華木蓮開花結實特性及其瀕危的影響因素1)
2021-12-03 07:13:36龍桂根陳發菊陳應德姚蕾鄭慶衍李乾明王勇
東北林業大學學報 2021年10期
龍桂根 陳發菊 陳應德 姚蕾 鄭慶衍 李乾明 王勇
(宜春市科學院,宜春,336000) (三峽大學) (宜春市科學院)
華木蓮(SinomanglietiaglaucaZ. X. Yu et Q. Y. Zheng)又名落葉木蓮,是木蘭科木蓮屬中唯一落葉樹種。華木蓮是1988年在江西省宜春市明月山國家森林公園境內發現的木蘭科新種,為我國特有種,國家一級瀕危保護植物,呈點狀狹窄地散生于袁州區洪江鄉木坪及玉京山一帶,野生種群數量十分稀少,為極度瀕危植物。該樹種發現以來,龍桂根等[1]對華木蓮生殖發育、居群遺傳結、種苗繁育、種質資源保護與篩選等方面進行了研究,并為保護、開發這一珍貴植物資源奠定了基礎。
華木蓮自然分布狹窄、個體數目少、遺傳基礎薄弱、自然更新困難等問題已經嚴重影響其天然種群的生存狀況[2-4]。華木蓮天然種群的穩定存在是研究、開發、利用這一特色種質資源的基礎,更是種質資源保護、優良種質挖掘、可持續生態林業發展的根本所在。繁育系統、生殖衰退往往是植物瀕危的關鍵因子[2-3]。為了更好地保護華木蓮野生種群,系統研究華木蓮繁育系統,分析其生殖生物學等內在因素對華木蓮瀕危的影響程度,是探尋并制定華木蓮野生種群保護策略的首要問題[2,5-6]。
本研究以華木蓮為研究對象,對其開花生物學特性、人工授粉及自然結實情況進行了初步研究,并以此為基礎分析其致瀕原因,提出保護措施,旨在為野生華木蓮的保護、遺傳育種及繁育提供參考,豐富瀕危植物保護生物學理論,為研究其他瀕危植物瀕危機制提供借鑒。
1 材料與方法
1.1 試驗材料
根據華木蓮自然分布情況、人工林生長情況、結實情況,選取個體數量在20株以上,長勢良好且結實相對穩定的3個不同地點華木蓮種群進行調查,其地理信息見表1。于2016、2017、2018、2019、2020年對3個不同地點的華木蓮開花結實特性進行觀測。所觀測的華木蓮樹齡均在10 a以上,生長健壯,連續正常開花結果4 a以上。

表1 3個華木蓮種群信息
1.2 華木蓮開花物候觀測
選取溫湯試驗林場種群中的5株華木蓮各15朵華木蓮花進行連續觀測,記錄花朵形狀、顏色、大小、雌蕊、雄蕊、花被片長度及片數。
華木蓮開花物候調查,主要是記錄始花時間、花朵開放、花瓣伸展、雄蕊個數、終花時間、單花持續時間、整株開花狀況、群體開花狀況等。
1.3 雜交指數的估算
雜交指數的估算按照Dafni的標準,根據花序直徑、花朵大小、開花過程中的測量數據,綜合評估華木蓮繁育系統[2,7]。隨機選擇10朵華木蓮花,測量其花朵直徑(d),以其平均值作為指標,d≤1 mm、1 mm
以上三者之和為雜交指數(OCI),其值為0時表明繁育系統是閉花授粉,為1時是專性自交,為2時是兼性自交,為3時是自交親和(有時需要傳粉者),為4時是部分自交親和,異交需要傳粉者[2,7]。
1.4 控制授粉試驗
4月初,隨機選擇3株樹各15個嫩綠色花苞進行4個不同處理的授粉試驗:花苞套袋處理,未作任何處理,僅套袋隔離;對照,不作任何處理,置于自然條件,同步觀測;人工控制授粉,在花苞成黃綠色時人為打開花瓣,并去雄,用同株或異株花朵的花粉進行授粉,授粉后,套袋隔離。套袋隔離3~5 d后,解開授粉袋,觀測其發育結實情況,直至9月中下旬結束觀測,以檢測其授粉特性。
1.5 果實特性測定
采種期為9月下旬,以樹上果實全變為麻黃色,果實微裂,見紅色假種皮為準。
結實特性調查:在采種時,對每個種群的開花結實情況(花期、果期)進行調查。
結實單株的頻率的統計:從3個種群中分別隨機選定3個20 m×20 m的固定樣地,從該種群出現開花結實的單株開始,每年統計結實單株的數量,連續統計5 a。5 a內開花結實單株數的最大值,即為該種群的結實單株數量,結實單株率=結實單株數量/全部活立木數量×100%。
萌芽試驗:隨機挑選15個單株家系的種子,分別取出帶有假種皮的種子及無假種皮的種子各100粒,分別播種,重復3次,待種子萌芽后,每天下午觀測種子萌芽情況,連續10 d萌芽數量無變化,則為最終萌芽數量,萌芽率=最終萌芽數量/種子粒數×100%。
假種皮質量測定:隨機挑選15個單株家系,分別采種后,分單株室內陰干至果殼全部裂開,取出種子并稱其質量(G1),搓洗假種皮,再次稱質量(G2),則單株種子質量為G1,假種皮質量分數=100%×(G1-G2)/G1。
1.6 數據處理
采用Excel2007對種群果實各性狀進行常規統計、分析,采用SPSS19.0對種群間果實性狀進行多重比較。
2 結果與分析
2.1 開花特性
2.1.1 花部特征
華木蓮的花單生枝頂,芳香,花被12片或16片,淡黃色,螺旋狀排列成3~4輪,每輪4片。外輪花被片為長橢圓形,長7.0~7.4 cm,寬1.8~2.0 cm;中輪花被片為披針形,長6.6~5.2 cm,寬1.3~1.7 cm;內輪花被片為披針狀線形,長5.5~6.0 cm,寬0.3~0.8 cm。雄蕊36~60枚,長0.6~0.7 cm,花絲極短,長約0.1 cm。心皮15~22枚,每枚心皮有6~8個胚珠。華木蓮雌雄同株同花,雄蕊聚生于圓錐狀花托基部周圍,長0.8~1.2 cm;雌蕊生于花托頂端(圖1)。

A.完全開放的花朵及完全閉合的花苞;B.除去部分花苞片的花苞;C.雌蕊和雄蕊;D.花苞片及雄蕊。
2.1.2 華木蓮開花過程
根據對華木蓮開花過程的觀測,華木蓮開花可以分為蕾期、初開期、盛開期、重合期、萎蔫期、凋零期。華木蓮蕾期從3月下旬開始,顯著特征為花芽開始顯著膨大,花苞苞片由嫩綠色轉為黃綠色,此時雌蕊已成熟,其柱頭呈白色并分泌粘液,可接受花粉,此階段持續約32 h,如圖2A;初開期,外輪花被片展開,內側2輪花被片仍緊密合攏,此時花粉成熟,授粉可充分進行,此階段持續約12 h,如圖2B;盛開期,所有花被片均展開,內輪花被片向外稍微傾斜,所有雄蕊呈散開狀,花粉全部成熟,柱頭處于不可授粉狀態,此階段持續約8 h,如圖2C;重合期,內側2輪花被片重新閉合,外輪花被片保持展開狀態,此階段持續時間約為8 h,與初開期間隔時間較短,為1.17~1.50 d左右,球果雛形形成,如圖2D;萎蔫期,從花被片頂端開始枯黃,到全部枯黃為止,持續時間約為20 h,如圖2E;凋零期,花被片、雄蕊均枯萎脫落,花期結束,如圖2F。由此可知,華木蓮單花花蕾期相對較長,約1.5 d,初開期至萎蔫期,花期為2~3 d。單株開花時期不一,陸續開放。

A.蕾期;B.初開期;C.盛開期;D.重合期;E.萎蔫期:F.凋零期。
2.1.3 雜交指數估算
根據對3個種群的華木蓮花朵觀測可知,華木蓮花朵直徑最小為7.5 cm,最大為16.8 cm,平均值為12.9 cm,為大型花,對應值為3。華木蓮花在花被片即將展開之時,其花藥未開裂,花粉未成熟,而雌蕊柱頭已經分泌出粘液,處于成熟狀態,雌蕊先熟,對應值為0。華木蓮花為雌雄同花,花被片散開時,其柱頭位置高于花藥,對應值為1。由此可見華木蓮雜交指數為4,為異花授粉為主,自花授粉為輔。
2.1.4 華木蓮授粉情況
對不同授粉處理的花朵結實情況統計結果見表2。結果表明,自然狀態的木蓮結實率僅為40.00%;套袋隔離狀態的結實率為13.33%,不能正常結實者在12 d左右花托自然脫落;同株異花授粉的結實率為73.33%;異株異花授粉的結實率為86.67%;人工去雄未授粉的花朵,花托全部脫落。結果表明,華木蓮以異花授粉為主,自花授粉為輔。
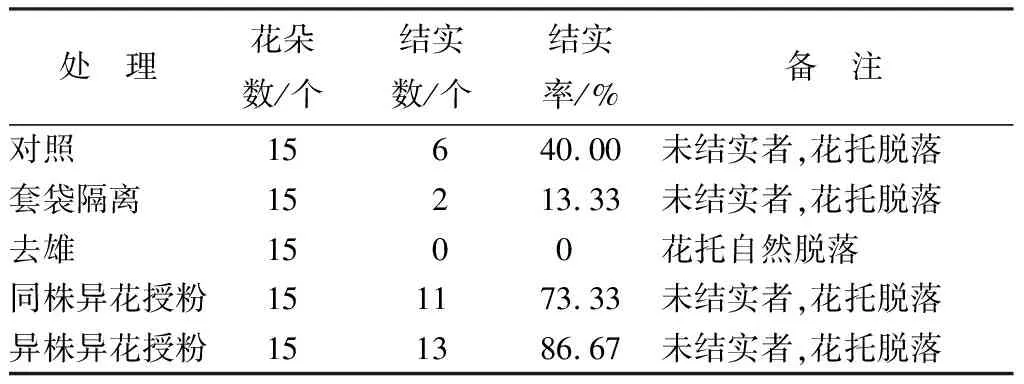
表2 華木蓮不同授粉處理的結實情況
根據華木蓮花朵特性、開花過程、雜交指數、套袋隔離花朵的結實特性,可以看出,由于華木蓮著花方式為頂生,花期短,開花后存在重新閉合現象,且其授粉方式為異花授粉,加之雌蕊先熟,從而導致華木蓮自然開花結實率低,甚至只開花不結實,這是其瀕危原因之一。
2.2 華木蓮的結實特性分析
2.2.1 華木蓮結實特性
對來自不同種群、不同樹齡的華木蓮活立木進行連續10 a的觀測,發現袁州區溫湯鎮、袁州區洪江鄉、奉新縣采育林場的華木蓮初始開花結實的樹齡分別為6、7、6 a,其平均樹齡為6.33 a;盛果期分別為11、13、10 a,盛果期出現的平均樹齡為11.33 a。所有結實量較多的單株一般處于林窗處或林緣,林分內的單株果實多集中在樹冠上部,這一現象表明華木蓮結實過程需要較多的光照。
近3 a觀測,發現wt-19(wt為溫湯林場華木蓮種群,19為單株編號)、hj-3(hj為洪江木坪村華木蓮種群,3為單株編號)、hj-35、hj-21、wt-7等10個單株在2017、2018年均有結實,2019年wt-19、wt-10、wt-11、wt-1等活立木未結實。同時可以看出,3 a內,華木蓮單株果實產量出現明顯的“大小年”現象,整體平均值2017年、2018年、2019年分別為11.2、19.0、5.9 kg·株-1。對于單株華木蓮活立木,每年果實產量存在較大變化,極度不穩定,如wt-4單株在2018年結實量達到50.1 kg·株-1,而2019年僅為3.0 kg·株-1。
3 a連續的觀測結果也顯示,華木蓮人工林結實量較相同樹齡的天然林結實量相比分別高3.53%、4.12%、3.75%,但差異不顯著。同時,華木蓮人工林、天然林種群中,結實單株分別占到整體林分的42.18%、37.95%,即未結實的單株占一半以上。

表3 部分華木蓮不同單株不同年份結實量
2.2.2 華木蓮種子假種皮質量分數
為更好地分析華木蓮果實特性,對華木蓮種子的單株產量、假種皮質量進行了測定分析,結果見表4。華木蓮種子假種皮質量平均占整個種子質量的58.37%,最大占66.16%,最小占41.30%。發芽試驗中,帶假種皮的種子發芽率僅有10.20%,其最低發芽率為3.50%;搓洗掉假種皮的種子發芽率達到95.70%,最低為81.00%。實際生產中,假種皮未搓洗干凈的種子發芽率降低,甚至不萌芽。趙武[8]、俞志雄等[9]的研究認為,肉質假種皮中大多數為萜烯類、醇類化合物,其可以保護種子,防止失水,但對種子萌芽具有明顯的抑制作用。自然脫落的華木蓮種子,少部分肉質假種皮被鼠類、鳥類消化,絕大部分種子的假種皮在地上容易迅速霉變腐爛,導致種子霉變而腐爛,這也是華木蓮自然條件難以進行林下更新的直接原因。

表4 華木蓮肉質假種皮的質量分數及其對種子發芽率的影響

續(表4)
3 結果與討論
華木蓮單花壽命2~3 d,盛開期僅8 h,整個種群持續開花,存在開花后重新閉合現象,難以吸引傳粉者,這是導致華木蓮瀕危的重要原因之一。本研究認為,華木蓮單花開花過程可分為6個階段,花期為2~3 d,開花后存在重新閉合直至凋零的現象。整個種群花期持續15 d左右,持續開花,無盛花高峰期。開花期為4月中下旬,多為陰雨天氣。華木蓮花朵對傳粉昆蟲基本沒有吸引力,導致授粉不充分,自然結實率低。華木蓮雜交指數為4,為異花授粉為主,自花授粉為輔。從雜交指數及套袋授粉可以看出,華木蓮繁育系統為異交,初開期雌蕊為成熟末期,而此時花藥未開裂,花粉剛開始成熟,兩者同時具有活力的時間僅為5 h左右。華木蓮雌蕊在上,雄蕊在下,雌雄異位,自花授粉難以進行,異花授粉又無活躍的授粉昆蟲[10-12],這也是華木蓮瀕危的重要因素之一。
華木蓮種群結實率低(42.18%~37.95%)、不穩定,原因是華木蓮結實需要較多的光照,且其花期短,開花后存在重新閉合的現象。華木蓮授粉方式為異花授粉,加之雌蕊先熟,導致華木蓮自然開花結實低,甚至只開花不結實,從而導致其瀕危。
華木蓮種子假種皮過厚,抑制萌芽。華木蓮種子肉質假種皮質量達到整個種子的57.37%,帶有假種皮的種子萌芽率僅有10.20%,這進一步說明較厚的假種皮會降低華木蓮種子的萌發率。
本研究認為,華木蓮花朵結構、開花過程、開花模式、授粉特性、種子生物學特征均為其瀕危的主要因素。華木蓮花期持續開花模式、同期開花數量少、單花花期極短、開花時期陰雨低溫,這些都影響華木蓮花朵對授粉昆蟲的吸引力。華木蓮雄蕊聚生于圓錐狀花托基部,雌蕊螺旋聚生其上部,柱頭可授性遠早于花粉成熟,進一步加劇華木蓮授粉的難度。授粉試驗表明,華木蓮自然授粉結實率低,僅為40%,而同株異花、異株異花授粉結實率達到73%、87%,這進一步說明華木蓮異花授粉結實率高。另外,華木蓮種子假種皮過厚,抑制種子萌芽并導致其霉變腐爛,直接降低了華木蓮天然更新的可能性。華木蓮幼苗前期喜陰后期喜陽的特性,也是華木蓮瀕危的原因。因此,在華木蓮開花期間,開展人工輔助授粉,并及時收集華木蓮種子,去除其肉質假種皮,人工促進其萌芽,是提高華木蓮結實率、種子萌芽率的可行性手段,也是提高其種群多樣性途徑。
本研究通過對華木蓮不同種群開花結實過程連續跟蹤觀測、分析,從其開花結實物候、繁育系統、授粉特征、結實特性等方面對華木蓮進行了初步研究,分析了其繁育生殖致瀕機制,為華木蓮進一步開發提供參考。
