大柴胡湯治療肝細胞癌lncRNA-miRNA-mRNA轉錄網絡整合分析
2021-12-15 00:25:38鄭飄陳博威寧迪敏肖唯田雪飛
中國中醫藥信息雜志 2021年12期
鄭飄,陳博威,寧迪敏,肖唯,田雪飛
大柴胡湯治療肝細胞癌lncRNA-miRNA-mRNA轉錄網絡整合分析
鄭飄1,陳博威2,寧迪敏1,肖唯3,田雪飛1
1.湖南中醫藥大學中西醫結合學院,湖南 長沙 410208;2.湖南中醫藥大學第一附屬醫院,湖南 長沙 410007;3.中南大學湘雅醫院中西醫結合研究所,湖南 長沙 410008
挖掘大柴胡湯治療肝細胞癌的lncRNA-miRNA-mRNA調控網絡,探討其抗肝癌作用機制。利用中藥系統藥理學數據庫與分析平臺(TCMSP)、GEO、miRDB、miRTarBase、TargetScan數據庫獲取大柴胡湯的活性成分、預測靶點及肝細胞癌的差異表達長鏈非編碼RNA(lncRNAs)、微小RNA(miRNAs)和mRNAs,通過R語言和Perl軟件篩選大柴胡湯治療肝細胞癌的關鍵lncRNAs、miRNAs、mRNAs。采用Cytoscape3.7.0軟件和STRING數據庫構建活性成分-靶點網絡、lncRNA-miRNA-mRNA轉錄網絡和蛋白相互作用網絡,進行GO和KEGG富集分析,并予分子對接驗證,進行整合分析。得到大柴胡湯治療肝細胞癌的97個有效成分和42個靶點,構建了由19個關鍵lncRNAs、23個關鍵miRNAs和42個關鍵mRNAs組成的lncRNA-miRNA-mRNA轉錄網絡。富集分析結果顯示,這些靶基因參與了腫瘤的生存、凋亡、自噬、氧化應激、蛋白激酶等生物學過程,主要通過PI3K-Akt信號通路、MAPK信號通路和TNF信號通路等發揮作用。大柴胡湯治療肝細胞癌具有多成分、多靶點、多途徑協同作用特點,可為相關研究提供依據和方向。
肝細胞癌;大柴胡湯;轉錄網絡;生物信息學;網絡藥理學;分子對接
肝細胞癌(hepatocellular carcinoma,HCC)是原發性肝癌最主要的組織學亞型,具有侵襲和轉移率高、復發風險高、中晚期并發癥嚴重等特點。目前,其臨床首選方法為手術、放療和化療。盡管靶向藥物的新發展為肝癌治療帶來希望,但價格昂貴、反應率低、耐藥性頻繁,導致其臨床應用局限。大柴胡湯出自《傷寒論》,由柴胡、大黃、枳實、黃芩、半夏、白芍、大棗和生姜組成,有疏肝理氣之功。臨床研究表明,大柴胡湯對HCC表現出良好療效,可增強HCC患者機體免疫力,減少并發癥的發生發展[1-2]。實驗研究顯示,大柴胡湯可促進肝癌HepG2細胞凋亡、抑制肝纖維化和調節代謝等[3-5]。然而,以往研究主要集中于單一靶點和單一機制,未能基于中醫整體觀念系統探討大柴胡湯治療HCC的分子機制。
在HCC的病理及治療研究中,由長鏈非編碼RNA(lncRNA)、微小RNA(miRNA)和mRNA組成的轉錄網絡逐漸受到關注,lncRNA在轉錄后水平上作為競爭性內源RNA,通過miRNA的特異性海綿吸附間接降低miRNA與下游靶基因的結合,從而實現基因調控[6]。研究發現,lncRNA和miRNA在HCC的發生和發展中起著至關重要的作用,其廣泛參與免疫抑制、氧化應激、血管生成、增殖、轉移和細胞代謝等生物過程,lncRNA、miRNA和mRNA的交聯已成為與HCC進展相關的新興課題[7-9]。中藥及其有效成分對lncRNA、mRNA和miRNA表達的調控也逐漸成為研究熱點,現已發現多種中藥及有效成分在腫瘤轉錄調控中的作用,如柴胡有效成分柴胡皂苷D介導腫瘤細胞凋亡和多耐藥逆轉效應可能通過lncRNA的調控實現[10],大黃活性成分大黃素甲醚通過調節miR-370誘導肝癌細胞凋亡[11],黃芩有效成分黃芩素對肝癌HepG2細胞增殖和凋亡的作用與miR-34a密切相關[12]。基于以往研究,筆者利用生物信息學方法挖掘大柴胡湯治療HCC的活性成分-靶點網絡并構建lncRNA-miRNA-mRNA轉錄網絡,以期闡明大柴胡湯多組分、多靶點、多途徑協同作用的分子機制,并為后續基礎和臨床研究提供依據。
1 資料與方法
1.1 活性成分篩選和靶點預測
通過中藥系統藥理學數據庫與分析平臺(TCMSP,http://tcmspw.com/tcmsp.php)獲取大柴胡湯組方藥物柴胡、大黃、枳實、黃芩、半夏、白芍、大棗和生姜的有效成分。根據藥物的藥代動力學特征,以口服生物利用度(OB)≥30%和類藥性(DL)≥0.18為條件進行活性成分篩選[13],通過該數據庫查詢活性成分的潛在靶點,通過Perl(https://www.perl.org)將靶點導入UniProt數據庫(http://www.uniprot.org),獲得目標基因官方名稱。
1.2 差異表達lncRNAs、miRNAs和mRNAs挖掘
HCC基因表達譜原始數據(GSE101728)來源于基因表達綜合數據庫GEO(https://www.ncbi.nlm.nih. gov/geo)。該數據的人體組織樣本來自復旦大學中山醫院,包含HCC患者的7對腫瘤組織和癌旁組織(基因芯片平臺:GPL21047)。采用Perl和R4.0軟件篩選差異表達的lncRNA,閾值為|log2(FC)|>1.5和<0.05[14],進而通過Mircode數據庫(http://www. mircode.org)篩選與lncRNA結合的miRNAs。將獲得的miRNAs通過miRDB(http://mirdb.org)、miRTarBase(http://mirtarbase.mbc.nctu.edu.tw)和TargetScan(http://www.targetscan.org)數據庫進行映射,最終得出由miRNA調控的mRNAs。
1.3 活性成分-靶點和lncRNA-miRNA-mRNA相互作用網絡構建
采用Perl將HCC差異表達基因與大柴胡湯潛在靶點取交集,獲得可能參與大柴胡湯治療HCC的活性成分和關鍵lncRNAs、miRNAs、mRNAs。采用Cytoscape3.7.0構建lncRNA-miRNA-mRNA的成分-靶點網絡和三元轉錄網絡并進行可視化。采用STRING數據庫(https://string-db.org/)構建關鍵mRNAs蛋白相互作用(PPI)網絡并進行可視化,設置物種為“homo sapiens”,最小相互作用得分為0.4。
1.4 靶點功能富集分析
使用R軟件Biocmanager和ClusterProfiler對獲得的關鍵靶點進行GO和KEGG富集分析。GO對基因產物可能的生物過程(BP)、細胞組分(CC)及分子功能(MF)進行描述,KEGG對基因組注釋信息進行分類,從而得出最顯著的生物學過程。
1.5 分子對接分析
從TCMSP獲得大柴胡湯主要活性成分的三維結構作為配體,從RCSB PDB數據庫(https://www.rcsb. org/)獲得關鍵靶點的三維結構作為受體。使用AutodockTools1.5.6對受體和配體進行能量最小化,并轉換為PDBQT格式,獲得用于分子對接模擬的三維網格盒,使用AutoDock Vina 1.1.2進行分子對接,采用PyMOL(https://www.pymol.org/)進行可視化。
2 結果
2.1 大柴胡湯活性成分及靶點
通過TCMSP數據庫獲得大柴胡湯121個活性成分,其中柴胡12個、大黃7個、枳實18個、黃芩28個、半夏26個、白芍8個、大棗18個、生姜4個,去除重復成分后,共篩選出108個候選活性成分。主要活性成分信息見表1。共檢索到1 438個靶點,合并重復項后,獲得295個靶點。
表1 大柴胡湯主要活性成分信息
編號名稱OB/%DL靶點數來源 編號名稱OB/%DL靶點數來源 MOL000098quercetin(槲皮素)46.430.28141柴胡,大棗 MOL007879tetramethoxyluteolin(四甲氧基丁醇)43.680.3727枳實 MOL000422kaempferol(山柰酚)41.880.2456柴胡,白芍 MOL000173wogonin(漢黃芩素)30.680.2342黃芩 MOL000358beta-sitosterol(β-谷甾醇)36.910.7528大黃,大棗,半夏, MOL002714baicalein(黃芩素)33.520.2134黃芩 黃芩,生姜,白芍 MOL002928oroxylin a(木蝴蝶素)41.370.2323黃芩 MOL000449stigmasterol(豆甾醇)43.830.7627柴胡,大棗,半夏, MOL001689acacetin(金合歡素)34.970.2423黃芩 黃芩,生姜 MOL000675oleic acid(油酸)33.130.1445半夏 MOL000492(+)-catechin((+)-兒茶素)54.830.249大棗,白芍 MOL002670cavidine(卡文定堿)35.640.8124半夏 MOL000096(-)-catechin((-)-兒茶素)49.680.249大黃,大棗 MOL006594eciphin(卵磷脂)43.350.0323半夏 MOL002914eriodyctiol (flavanone)41.350.246黃芩,枳實 MOL006932l-Pseudoephedrine(左旋偽麻黃堿)45.010.0321半夏 (淫羊藿醇(黃烷酮)) MOL001924paeoniflorin(芍藥苷)53.870.794白芍 MOL000359sitosterol(谷甾醇)36.910.753黃芩,白芍 MOL001919(3S,5R,8R,9R,10S,14S)-3,17-43.560.532白芍 MOL000354isorhamnetin(異鼠李素)49.600.3130柴胡 dihydroxy-4,4,8,10,14-pentamethyl- MOL004609areapillin(茵陳黃酮)48.960.4114柴胡 2,3,5,6,7,9-hexahydro-1H-cyclopenta MOL0045983,5,6,7-tetramethoxy-2-(3,4,5-31.970.5910柴胡 [a]phenanthrene-15,16-dione trimethoxyphenyl) chromone MOL001918paeoniflorgenone(芍藥苷元)87.590.371白芍 (3,5,6,7-四甲氧基-2-(3,4,5- MOL000211mairin(丁子香萜)55.380.781白芍 三甲氧基苯基)色酮) MOL000627stepholidine(千金藤堿)33.110.5425大棗 MOL000490petunidin(矮牽牛花素)30.050.318柴胡 MOL007213nuciferin(核黃素)34.430.4024大棗 MOL000471aloe-emodin(蘆薈大黃素)83.380.2422大黃 MOL012921stepharine(千金藤林堿)31.550.3324大棗 MOL002235eupatin(澤蘭黃醇素)50.800.4114大黃 MOL002773beta-carotene(β-胡蘿卜素)37.180.5821大棗 MOL002281toralactone(決明子內酯)46.460.247大黃 MOL0061296-methylgingediacetate248.730.323生姜 MOL002268rhein(大黃酸)47.070.286大黃 (6-甲基銀杏二乙酸酯2) MOL000006luteolin(木犀草素)36.160.2554枳實 MOL001771poriferast-5-en-3beta-ol36.910.752生姜 MOL004328naringenin(柚皮素)59.290.2134枳實 (多孔甾醇-5-烯-3β-醇) MOL005828nobiletin(川陳皮素)61.670.5231枳實
2.2 肝細胞癌差異表達lncRNAs、miRNAs和mRNAs
通過GSE101728的基因芯片共獲得HCC差異基因447個(見圖1a),其中49個差異性lncRNAs(圖1b),篩選出832個lncRNA-miRNA調控關系和1 920個miRNA-mRNA調控關系。
2.3 活性成分-靶點和lncRNA-miRNA-mRNA轉錄網絡
將HCC差異表達基因與大柴胡湯潛在靶點取交集,獲得97個有效活性成分、19個關鍵lncRNAs、23個關鍵miRNAs和42個關鍵mRNAs。大柴胡湯治療HCC的活性成分-靶點網絡見圖2,其中活性成分槲皮素、木犀草素、漢黃芩素、川陳皮素和山柰酚的節點度較高(見表2)。lncRNA-miRNA-mRNA轉錄網絡見圖3,節點度較高的lncRNAs RUSC1-AS1、DIO3OS、LINC00487、FAM99A,miRNA miR-17-5p、miR-125b-5p、miR-24-3p和miR-107可能在網絡中起關鍵作用。核心靶點PPI網絡包括38個節點和284條邊,在潛在靶基因中,MYC、MMP2和CCND1節點度相對較高,可能是核心作用基因(見圖4)。
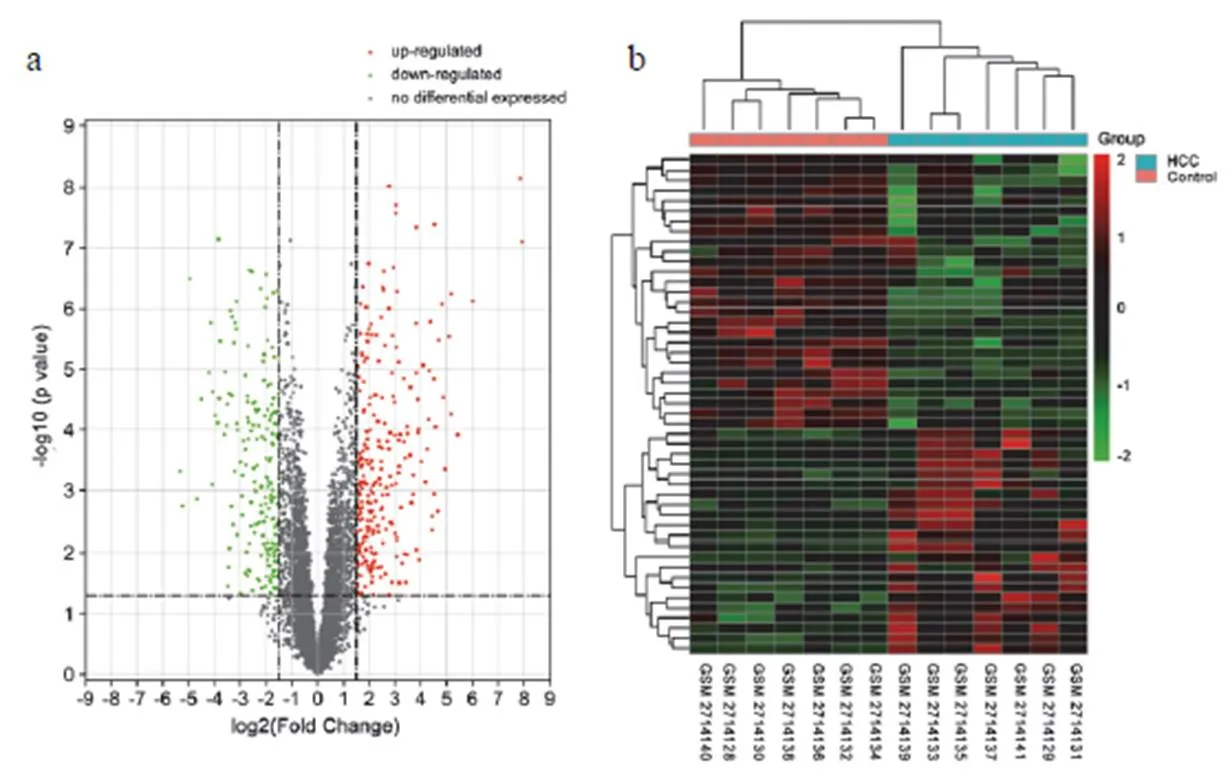
注:a.火山圖;b. lncRNAs熱圖
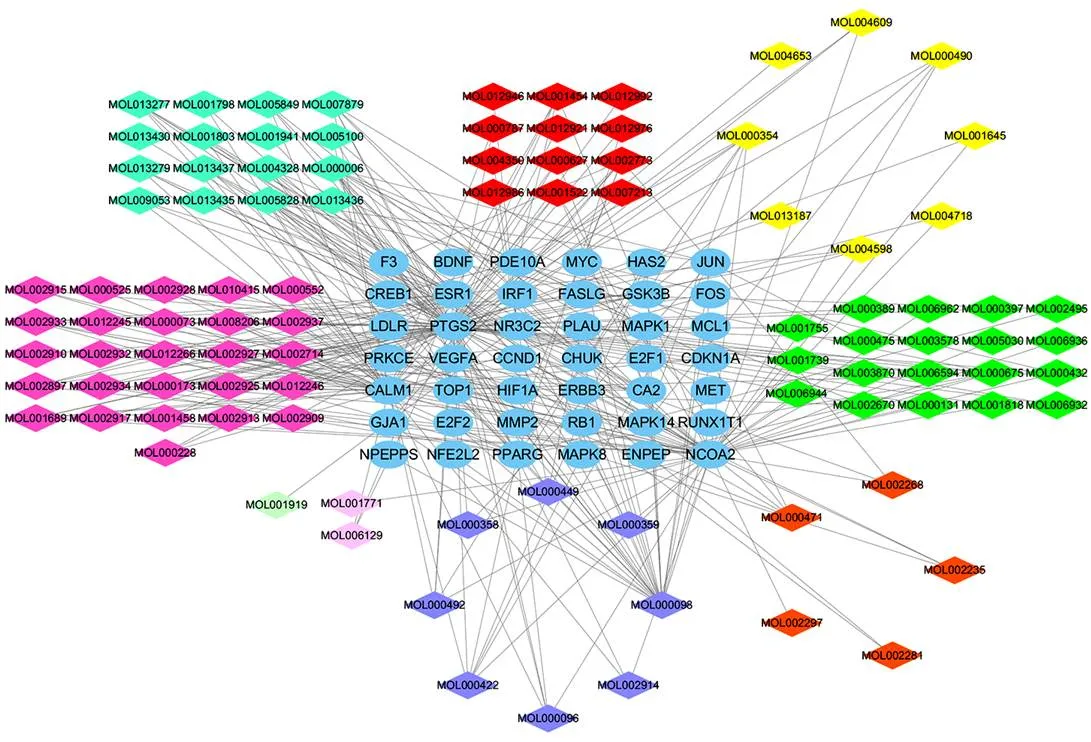
注:藍色為靶點基因;黃色為柴胡成分,橙色為大黃成分,青色為枳實成分,品紅為黃芩成分,綠色為半夏成分,淺綠為白芍成分,紅色為大棗成分,粉色為生姜成分,紫色為共有成分
表2 大柴胡湯治療HCC前10位候選活性成分及節點度
編號名稱節點度 MOL000098quercetin(槲皮素)26 MOL000006luteolin(木犀草素)13 MOL000173wogonin(漢黃芩素)10 MOL005828nobiletin(川陳皮素) 9 MOL000422kaempferol(山柰酚) 7 MOL000354isorhamnetin(異鼠李素) 7 MOL002773beta-carotene(β-胡蘿卜素) 7 MOL0029335,7,4’-Trihydroxy-8-methoxyflavone(5,7,4’-三羥基-8-甲氧基黃酮) 7 MOL000675oleic acid(油酸) 6 MOL002714baicalein(黃芩素) 6
2.4 關鍵靶點GO和KEGG富集分析
通過GO和KEGG富集分析揭示關鍵靶點參與的生物過程和途徑[15]。共獲得679個GO富集結果,其中分子功能主要為激活轉錄因子結合、RNA聚合酶Ⅱ轉錄因子結合、轉錄因子活性等,生物過程主要為對氧水平的響應、對金屬離子的響應、對體液水平的調節及成纖維細胞分化等,細胞組分主要為蛋白激酶復合物,絲氨酸、蘇氨酸蛋白激酶復合物,轉錄因子復合物等。共獲得107條KEGG通路富集結果,包括乙型肝炎、癌癥microRNA、蛋白聚糖、PI3K-Akt(磷酸肌醇3激酶-蛋白激酶B)信號通路、MAPK(促分裂原激活蛋白激酶)信號通路、TNF(腫瘤壞死因子)信號通路等。見圖5。
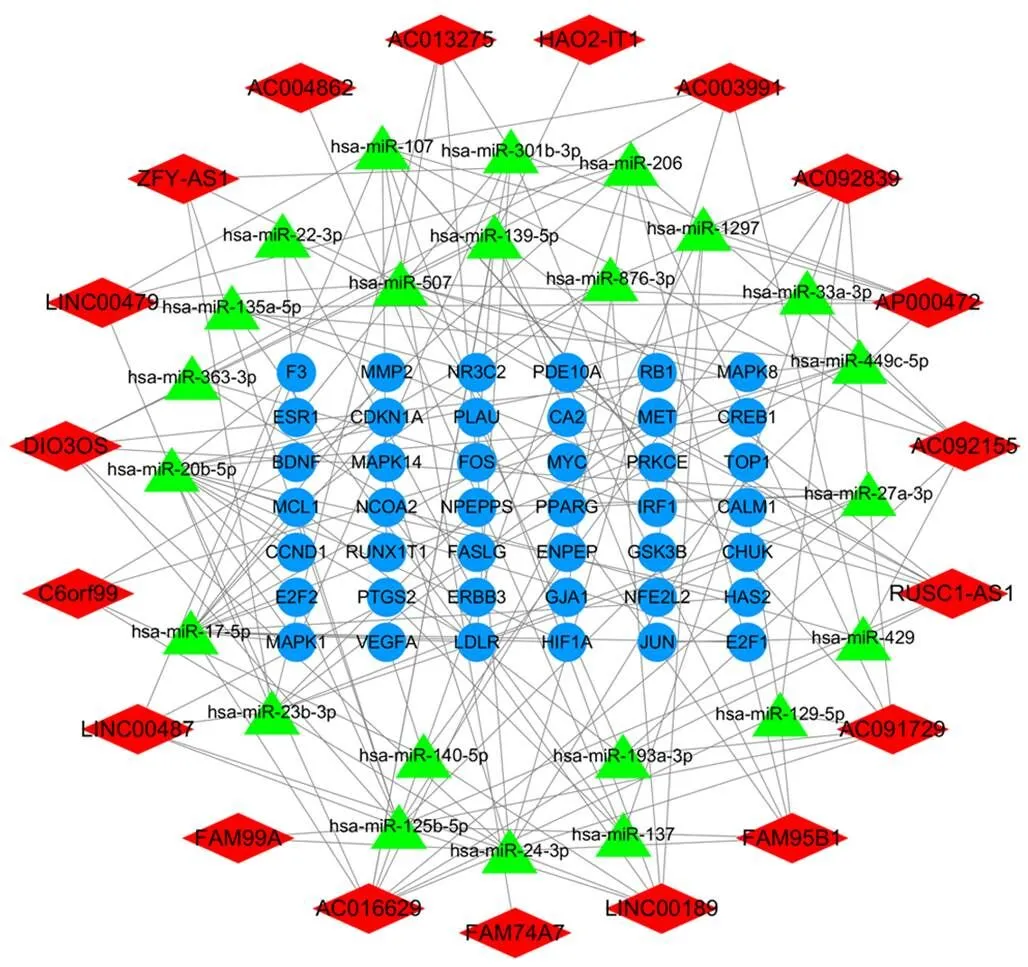
注:紅色為lncRNA,綠色為miRNA,藍色為mRNA
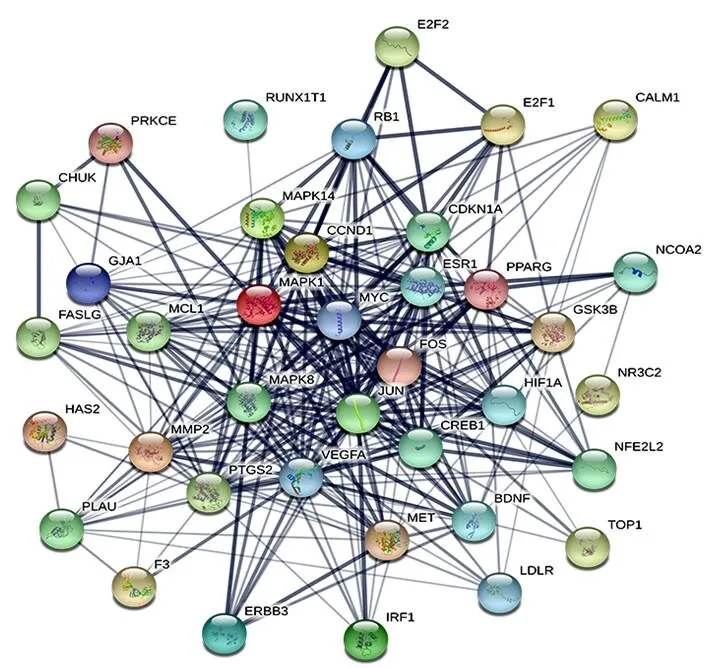
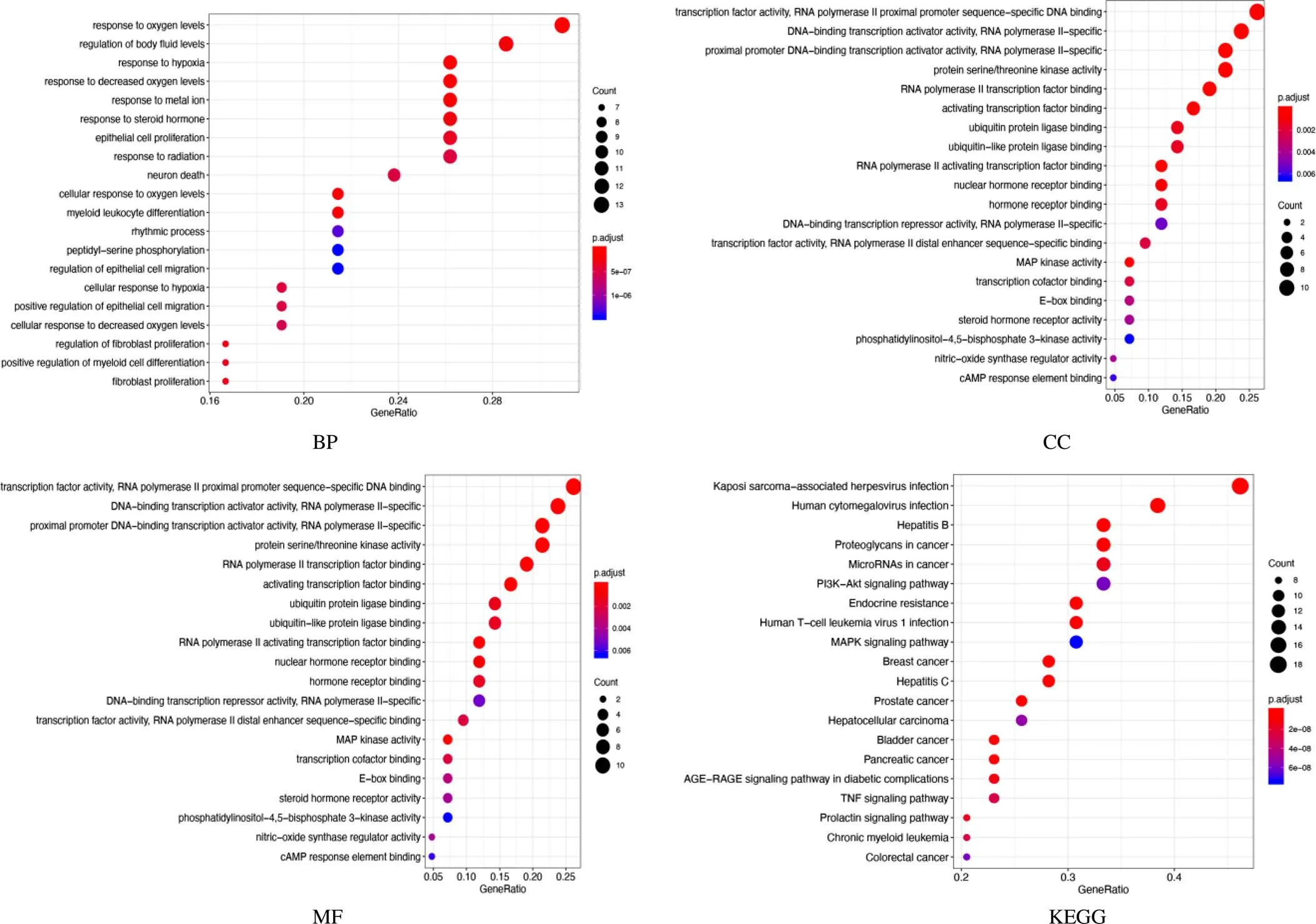
圖5 大柴胡湯治療HCC差異表達基因富集分析
2.5 分子對接分析
將大柴胡湯治療HCC的活性成分槲皮素、木犀草素、漢黃芩素、川陳皮素和山柰酚分別與PIK3CA(PDB ID:4tuu)、AKT1(PDB ID:6ccy)和MYC(PDB ID:6g6k)進行分子對接。PIK3CA和AKT1是KEGG主要通路PI3K-Akt信號通路中的標記蛋白,MYC是PPI網絡中節點度最高的靶點。當結合能<-5 kcal/mol時,配體與受體可以實現較好的結合[16]。分子對接結果表明,各活性成分均能與PIK3CA、AKT1和MYC很好結合(見表3),其中木犀草素與3個靶點的親和力較佳(見圖6)。
表3 大柴胡湯前5位候選活性成分與主要蛋白分子對接結果
活性成分分子式結合能/(kcal/mol) PIK3CAAKT1MYC 槲皮素C15H10O7-7.2-7.8-5.5 木犀草素C15H10O6-7.8-7.9-6.0 漢黃芩素C16H12O5-7.5-7.6-5.4 川陳皮素C21H22O8-7.5-7.1-4.9 山柰酚C15H10O6-8.3-7.8-5.5
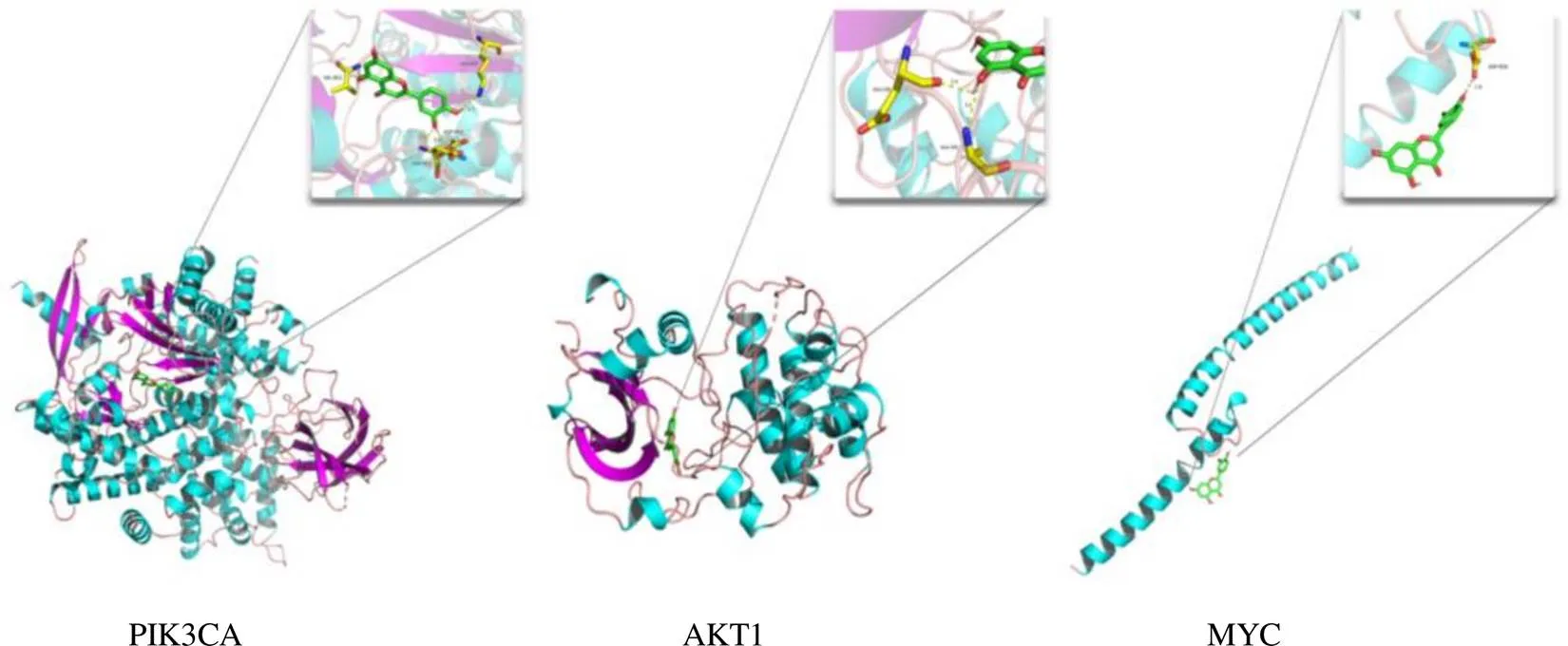
圖6 木犀草素與核心靶點分子對接模式圖
3 討論
由于HCC確診時通常已為晚期,且伴隨復雜的并發癥,針對性治療選擇有限,5年存活率僅為18%[17-18]。單一藥物成分和治療措施無法有效控制HCC的病理變化,中藥多成分、多靶點的作用特征為該病的治療提供了新思路[19]。中醫認為,腫瘤病機以熱毒瘀滯、氣機不暢為主,而肝為氣機調節的樞紐。因此,肝癌治療主要采用清熱、散結、疏通氣機之法,大柴胡湯為代表性處方[20-21]。既往的基礎研究及循證醫學證據表明,大柴胡湯對肝癌具有良好療效[6-7]。本研究從108種候選活性成分中篩選出大柴胡湯治療HCC的主要活性成分。其中,槲皮素(MOL000098,OB=46.43%,DL=0.28)是大柴胡湯最重要的化合物之一,其通過調節AKT-mTOR、PI3K、MAPK等途徑,以調節miRNA、增強金屬離子鋅活性、減少活性氧形成、下調絲氨酸-蘇氨酸激酶,在HCC治療中發揮抗氧化、抗增殖、促凋亡和抗糖酵解活性[22-26]。木犀草素(MOL000006,OB=36.16%,DL=0.25)通過調節PI3K、Akt信號通路在促進肝癌細胞凋亡中起重要作用,在HCC病理過程中可抑制炎癥和肝臟纖維化[27-28]。漢黃芩素(MOL000173,OB=30.68%,DL=0.23)通過下調p-Akt、周期蛋白D(cyclin D)和基質金屬蛋白酶2(MMP2)抑制HCC的增殖和侵襲[29]。川陳皮素(MOL005828,OB=61.67%,DL=0.52)在體內外對HCC均可發揮明顯的抑制作用[30]。山柰酚(MOL000422,OB=41.88%,DL=0.24)可有效緩解HCC中的肝纖維化,降低缺氧條件下缺氧誘導因子-1(HIF-1)和MAPK活性,并抑制肝癌細胞的存活;它與木犀草素結合,通過活性氧介導的線粒體,靶向獲得對HCC中腫瘤細胞的選擇性殺傷能力[31-33]。以上研究均支持本研究網絡預測結果,反映了大柴胡湯治療HCC的“多成分”特征。
非編碼RNA(ncRNA)通過調節蛋白質編碼基因的轉錄以加速癌癥的進展。作為典型的ncRNA,越來越多參與包括HCC在內的腫瘤發展過程的lncRNA和miRNA被評估鑒定出來[34]。本研究基于GEO數據庫HCC表達譜篩選的差異表達基因構建了大柴胡湯治療HCC的lncRNA-miRNA-mRNA轉錄網絡。該網絡中有42個潛在mRNAs,其中在PPI網絡中節點度較高的關鍵靶點均與HCC相關,節點度最高的MYC(包括c-MYC、n-MYC和l-MYC)為重要的癌基因。不受控制的信號傳導是肝癌的關鍵驅動力,MYC通過與ncRNA的復雜串擾參與HCC細胞周期過程、細胞凋亡、代謝、血管生成和轉移[35-36]。MMP2、E2F1和CCND1(cyclin D1)主要參與肝癌的細胞周期、增殖和凋亡,VEGFA參與血管生成過程[37-38]。分子對接結果表明,MYC與大柴胡湯治療HCC的主要活性成分結合良好。miRNA通過調節靶基因在HCC發生中起特定作用。在轉錄網絡miRNA中節點度最高的miR-17-5p(屬于miR-17-92簇)是HCC的關鍵標志物,可調控多個功能靶點。miR-17-5p通過c-MYC的調節回路,以及MMP2和MAPK的激活促進HCC的血管生成、增殖、侵襲和轉移。在非轉移性肝癌中,miR-17-5p與E2F1和c-MYC形成反饋回路,導致Caspase介導的細胞凋亡[39-41]。miR-24-3p和miR-125b-5p為乙型肝炎病毒(HBV)相關HCC的生物標志物,并且miR-107靶向MMP2以介導HCC侵襲[42-44]。lncRNA參與了HCC發生和發展的許多方面,其中RUSC1-AS1能通過miR-7-5p促進HCC細胞增殖并抑制細胞凋亡,DIO3OS通過競爭性結合miRNA調節HCC的惡性侵襲,LINC00487被確定為HCC患者纖維化的預測因子,FAM99A與HIF-1α相關聯,以在缺氧誘導下介導HCC的轉移[45-48]。因此,在此轉錄網絡中,大柴胡湯可能通過RUSC1-AS1等lncRNAs,參與調控miR-17-5p等miRNAs對mRNA的MYC、E2F1反饋回路,進而參與HCC血管生成、增殖、侵襲和轉移過程,發揮治療作用。大柴胡湯治療HCC的lncRNA-miRNA-mRNA轉錄網絡在一定程度上反映了大柴胡湯治療HCC的“多靶點”特征。
大柴胡湯治療HCC的靶基因為轉錄網絡中分子功能的實現者,進一步對其進行GO和KEGG富集分析,結果表明,肝癌中大柴胡湯調控的轉錄本與轉錄因子活性、對氧水平的響應、蛋白激酶復合物、成纖維細胞分化、PI3K-Akt信號通路、MAPK信號通路和TNF信號通路密切相關。該預測結果與以往研究一致,在細胞和分子水平上,HCC的發生和發展機制與氧化應激、通過miRNA的HBV和宿主的相互調節、纖維化微環境和蛋白激酶引起的信號轉導異常密切相關[43]。PI3K-Akt信號通路是調節細胞生長、代謝和存活的經典途徑,在HCC的1/3~1/2中被激活,并且與腫瘤級別、血管浸潤、肝內轉移、細胞凋亡和基質金屬蛋白酶的上調密切相關。MAPK途徑激活轉錄因子基因c-MYC、c-FOS和c-JUN,以驅動HCC的生長和分化。TNF通過細胞因子激活生長信號以影響HCC的侵襲,TNF-α信號通路、MAPK信號通路則促進HCC的增殖和轉移[49-51]。分子對接結果顯示,大柴胡湯治療HCC的主要活性成分可與PI3K-Akt信號通路中的相關蛋白很好地結合,從而進一步證明了預測的準確性。大柴胡湯對HCC的治療與多種信號通路和多種生物學過程有一定聯系,反映了大柴胡湯治療HCC的“多途徑”特征。
本研究通過整合分析探索了大柴胡湯治療HCC的機制,提示大柴胡湯的多種活性成分在HCC治療中具有協同作用,并且涉及對lncRNA-miRNA-mRNA網絡串擾的調節。轉錄調控網絡中的靶基因主要與腫瘤的存活、凋亡、自噬、氧化應激、蛋白激酶等密切相關,并可能通過PI3K-Akt信號通路、MAPK信號通路和TNF信號通路發揮作用,影響HCC的增殖和存活。本研究結果反映了大柴胡湯治療疾病多成分、多靶點、多途徑的特點,可為大柴胡湯治療HCC提供理論依據,并為進一步臨床和實驗研究提供方向。
[1] 閆京濤,范宏宇,李竹生.大柴胡湯治療肝熱血瘀型肝癌經驗體會[J].光明中醫,2020,35(11):1728-1730.
[2] 王鑫,杜娟,楊洋.加味大柴胡湯聯合化療治療晚期膽道腫瘤壓迫引起的梗阻性黃疸的療效觀察[J].現代中醫藥,2020,40(3):83-87.
[3] 陸國輝,李艷茹.大柴胡湯含藥血清通過Sirt3線粒體途徑誘導人肝癌HepG2細胞凋亡的研究[J].中藥藥理與臨床,2014,30(5):17-21.
[4] 許小凡,姜婷婷,劉芳,等.大柴胡湯調控TGF-β/Smad信號通路對DBTC聯合乙醇誘發小鼠胰腺纖維化的防治作用[J].中國應用生理學雜志, 2016,32(5):444-448,482.
[5] YANG J M, SUN Y, WANG M, et al. Regulatory effect of a Chinese herbal medicine formula on nonalcoholic fatty liver disease[J]. World Journal of Gastroenterology,2019,25(34):5105-5119.
[6] ROBINSON E K, COVARRUBIAS S, CARPENTER S. The how and why of lncRNA function:an innate immune perspective[J]. Biochim Biophys Acta Gene Regul Mech,2020,1863(4):194419.
[7] HUANG Z, ZHOU J K, PENG Y, et al. The role of long noncoding RNAs in hepatocellular carcinoma[J]. Mol Cancer,2020,19(1):77.
[8] KLIESER E, MAYR C, KIESSLICH T, et al. The crosstalk of miRNA and oxidative dtress in the liver:from physiology to pathology and clinical implications[J]. Int J Mol Sci,2019,20(21):5266.
[9] PENG Y, CROCE C M. The role of microRNAs in human cancer[J]. Signal Transduct Target Ther,2016,1:15004.
[10] 黃瑞紅,陽巧鳳,黃瑞玉,等.LncRNA LOC100505501在柴胡皂苷D逆轉MCF-7/ADR細胞耐藥中的作用[J].中南藥學,2018,16(12):1720-1724.
[11] 李燕,潘小平,王海霞,等.大黃素甲醚調節miR-370誘導肝癌細胞凋亡的實驗研究[J].中華介入放射學電子雜志,2018,6(2):148-153.
[12] 宋超,劉霞,邵啟祥,等.黃芩素對肝癌HepG2細胞增殖和凋亡的影響及miR-34a在其機制中的作用[J].江蘇大學學報(醫學版),2011,21(6):470-475.
[13] GUO W, HUANG J, WANG N, et al. Integrating network pharmacology and pharmacological evaluation for deciphering the action mechanism of herbal formula Zuojin pill in suppressing hepatocellular carcinoma[J]. Front Pharmacol,2019,10:1185.
[14] NEUTEBOOM R F, VERBRAAK E, WIERENGA-WOLF A F, et al. The monocyte transcriptome during pregnancy in multiple sclerosis:prominent expression of the Fc-receptor CD64[J]. Multiple Sclerosis Journal,2011,17(4):389-396.
[15] 陳博威,易健,劉柏炎,等.基于網絡藥理學與分子對接探討黃芪-女貞子藥對治療阿爾茨海默病的作用機制[J].天然產物研究與開發, 2020,32(12):2040-2050.
[16] 張良,楊雙蓉,李淑瑩,等.基于網絡藥理學和實驗驗證探討槲皮素治療心力衰竭的分子機制[J].中國實驗方劑學雜志,2021,27(15):156-165.
[17] FORNER A, REIG M, BRUIX J. Hepatocellular carcinoma[J]. Lancet, 2018,391(10127):1301-1314.
[18] VILLANUEVA A. Hepatocellular carcinoma[J]. N Engl J Med,2019, 380(15):1450-1462.
[19] CHEN F, ZHONG Z, TAN H Y, et al. Uncovering the anticancer mechanisms of Chinese herbal medicine formulas:therapeutic alternatives for liver cancer[J]. Front Pharmacol,2020,11:293.
[20] XI S Y, MINUK G Y. Role of traditional Chinese medicine in the management of patients with hepatocellular carcinoma[J]. World J Hepatol,2018,10(11):799-806.
[21] REN B, TAN L, XIONG Y, et al. Integrated analysis of the mechanisms of Da-Chai-Hu decoction in type 2 diabetes mellitus by a network pharmacology approach[J]. Evid Based Complement Alternat Med,2020,2020:9768414.
[22] DABBAGH-BAZARBACHI H, CLERGEAUD G, QUESADA I M, et al. Zinc ionophore activity of quercetin and epigallocatechin-gallate:from Hepa 1-6 cells to a liposome model[J]. J Agric Food Chem,2014, 62(32):8085-8093.
[23] JI Y, LI L, MA Y X, et al. Quercetin inhibits growth of hepatocellular carcinoma by apoptosis induction in part via autophagy stimulation in mice[J]. J Nutr Biochem,2019,69:108-119.
[24] SHAALAN Y M, HANDOUSSA H, YOUNESS R A, et al. Destabilizing the interplay between miR-1275 and IGF2BPs byand quercetin in hepatocellular carcinoma[J]. Nat Prod Res,2018, 32(18):2217-2220.
[25] WU H, PAN L, GAO C, et al. Quercetin inhibits the proliferation of glycolysis-addicted HCC cells by reducing hexokinase 2 and Akt-mTOR pathway[J]. Molecules,2019,24(10):1993.
[26] SALAMA Y A, EL-KAREF A, EL GAYYAR A M, et al. Beyond its antioxidant properties:Quercetin targets multiple signalling pathways in hepatocellular carcinoma in rats[J]. Life Sci,2019, 236:116933.
[27] IM E, YEO C, LEE E O. Luteolin induces caspase-dependent apoptosis via inhibiting the AKT, osteopontin pathway in human hepatocellular carcinoma SK-Hep-1 cells[J]. Life Sci,2018,209:259-266.
[28] SAGAWA H, NAIKI-ITO A, KATO H, et al. Connexin 32 and luteolin play protective roles in non-alcoholic steatohepatitis development and its related hepatocarcinogenesis in rats[J]. Carcinogenesis,2015,36(12):1539-1549.
[29] LIU X, TIAN S, LIU M, et al. Wogonin inhibits the proliferation and invasion, and induces the apoptosis of HepG2 and Bel7402 HCC cells through NF-κB, Bcl-2, EGFR and EGFR downstream ERK, AKT signaling[J]. Int J Mol Med,2016,38(4):1250-1256.
[30] MA X, JIN S, ZHANG Y, et al. Inhibitory effects of nobiletin on hepatocellular carcinoma in vitro and in vivo[J]. Phytother Res,2014,28(4):560-567.
[31] XU T, HUANG S, HUANG Q, et al. Kaempferol attenuates liver fibrosis by inhibiting activin receptor-like kinase 5[J]. J Cell Mol Med,2019,23(9):6403-6410.
[32] MYLONIS I, LAKKA A, TSAKALOF A, et al. The dietary flavonoid kaempferol effectively inhibits HIF-1 activity and hepatoma cancer cell viability under hypoxic conditions[J]. Biochem Biophys Res Commun,2010,398(1):74-78.
[33] SEYDI E, SALIMI A, RASEKH H R, et al. Selective cytotoxicity of luteolin and kaempferol on cancerous hepatocytes obtained from rat model of hepatocellular carcinoma:involvement of ROS-mediated mitochondrial targeting[J]. Nutr Cancer,2018,70(4):594-604.
[34] ANASTASIADOU E, JACOB L S, SLACK F J. Non-coding RNA networks in cancer[J]. Nat Rev Cancer,2018,18(1):5-18.
[35] DANG H, TAKAI A, FORGUES M, et al. Oncogenic activation of the RNA binding protein NELFE and MYC signaling in hepatocellular carcinoma[J]. Cancer Cell,2017,32(1):101-114.
[36] SWIER L J Y M, DZIKIEWICZ-KRAWCZYK A, WINKLE M, et al. Intricate crosstalk between MYC and non-coding RNAs regulates hallmarks of cancer[J]. Mol Oncol,2019,13(1):26-45.
[37] TAI L, HUANG C J, CHOO K B, et al. Oxidative stress down-regulates MiR-20b-5p, MiR-106a-5p and E2F1 expression to suppress the G1, S transition of the cell cycle in multipotent stromal cells[J]. Int J Med Sci,2020,17(4):457-470.
[38] YANG W, LI Z, QIN R, et al. YY1 Promotes endothelial cell-dependent tumor angiogenesis in hepatocellular carcinoma by transcriptionally activating VEGFA[J]. Front Oncol,2019,9:1187.
[39] YANG F, YIN Y, WANG F, et al. miR-17-5p promotes migration of human hepatocellular carcinoma cells through the p38 mitogen-activated protein kinase-heat shock protein 27 pathway[J]. Hepatology,2010,51(5):1614-1623.
[40] LIU D, DONG L, LIU Y, et al. A c-Myc, miR-17-5p feedback loop regulates metastasis and invasion of hepatocellular carcinoma[J]. Tumour Biol,2016,37(4):5039-5047.
[41] EL TAYEBI H M, OMAR K, HEGY S, et al. Repression of miR-17-5p with elevated expression of E2F-1 and c-MYC in non-metastatic hepatocellular carcinoma and enhancement of cell growth upon reversing this expression pattern[J]. Biochem Biophys Res Commun, 2013,434(3):421-427.
[42] MENG F L, WANG W, JIA W D. Diagnostic and prognostic significance of serum miR-24-3p in HBV-related hepatocellular carcinoma[J]. Med Oncol,2014,31(9):177.
[43] JIN Y, WONG Y S, GOH B K P, et al. Circulating microRNAs as potential diagnostic and prognostic biomarkers in hepatocellular carcinoma[J]. Sci Rep,2019,9(1):10464.
[44] XIAO D, GAO H X. Mechanism of miR-107-targeting of regulator of G-protein signaling 4 in hepatocellular carcinoma[J]. Oncol Lett,2019,18(5):5145-5154.
[45] CHEN Y A, CHENG L, ZHANG Y, et al. LncRNA RUSC1-AS1 promotes the proliferation of hepatocellular carcinoma cells through modulating NOTCH signaling[J]. Neoplasma,2020,67(6):1204-1213.
[46] SONG M, ZHONG A, YANG J, et al. Large-scale analyses identify a cluster of novel long noncoding RNAs as potential competitive endogenous RNAs in progression of hepatocellular carcinoma[J]. Aging (Albany NY),2019,11(22):10422-10453.
[47] YE J, WU S, PAN S, et al. Risk scoring based on expression of long non coding RNAs can effectively predict survival in hepatocellular carcinoma patients with or without fibrosis[J]. Oncol Rep,2020,43(5):1451-1466.
[48] ZHAO B, KE K, WANG Y, et al. HIF-1α and HDAC1 mediated regulation of FAM99A-miR92a signaling contributes to hypoxia induced HCC metastasis[J]. Signal Transduct Target Ther,2020,5(1):118.
[49] ALQAHTANI A, KHAN Z, ALLOGHBI A, et al. Hepatocellular carcinoma:molecular mechanisms and targeted therapies[J]. Medicina (Kaunas),2019,55(9):526.
[50] CHEN C, WANG G. Mechanisms of hepatocellular carcinoma and challenges and opportunities for molecular targeted therapy[J]. World J Hepatol,2015,7(15):1964-1970.
[51] ZHANG G P, YUE X, LI S Q. Cathepsin C interacts with TNF-α, p38 MAPK signaling pathway to promote proliferation and metastasis in hepatocellular carcinoma[J]. Cancer Res Treat, 2020,52(1):10-23.
Integration Analysis of lncRNA-miRNA-mRNA Transcriptional Network in the Treatment of Hepatocellular Carcinoma withDecoction
ZHENG Piao1, CHEN Bowei2, NING Dimin1, XIAO Wei3, TIAN Xuefei1
To characterize the lncRNA-miRNA-mRNA regulatory network ofDecoction in the treatment of hepatocellular carcinoma (HCC); To explore its anti-liver cancer mechanism.The active components ofDecoction and predicted targets, as well as the differentially expressed lncRNAs, miRNAs, and miRNAs of HCC were obtained by Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP), GEO, miRDB, miRTarBase and TargetScan databases. Furthermore, R language and Perl software were chosen to screen the key lncRNAs, miRNAs and mRNAs forDecoction for the treatment of HCC. With the help of Cytoscape 3.7.0 software and STRING database, the active components-target network, lncRNA-miRNA-mRNA regulatory network and protein-protein interaction network were built. GO and KEGG enrichment analysis were performed, and combined with molecular docking to construct integrated analysis.A total of 97 active components and 42 targets were obtained inDecoction for the treatment of HCC, and a lncRNA-mRNA-miRNA regulatory network composed of 19 key lncRNAs, 23 key miRNAs, and 42 key mRNAs was built. Enrichment analysis showed that these important target genes were involved in tumor survival, apoptosis, autophagy, oxidative stress, protein kinases and other biological processes, and mainly played a therapeutic role through PI3K-Akt signaling pathway, MAPK signaling pathway and TNF signaling pathway.This study indicates the characteristics ofDecoction treatment of HCC with multi-components, multi-targets, and multi-pathways synergistic therapy, and can provide basis and direction for future research.
hepatocellular carcinoma;Decoction; transcriptional network; bioinformatics; network pharmacology; molecular docking
R273.57;R285
A
1005-5304(2021)12-0022-08
10.19879/j.cnki.1005-5304.202104055

湖南省自然科學基金(2020JJ4066);湖南省研究生科研創新項目(CX20190541);湖南中醫藥大學中西醫結合一流學科開放基金(2018ZXYJH03)
田雪飛,E-mail:003640@hnucm.edu.cn
(2021-04-04)
(修回日期:2021-06-08;編輯:陳靜)
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中國生殖健康(2019年3期)2019-02-01 06:12:26
中成藥(2016年8期)2016-05-17 06:08:14
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
