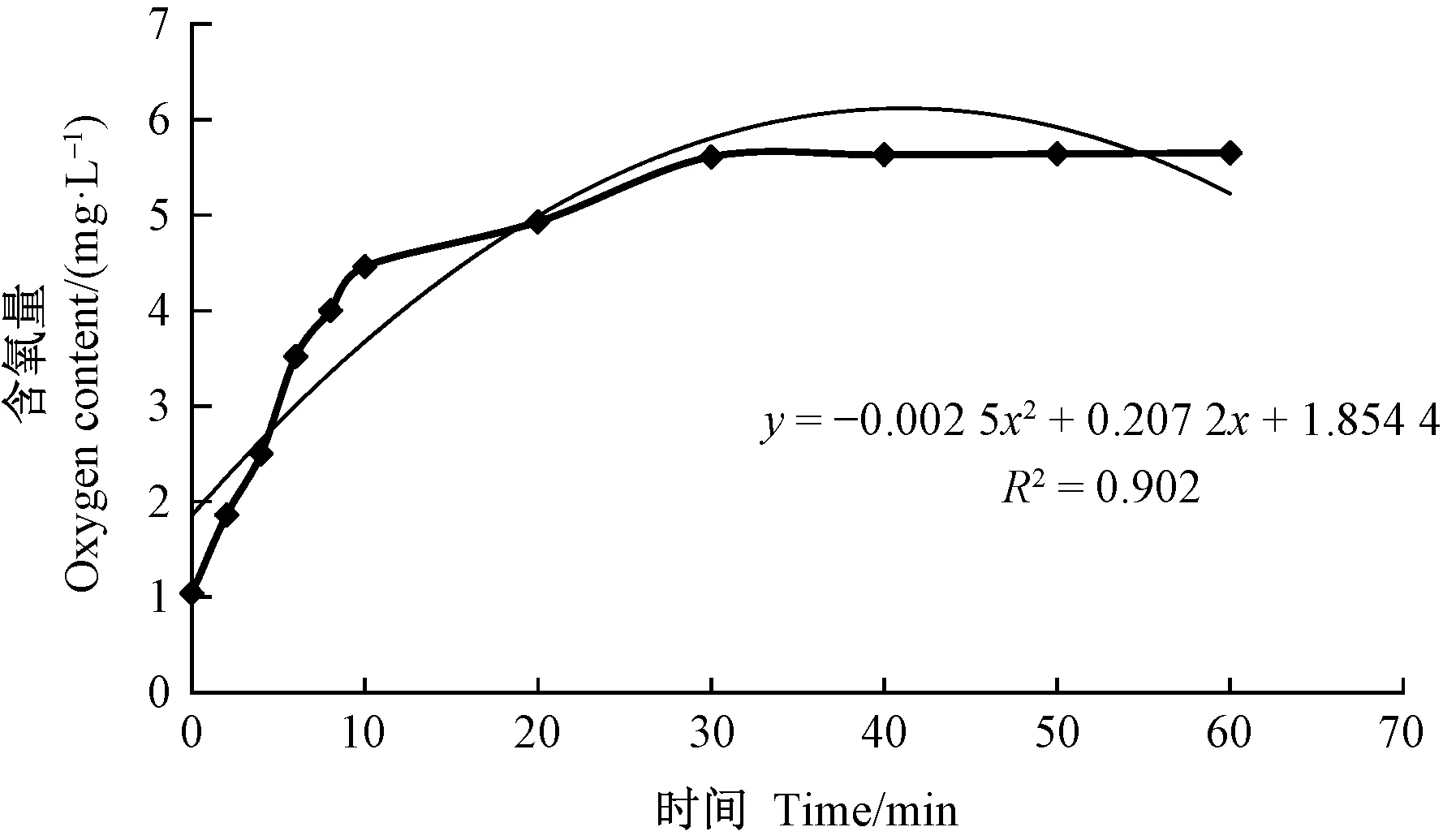
圖1 灌溉水中的含氧量隨通氣時間的變化
2.2 增氧灌溉對1、2年生平邑甜茶地上部生長的影響
采用增氧灌溉1個月后,盆栽平邑甜茶的株高、地徑以及地上部干質量均有顯著增加(表2)。1年生平邑甜茶株高8月份提高最顯著(P<0.05),2年生平邑甜茶中T3處理株高相較于T1在7月份的提高最為顯著; 1年生平邑甜茶處理地經在8月份提高最為顯著,2年生平邑甜茶中T3處理地徑在9月份相比T1提高最為明顯; 1年生平邑甜茶地上部干質量7月份提高最為顯著,2年生平邑甜茶在地上部干質量9月份提高最為顯著。由此可知,澆灌水中的含氧量與株高的增長量、干質量呈正比。

表2 增氧灌溉對1、2年生平邑甜茶地上部生長的影響
2.3 增氧灌溉對1、2年生平邑甜茶相對葉綠素含量的影響
3種不同含氧量灌溉水處理的葉綠素含量之間差異顯著(表3)。1年生平邑甜茶在8月份提高最顯著,2年生平邑甜茶中,除7月份無明顯差異外,8、9月均有顯著差異,且T3處理相比T1在8月份提高9.9%,最為顯著。
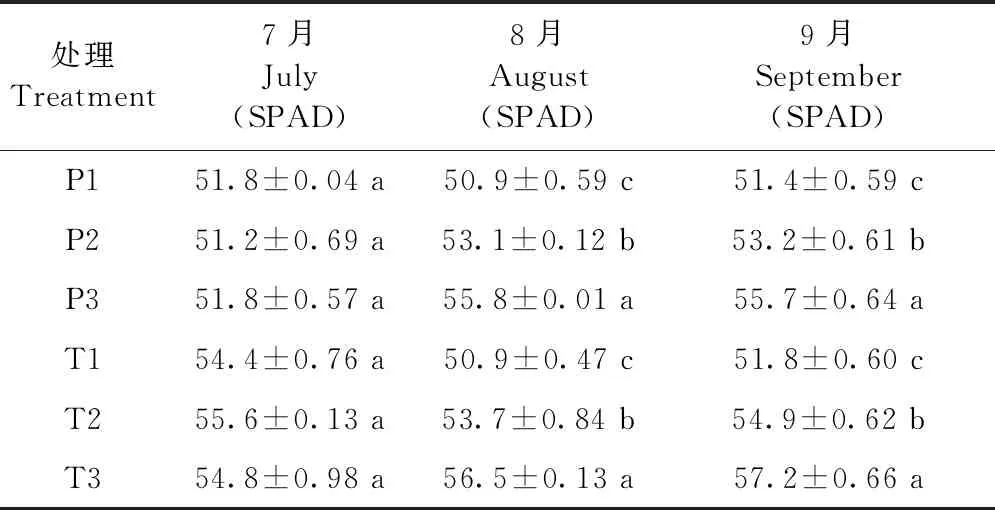
表3 增氧灌溉對1、2年生平邑甜茶相對葉綠素含量的影響
2.4 增氧灌溉對1、2年生平邑甜茶葉片凈光合速率的影響
增氧灌溉1個月后,P3、P2處理的葉片凈光合速率差異不顯著,T1、T2處理的葉片凈光合速率差異不顯著,但P3、T3處理相比P1、T1均有明顯提升; 1、2年生平邑甜茶葉片凈光合速率均在8月份提升最為顯著,P3處理的葉片凈光合速率相比P1提高了128.5%,T3處理的葉片凈光合速率相比T1提高18.0%(表4)。
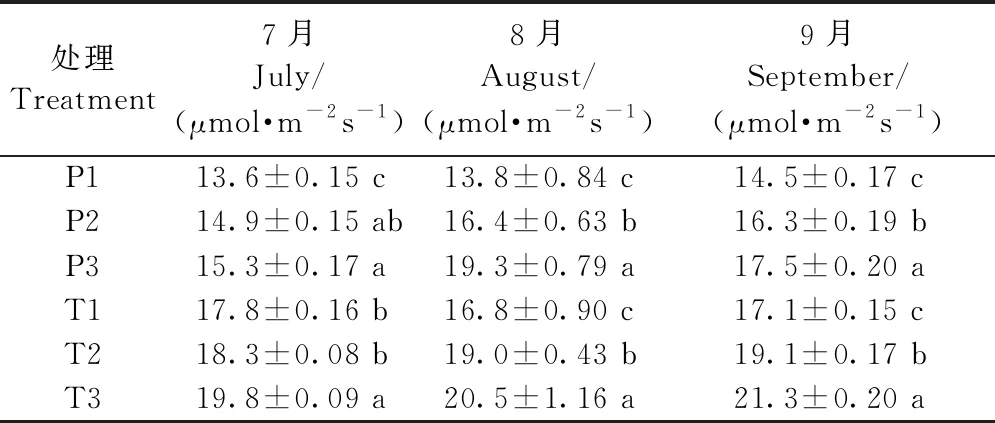
表4 增氧灌溉對1、2年生平邑甜茶葉片凈光合速率的影響
2.5 增氧灌溉對1、2年生平邑甜茶葉片最大光化學效率(Fv/Fm)的影響
7、8月份,2年生平邑甜茶3種處理之間Fv/Fm差異不顯著,但8月份T3處理的葉片Fv/Fm顯著高于T1、T2,1年生平邑甜茶3種處理葉片的Fv/Fm,P2處理在7月份提高最為顯著,相比P1提高了8.3%;9月份,P3處理的葉片Fv/Fm相比P1提高14.1%,最為顯著,T2、T3處理的葉片Fv/Fm相比T1,分別提高了4.7%、8.9%(表5)。
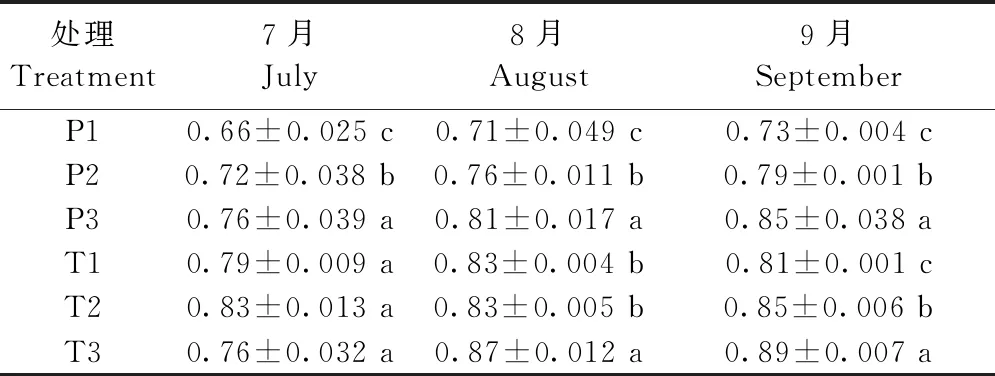
表5 增氧灌溉對1、2年生平邑甜茶葉片Fv/Fm的影響
2.6 增氧灌溉對1、2年生平邑甜茶根系結構的影響
1、2年生平邑甜茶根系長度均在8月份提升最為顯著(圖2),分別提高30.3%、32.1%; P2、P3處理的根系根尖數7月份差異不顯著,但在9月份提升最明顯,2年生平邑甜茶3種不同處理中,以8月份提升最明顯(圖3); 1、2年生平邑甜茶根系表面積均在9月份提升最明顯(圖4); 1年生平邑甜茶根系呼吸速率在7月份提升最顯著,而2年生平邑甜茶在8月份提升最顯著,T3處理的根系呼吸速率相比T1顯著增加31.9%(圖5)。灌溉水中含氧量的變化與地下部根系長度、表面積、根尖數呈成正比,且隨著時間的延長,效果越明顯。
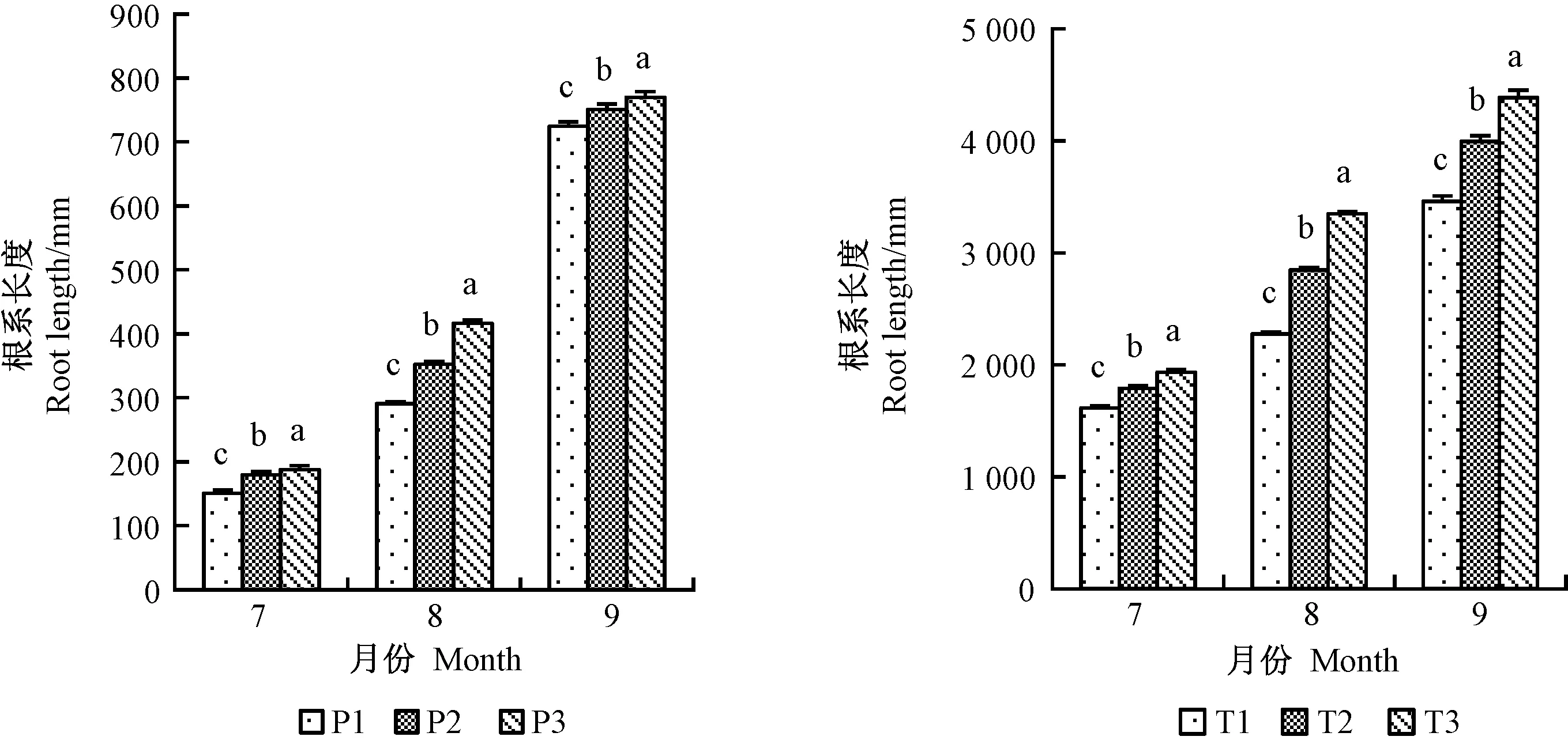
圖2 增氧灌溉對1、2年生平邑甜茶根系長度的影響
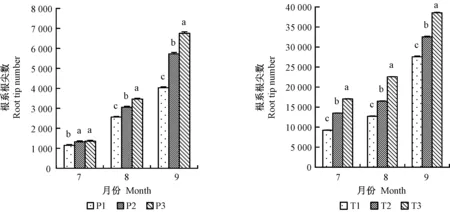
圖3 增氧灌溉對1、2年生平邑甜茶根系根尖數的影響
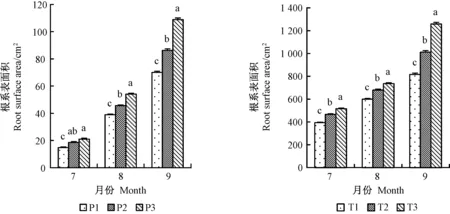
圖4 增氧灌溉對1、2年生平邑甜茶根系表面積的影響
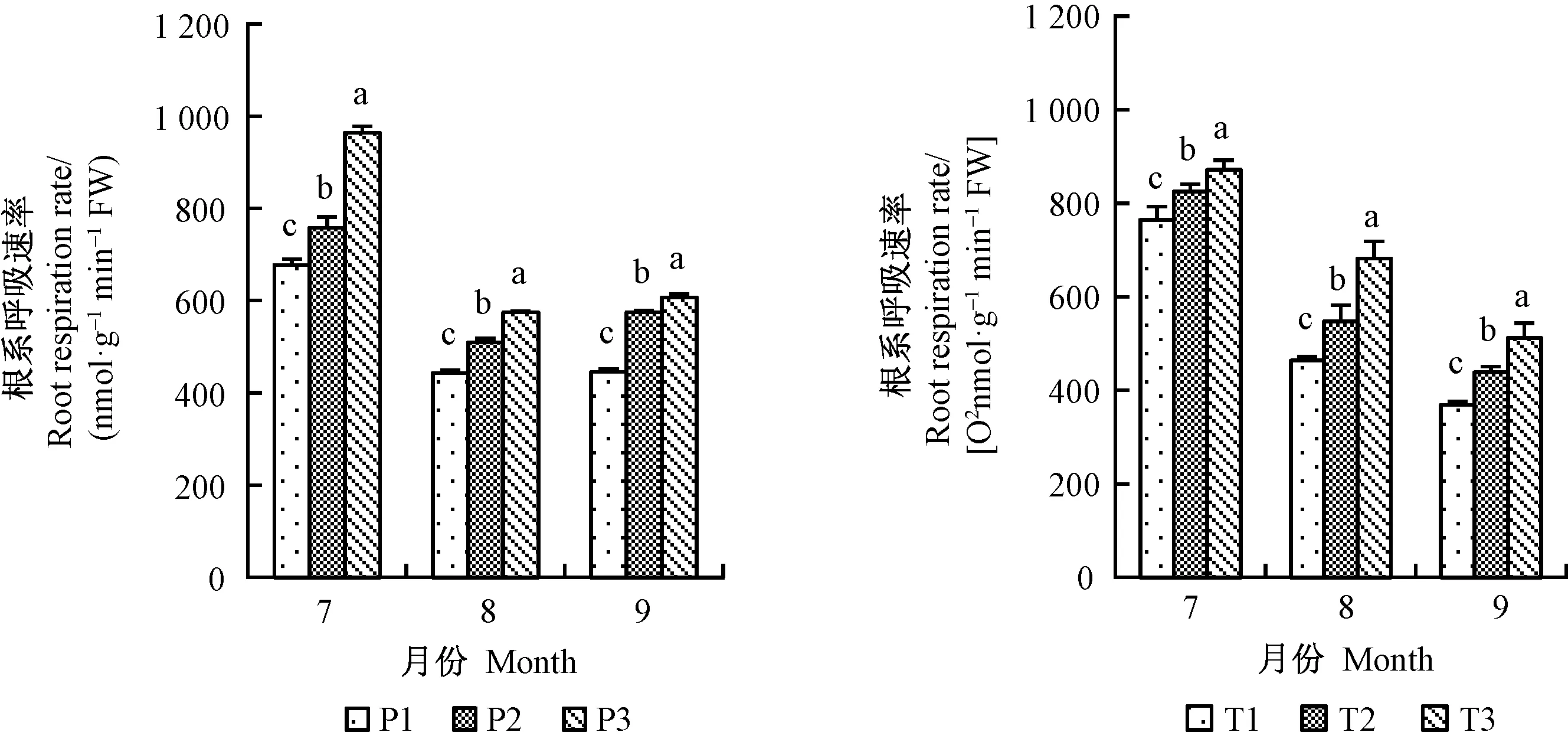
圖5 增氧灌溉對1、2年生平邑甜茶根系呼吸速率的影響
2.7 增氧灌溉對1、2年生平邑甜茶根系抗氧化酶活性的影響
增氧灌溉均可不同程度提高平邑甜茶的根部SOD、POD和CAT酶活性。7月份,1年生平邑甜茶P1、P2和T2、T3處理的SOD活性之間差異不顯著,且P3處理的SOD活性顯著高于P1、P2;8、9月份,各處理之間的根酶SOD活性均差異顯著,SOD活性在8月份均達到最高(圖6)。 7、8月份P1、P2、P3 3種處理的CAT活性差異顯著,且以7月份提升最明顯,2年生平邑甜茶8月份提高最顯著,P3處理9月份CAT活性遠高于P1和P2(圖7)。P3處理的POD活性8月份提升最為明顯,相比P1提高30.7%,T2、T3處理雖在7月份的POD活性差異不顯著,但相對于其他兩月,T3處理相比T1處理的POD活性提高最明顯,提高27.2%(圖8)。以上結果表明灌溉水中氧氣含量的增加提高根部酶活性,并促進根系生存發育。。
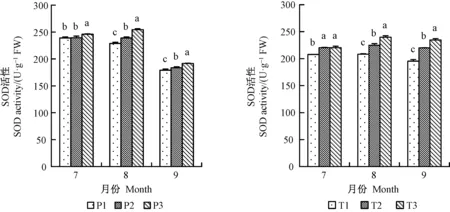
圖6 增氧灌溉對1、2年生平邑甜茶根酶SOD活性的影響
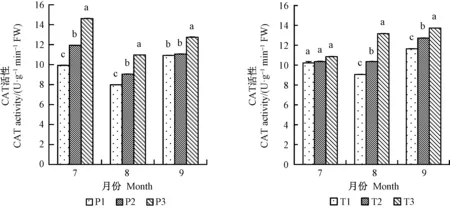
圖7 增氧灌溉對1、2年生平邑甜茶根酶CAT活性的影響
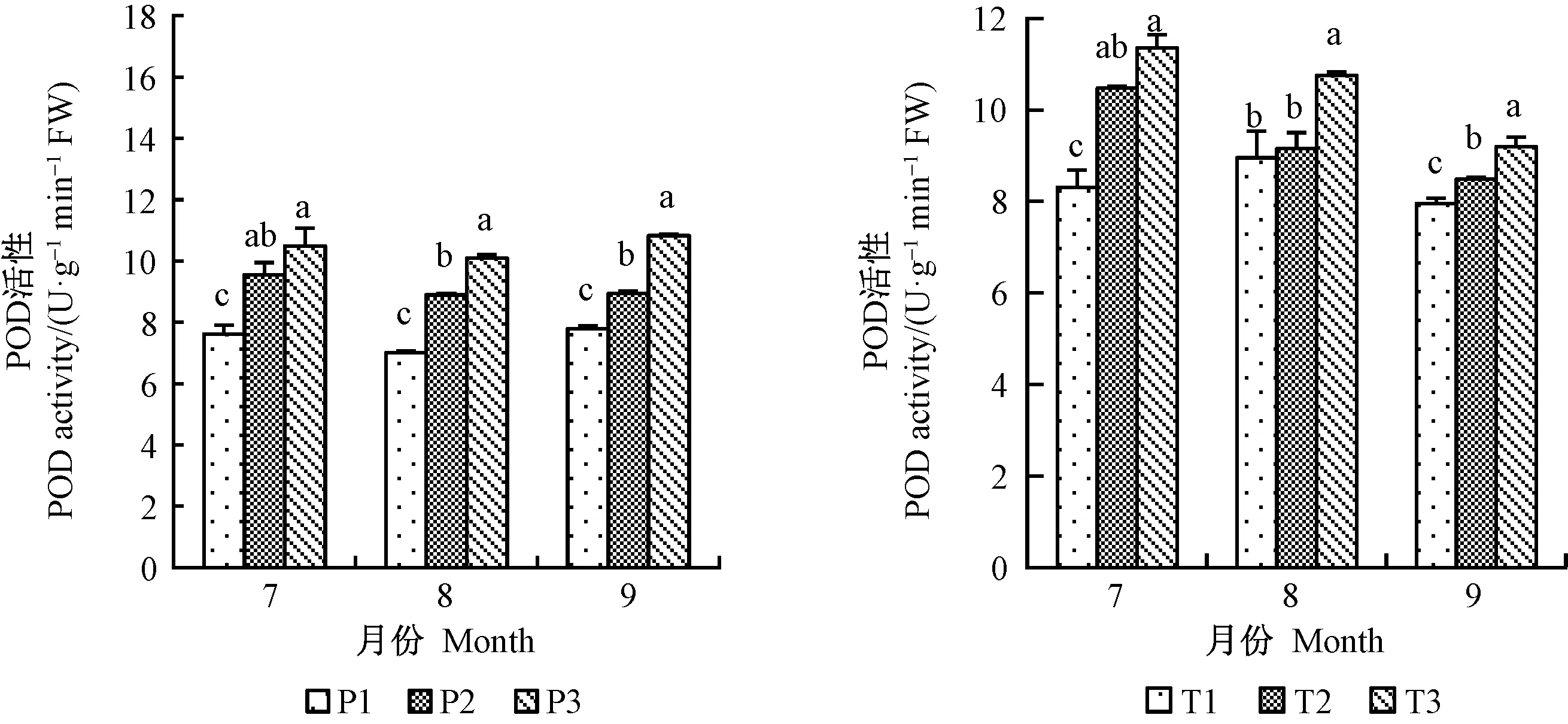
圖8 增氧灌溉對1、2年生平邑甜茶根酶POD活性的影響
2.8 增氧灌溉對1、2年生平邑甜茶葉片礦質營養的影響
由表6可知,3種不同含氧量的灌溉水澆灌盆栽平邑甜茶,礦質元素含量差異顯著。 氮、磷、鉀含量變化趨勢相同,灌溉水中的含氧量越高,葉片中氮、磷、鉀的含量越高; P2、P3處理的葉片鎂含量相比P1分別顯著提高15.8%、27.5%,T3處理相比T1葉片鎂含量顯著提高11.9%; 鐵含量2組處理差異均顯著,P2、P3處理的葉片鐵含量相比P1分別提高12.2%、29.5%,T2、T3處理的葉片鐵含量相比T1分別提高14.2%、24.6%; P2、P3處理的葉片鈣含量相比P1分別提高29.5%、43.8%, 鋅含量相比P1分別提高4.7%、8.5%,T3處理葉片鈣含量相比T1顯著提高12.5%,鋅含量T3處理相比T1顯著提高5.2%。因此,增氧灌溉可增加植物葉片礦質元素的積累,并促進植株生長。

表6 增氧灌溉對1、2年生平邑甜茶葉片礦質營養的影響
2.9 增氧灌溉對1、2年生平邑甜茶土壤生理生化的影響
2.9.1 增氧灌溉對1、2年生平邑甜茶土壤酶活性的影響 盆栽平邑甜茶所處土壤中,土壤脲酶、磷酸酶和蔗糖酶的活性最高。7月份1年生平邑甜茶土壤脲酶活性P2、P3差異不顯著,但明顯高于P1,且在8月份提升最顯著,P3處理的土壤脲酶活性相比P1顯著提高15.4%;8月份2年生平邑甜茶提升最顯著;9月份T1、T2處理的土壤脲酶活性差異不顯著,且活性低于前2個月,排除試驗誤差的影響,土壤脲酶活性與灌溉水含氧量呈正比(圖9)。1年生平邑甜茶的土壤磷酸酶活性在8月份提升最為顯著,P3處理相比P1顯著提高了26.8%,2年生平邑甜茶土壤磷酸酶活性在9月份提升最顯著(圖10)。 1、2年生平邑甜茶間土壤蔗糖酶活性差異顯著,且均有顯著的提高(圖11)。因此,根區通氣有利于根系進行有氧呼吸,提高植株的根系活力,根系分泌物增多,并提高土壤酶活性。
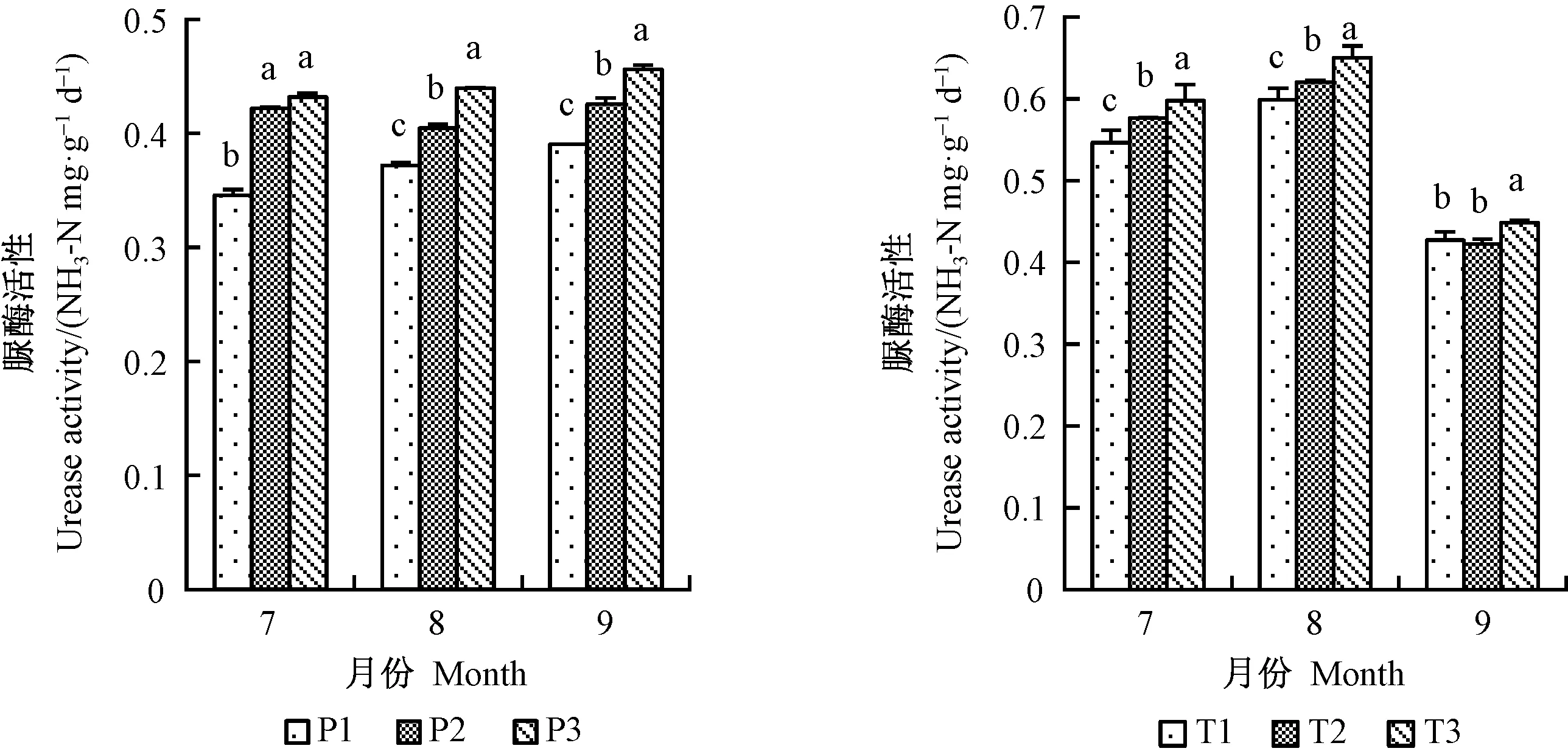
圖9 增氧灌溉對1、2年生平邑甜茶土壤脲酶活性的影響

圖10 增氧灌溉對1、2年生平邑甜茶土壤磷酸酶活性的影響
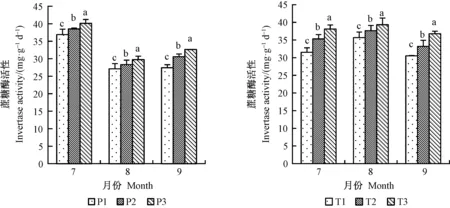
圖11 增氧灌溉對1、2年生平邑甜茶土壤蔗糖酶活性的影響
2.9.2 增氧灌溉對1、2年生平邑甜茶土壤微細菌、微真菌、放線菌數量的影響 提高灌溉水中的含氧量均可不同程度地提高植株根區土壤細菌和放線菌的數量、降低植株根區真菌的數量。采用3種含氧量的灌溉水澆灌盆栽平邑甜茶1個月后,其土壤微細菌數量之間均差異顯著(表7),1年生平邑甜茶在7月份改善效果最佳,2年生平邑甜茶在8月份改善效果最佳。P3、T3處理的土壤微真菌數量均顯著低于P1、T1,其中,1年生平邑甜茶在8月份差異最顯著,P3處理的土壤微真菌數量相比P1顯著降低35.5%,2年生平邑甜茶在9月份差異最顯著,T3處理相比T1顯著降低39.9%(表8)。P3處理的土壤放線菌數量在9月份相比P1提升最顯著,而T3處理的土壤放線菌數量則在8月份相比T1提升最顯著(表9)。
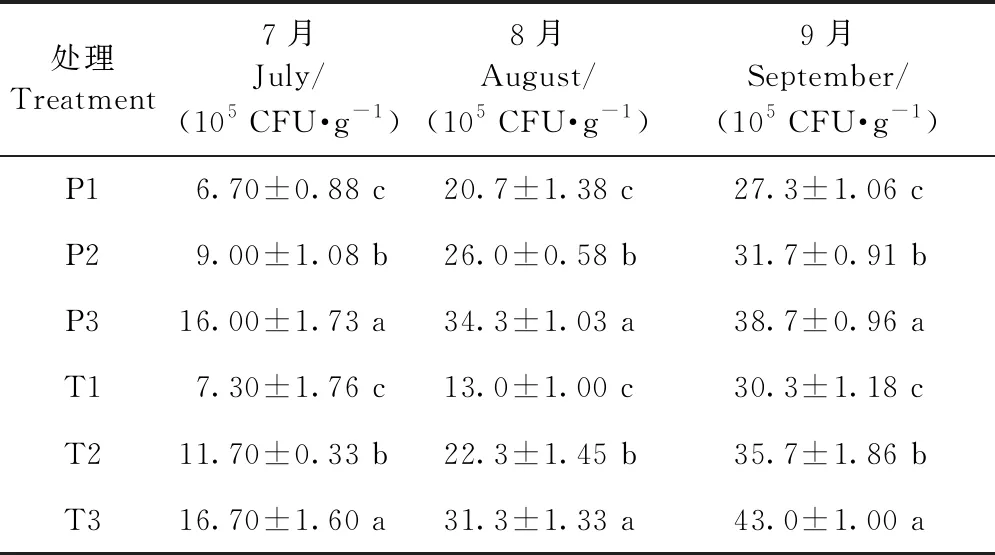
表7 增氧灌溉對1、2年生平邑甜茶土壤微細菌數量的影響
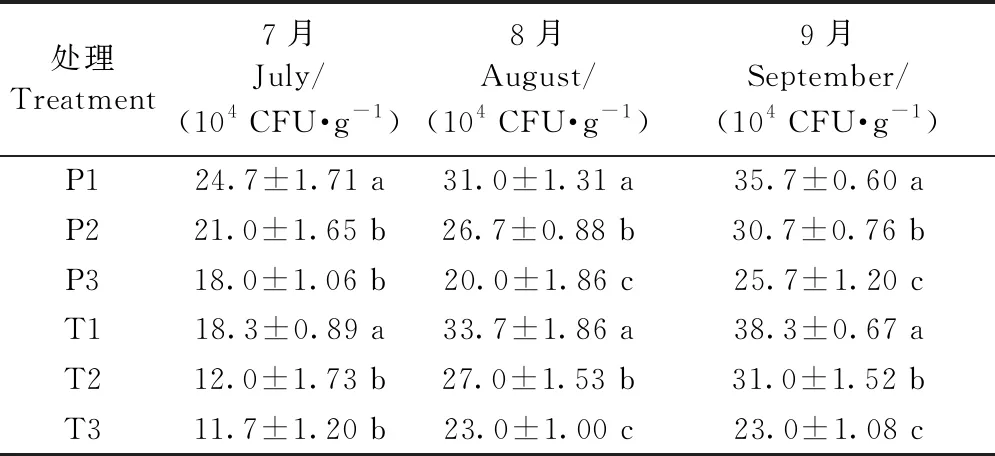
表8 增氧灌溉對1、2年生平邑甜茶土壤微真菌數量的影響
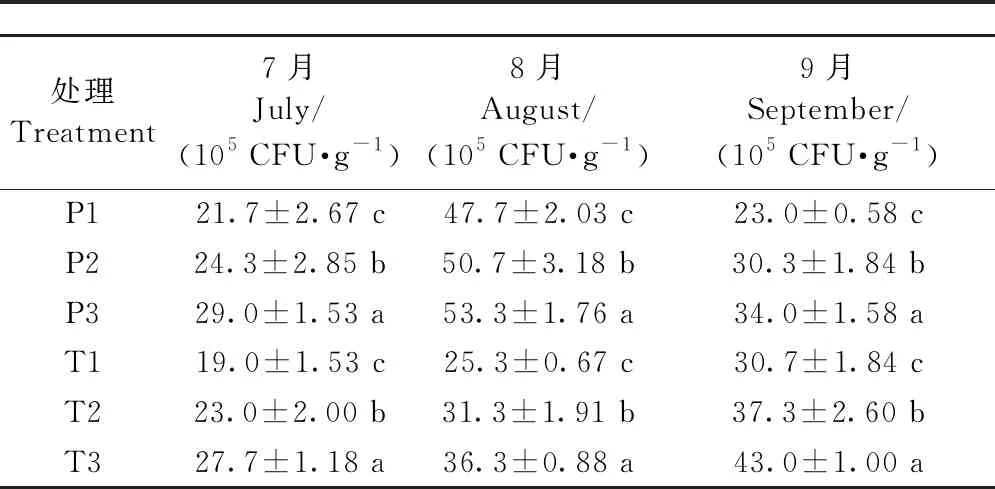
表9 增氧灌溉對1、2年生平邑甜茶土壤微放線菌數量的影響
3 討論
3.1 增氧灌溉對平邑甜茶地上部生長的影響
植物根部氧環境的調節影響植株地上部分的生長狀況(趙鋒等, 2010),在蘋果和桃(Amygdaluspersica)樹的研究中發現,葉片光合速率與葉片干質量有關(Angelaetal., 2018)。與常規灌水相比,對甜瓜(Cucumismelo)進行增氧灌溉處理能顯著促進其根系對水分的吸收,增加莖粗(張敏等, 2010); 與正常滴灌相比,不同濃度的灌溉水增氧處理,有利于植物根系對水分的吸收利用,影響植物株高(劉義玲等, 2010)。本試驗中,澆灌水含氧量與株高、地徑、地上部干質量成正比,各項數據表明,隨含氧量的增加,各處理總體均為上升趨勢, 其中含氧量(5±0.1) mg·L-1的處理效果最佳。
光合作用是植物生長發育的基礎,而光合色素是植物進行光合作用的基礎(夏慶平等, 2011)。SPAD作為葉綠素相對含量與葉綠素實際含量具有顯著的正相關關系,可在一定程度上反映蘋果葉片中真實葉綠素含量(Uddlingetal., 2007),而葉綠素含量在一定程度上與葉片的凈光合速率呈正相關關系。熒光特性對植物光合作用中的光能吸收、傳遞等過程具有重要意義(李中勇等, 2013),植物葉片的PSⅡ作為光能捕獲和傳遞的活性中心,其活性的正常與否直接影響著光合原初反應的進行(呂三三等, 2015)。因此植株長勢、葉綠素含量和光合熒光性能關系密切(高付鳳等, 2017)。本研究中,8、9月份時,灌溉水中的含氧量與2種砧木葉片的葉綠素含量和葉片凈光合速率成正比; 并與葉片Fv/Fm的反應趨勢基本相同。植物營養是生長的基礎,進行增氧灌溉,灌溉水將氧氣輸送到地下,改善幼苗生長的根際環境,促進根系對水分的吸收,提高地上部植株葉片的光合性能與熒光特性,光合作用積累更多的有機物,進而促進幼苗的生長發育和干物質的積累。
3.2 增氧灌溉對平邑甜茶葉片礦質營養的影響
礦質元素在植物體內物質組成和代謝過程中都起著重要的作用,植株葉片是樹體反映礦質營養元素含量最為敏感的器官之一,除了大量營養元素之外,植物還需要鈣、鐵、鋅、鎂等中微量營養元素,鈣對植物的細胞壁和組織具有重要的穩定作用,它可與中膠層果膠質形成果膠酸鈣而被固定,也能將質膜上的磷脂分子聯合起來,從而增強質膜的選擇性和疏水性(劉秀春, 2004)。鐵與葉綠體結構的形成有密切的關系,在植物新陳代謝等生理活動中也起著相當重要的作用(金崇偉, 2008)。鋅是一些轉錄調節蛋白的重要組成成分,因此,植物缺鋅往往會造成 RNA 轉錄受阻,繼而造成氨基酸含量高但蛋白質含量低的現象。鎂是葉綠素的組成成分,能夠促進營養物質積極參與光合作用,鎂還有助于磷酸鹽的代謝,植物的呼吸和許多酶系統的活化。因此可以利用葉片分析營養診斷來反映樹體對土壤中礦質元素的吸收狀況(宋曉暉等, 2011; Rouphaeletal., 2017)。本研究結果表明,灌溉水中含氧量越高,葉片中氮、磷、鉀的含量越高,且灌溉水中的含氧量與平邑甜茶葉片中鈣、鐵、鋅、鎂元素含量基本呈正比。通過葉片礦質元素分析表明增氧灌溉的效果顯著,綜合來看,含氧量為(5±0.1) mg·L-1的灌溉水處理效果最佳。
3.3 增氧灌溉對平邑甜茶地下部根系的影響
根系是果樹栽培的基礎,根系具有支撐、固定、吸收和貯藏營養物質等作用(鄧蘭生等, 2007),通過吸收水分、礦質養分以及合成內源激素等方法影響地上部的生長發育(楊洪強等, 2007)。本研究結果表明,灌溉水中含氧量的改善與地下部根系長度、表面積、根尖數的變化成正比,且隨著時間的延長,效果越明顯。這可能是因為灌溉水中含氧量增加,植物根際環境得到改善,根系呼吸作用加強,促進根系生長,增強了根系的吸收能力,有利于根系有效吸收土壤中的養分和水分。
根系呼吸速率作為根系發育與吸收功能的指標,可反映植物新陳代謝的強弱(王柯等, 2019)。本試驗中,提高灌溉水中的含氧量均可顯著改善其根系呼吸速率。果園在進行土壤漫灌時,由于土壤中的氣體被灌溉水趕出,使得土壤中暫時缺氧,導致植物根系呼吸困難,影響根系對營養元素和水分的吸收和利用(Srivastavaetal., 2014)。但是在進行增氧灌溉時,灌溉水可以將氧氣輸送到地下根部周圍,明顯改善了土壤氧含量的下降,促進了植株根系生長,根系呼吸作用增強,提高了土壤氧的有效性(Zhuetal., 2019),加快植株生長。
當植株生長受到一定程度的抑制時,活性氧大量產生,打破植物體內的抗氧化酶系統與活性氧之間的動態平衡,多余的活性氧對植物造成傷害,這些變化可以通過酶活性的高低來體現(李林懋等, 2014)。SOD、POD、CAT 等抗氧化酶可以保護植物體不受過量自由基的傷害,其活性的高低能夠反映逆境脅迫對植株的傷害程度(Ahmadetal., 2010)。本研究中,增加灌溉水的含氧量,其抗氧化酶系統做出相應的反應,可以抵御外界不良條件侵害,均可不同程度地提高砧木的SOD、POD和CAT活性,從而降低體內活性氧的數量,減少活性氧過量積累對植株造成的損傷,促進植物根系健康生長,提高自身抗逆性。
3.4 增氧灌溉對平邑甜茶土壤根際環境的影響
土壤根際環境可以影響土壤酶的活性。蔗糖酶活性的強度反映了底物的成熟程度和生育水平,在樹體的營養生長中起著重要作用; 脲酶是對尿素轉化起關鍵作用的酶,其酶促反應產物是可供植物利用的氮源,其活性可以用來顯示土壤的氮供應能力(許云翔等, 2019); 磷酸酶可以促進有機磷化合物或無機磷酸鹽轉化為植物可利用的無機磷,是衡量土壤肥力的重要指標。根區通氣為植株根系的生長創造良好的生長環境,有利于根系有氧呼吸,提高植株的根系活力(趙旭等, 2010),同時,增氧灌溉也可顯著提高脲酶和過氧化氫酶的平均活性(Duetal., 2020),根系分泌物增多,土壤酶活性也隨之提高。本研究中,當灌溉水含量為(5±0.1)mg·L-1時,土壤脲酶、磷酸酶和蔗糖酶的活性最高,3個月中,含氧量均與土壤酶活性呈正比,但在9月份,2年生盆栽平邑甜茶的土壤脲酶活性較低,且3種處理間差異不顯著,這可能是因為9月份土壤水分蒸發較慢,灌溉次數減少所導致。
土壤微生物是土壤生態系統的重要組成部分,是土壤健康和質量的敏感指標(Danieletal., 2017),在植物殘體降解、營養轉化周期等方面起著重要作用(Schutteretal., 2001)。土壤微生物具有多功能活性,特別是植物生長促進和通過增加對資源的競爭、化合物的分泌和病原體的直接寄生來抑制土傳病原體(Mirzaetal., 2020)。土壤環境長期缺氧會導致土壤的理化和生物特性發生改變,土壤微生物區系也發生明顯變化(Guoetal., 2014),植物主要表現為株高降低、葉面積減小、葉綠素含量降低、光合速率下降等。本研究結果表明: 提高灌溉水的含氧量均可不同程度提高植株根區土壤細菌和放線菌數量、降低植株根區土壤真菌數量。根系活力的提高以及根系分泌物的增多促進了微生物的繁衍,土壤細菌與放線菌數量由此增加,土壤含水量是影響真菌群落分布主要環境因子(曹紅雨等, 2019),多次的灌水處理可能導致了真菌數量的明顯下降。因此,通過增氧灌溉改善植物根區土壤氧氣狀況,滿足植物對水、肥、氣的需求,某些病原菌微生物數量急劇減少,而有益微生物增加,有利于土壤有機物質的分解,促進植株生長。
4 結論
增氧灌溉提高了根區的溶解氧含量,促進根系的呼吸作用,增大根系的分布范圍,加快根系的代謝活動,使得土壤酶的活性增加,并提高土壤微生物活性,改善植株的礦質營養含量,最終有利于株高和地徑的增長,改善葉綠素含量、光合熒光指標,促進有機物質的積累。
本試驗結果表明,灌溉水中的含氧量在(1±0.1)~(5±0.1) mg·L-1的范圍內,含氧量越高,對改善盆栽平邑甜茶生長、根區土壤環境以及土壤微生物結構的效果越明顯。
