片段化森林中檵木幼苗種內(nèi)葉經(jīng)濟(jì)性狀變異及其驅(qū)動(dòng)因子
2021-12-02 01:09:54陳春何建平鄭詩(shī)璐吳雁南王瑩徐高福于明堅(jiān)
浙江大學(xué)學(xué)報(bào)(理學(xué)版) 2021年6期
關(guān)鍵詞:植物經(jīng)濟(jì)
陳春,何建平,鄭詩(shī)璐,吳雁南,王瑩,4,徐高福,于明堅(jiān)*
(1.浙江大學(xué)生命科學(xué)學(xué)院,浙江杭州 310058;2.淳安縣新安江生態(tài)開(kāi)發(fā)集團(tuán)有限公司,浙江 淳安 311700;3.西澳大學(xué) 生物科學(xué)學(xué)院,珀斯 6009;4.浙江大學(xué) 農(nóng)業(yè)與生物技術(shù)學(xué)院,浙江 杭州 310058)
面對(duì)外界多變的環(huán)境,植物在生長(zhǎng)與生存間進(jìn)行權(quán)衡[1],為使資源獲取和利用最大化,與植物資源利用策略相關(guān)的葉功能性狀(又稱葉經(jīng)濟(jì)性狀,leaf economic trait)相互關(guān)聯(lián)、協(xié)同變化,形成葉經(jīng)濟(jì)譜(leaf economics spectrum)[2-4]。葉經(jīng)濟(jì)譜對(duì)了解植物與環(huán)境之間的關(guān)系、群落構(gòu)建和生態(tài)系統(tǒng)過(guò)程等具有重要作用[5-7]。以往對(duì)葉經(jīng)濟(jì)譜的研究大多集中于物種水平[1,8-9],但種內(nèi)性狀變異占總變異的25%[10],其對(duì)種群延續(xù)乃至群落構(gòu)建的意義不容忽視[11-13]。近期有學(xué)者對(duì)種間葉經(jīng)濟(jì)譜之間的關(guān)系是否也存在于種內(nèi)進(jìn)行了探討[14-18]。如LAFOREST?LAPOINTE 等[15]以歐洲赤松(Pinus sylvestris)為研究對(duì)象,發(fā)現(xiàn)比葉面積與葉氮含量無(wú)關(guān),這與WRIGHT 等[2]發(fā)現(xiàn)的基于物種水平的全球葉經(jīng)濟(jì)譜間的關(guān)系不一致。而MARTIN 等[19]通過(guò)研究咖啡(Coffea arabica)的性狀指出,咖啡也存在一定的種內(nèi)葉經(jīng)濟(jì)譜,但明顯弱于物種水平的葉經(jīng)濟(jì)譜。生境片段化是影響生物多樣性的重要因素之一,往往會(huì)導(dǎo)致種群?jiǎn)适Ш瓦m合度下降[20]。全球森林片段化現(xiàn)象十分嚴(yán)重,約有70%的森林群落處于邊緣1 km 范圍內(nèi);我國(guó)大多數(shù)森林斑塊面積較小,小于10 hm2[21-22]。目前針對(duì)片段化生境中的種內(nèi)葉經(jīng)濟(jì)譜的研究較少。
種內(nèi)葉經(jīng)濟(jì)譜的形成,主要由環(huán)境因子驅(qū)動(dòng)[23]。其中,光照、土壤養(yǎng)分和水分等生境因子的異質(zhì)性是種內(nèi)葉經(jīng)濟(jì)性狀變異的重要來(lái)源[17,24-26]。例如,比葉面積隨光照強(qiáng)度的增強(qiáng)逐漸降低[27]、隨土壤氮元素含量的增加而增加[28]。在生境片段化過(guò)程中,面積效應(yīng)和邊緣效應(yīng)等促使生境條件如光照、土壤性質(zhì)產(chǎn)生強(qiáng)烈的變化,從而對(duì)植物產(chǎn)生影響[29]。種內(nèi)葉經(jīng)濟(jì)性狀的變異與協(xié)調(diào)可通過(guò)作用于個(gè)體的生長(zhǎng)和存活,進(jìn)而影響植物在片段化生境中的適合度[30],而幼苗期是植物更新的關(guān)鍵階段,也是植物生活史中對(duì)生境變化最為敏感的階段,在很大程度上決定了植物在群落內(nèi)的適合度[31]。針對(duì)片段化生境中幼苗種內(nèi)葉經(jīng)濟(jì)譜及其機(jī)理的研究,對(duì)于理解片段化生境中植物的適應(yīng)和更新策略乃至群落物種多樣性維持機(jī)制均十分重要。
檵木(Loropetalum chinense)為金縷梅科檵木屬植物,廣泛分布于北回歸線以北地區(qū),常見(jiàn)于我國(guó)中部、南部及西南各省[32]。檵木具有很強(qiáng)的適應(yīng)性,可成為早期群落演替的先鋒樹(shù)種,對(duì)生態(tài)系統(tǒng)功能維持具有重要作用[33],同時(shí)還具有一定的藥用價(jià)值[34]。浙江千島湖庫(kù)區(qū)是1959 年因水庫(kù)大壩建設(shè)形成的片段化森林,檵木作為千島湖庫(kù)區(qū)島嶼和周邊大陸馬尾松林內(nèi)的常見(jiàn)種,在千島湖長(zhǎng)期監(jiān)測(cè)樣地中其相對(duì)多度占40.48%[35]。本研究以千島湖庫(kù)區(qū)馬尾松林內(nèi)的檵木幼苗為對(duì)象,測(cè)定了7 種與資源獲取、生長(zhǎng)和存活緊密相關(guān)的葉經(jīng)濟(jì)性狀,以及幼苗所處的生境條件,探討其是否存在種內(nèi)葉經(jīng)濟(jì)譜,并比較了大陸、大島內(nèi)部、大島邊緣、小島4 種不同生境下檵木幼苗葉經(jīng)濟(jì)性狀的差異,以及性狀與生境條件之間的關(guān)系,旨在揭示片段化生境中檵木幼苗種內(nèi)葉經(jīng)濟(jì)性狀的變化機(jī)制,為進(jìn)一步了解片段化生境中常見(jiàn)植物的更新策略提供基礎(chǔ)。
1 材料和方法
1.1 研究地區(qū)概況
研究地千島湖(新安江水庫(kù))位于我國(guó)東部的浙江省杭州市淳安縣內(nèi)(29°22′N(xiāo)~29°50′N(xiāo),118°34′E~119°15′E;圖1),由1959 年新安江水電站修建形成。千島湖水域面積約540 km2,水位在108 m 時(shí)共有1 078 個(gè)面積大于0.25 hm2的陸橋島嶼。該地區(qū)屬于典型的亞熱帶季風(fēng)氣候,溫暖濕潤(rùn)、四季分明,雨熱同期。年平均氣溫為17 ℃,年平均降水量為1 430 mm。土壤類(lèi)型主要為紅壤、黃壤和巖性土。在水庫(kù)建成前,原有森林遭到砍伐,各島嶼在同一時(shí)間開(kāi)始次生演替,至今已有60 多年,現(xiàn)有植被組成相似,森林覆蓋率達(dá)88.5%,主要為次生馬尾松林,林下多為檵木、短尾越橘(Vaccinium carlesii)、杜鵑(Rhododendron simsii)和格藥柃(Eurya muricata)等物種[35]。

圖1 島嶼與大陸樣點(diǎn)分布Fig.1 Study islands and mainland in the Thousand Island Lake region
1.2 數(shù)據(jù)采集
1.2.1 植物樣品采集與性狀測(cè)定
選取5 個(gè)小島(0.36~0.84 hm2),5 個(gè)大島(3.70~1 153.88 hm2)和5 個(gè)大陸點(diǎn)(圖1;附表1),小島和大陸每個(gè)點(diǎn)選10 株幼苗,大島邊緣(距邊緣<20 m)和大島內(nèi)部(距邊緣>40 m)各選10 株,幼苗高度均<1.3 m。2019 年7 月,對(duì)每株幼苗采集5~10 片健康向陽(yáng)葉片,裝于密封袋中,冷藏保存,并在采集后12 h 內(nèi)進(jìn)行性狀測(cè)定。在葉性狀中,葉面積(leaf area,LA)與水分平衡、光合能力有關(guān);葉干物質(zhì)質(zhì)量分?jǐn)?shù)(leaf dry matter content,LDMC)與葉碳氮比(leaf carbon to nitrogen ratio,C∶N)可反映植物對(duì)外界的防御能力[36-37];而比葉面積(specific leaf area,SLA)體現(xiàn)植物對(duì)光的捕獲效率,與葉綠素濃度(chlorophyll concentration,Chl)在一定程度上反映植物的光合能力[38-39];碳是重要的結(jié)構(gòu)物質(zhì),也參與新陳代謝[40];氮與植物的最大光合效率緊密相關(guān)[36]。本研究將測(cè)定上述性狀。將采集到的葉片擦拭干凈后去除葉柄,稱鮮重,置入掃描儀中用萬(wàn)深葉片分析系統(tǒng)(2018 版;www.wseen.com)掃描并計(jì)算葉片總面積,即為L(zhǎng)A 乘以葉片數(shù)。用葉綠素儀(SPAD-502 Plus,Konica Minolta,Japan)避開(kāi)葉脈測(cè)定每張葉片的葉綠素,測(cè)3 個(gè)點(diǎn)求均值,用其代表該葉片單位面積的葉綠素總濃度。葉片于60 ?C 烘箱中烘72 h,取出稱干重。LDMC 為干重與鮮重之比,SLA 為面積與干重之比,Chl 為葉綠素濃度乘以SLA。用測(cè)得的所有葉片性狀值的均值表示該個(gè)體相應(yīng)的性狀值。各項(xiàng)性狀測(cè)定完成后,將葉片干燥粉碎過(guò)80 目篩,稱取50 mg 待測(cè)樣品,用錫箔紙包裹,用元素分析儀(vario MACRO cube,Elementar,Germany)CN 模式測(cè)定葉全碳量(leaf total carbon content,C)、葉全氮量(leaf total nitrogen content,N)。

附表1 島嶼和大陸樣點(diǎn)基本信息Table S1 The general information of study islands and mainland sites
1.2.2 生境因子測(cè)定
2019 年6 月,用裝有魚(yú)眼鏡頭(Sigma EX DC 4.5 mm)的相機(jī)于每個(gè)個(gè)體所在位置1.3 m 處朝上拍攝,其后用HemiView 軟件(Hemiview 2.1 SR4)分析照片,得到群落郁閉度。2019 年10 月,在每個(gè)個(gè)體下方用環(huán)刀取土帶回實(shí)驗(yàn)室測(cè)定土壤最大持水量(maximum soil water retaining capacity,MWRC),測(cè)定方法見(jiàn)《森林土壤水分-物理性質(zhì)的測(cè)定》(中華人民共和國(guó)林業(yè)行業(yè)標(biāo)準(zhǔn)LY/T1215-1999)[41]。同時(shí)在每個(gè)個(gè)體附近3個(gè)不同方向隨機(jī)選取3 個(gè)位置,去除上層枯落葉后,在土層0~20 cm 內(nèi)用土鉆取樣,將3 個(gè)土樣混合,去除大塊石頭和粗根系、草等雜物,裝入自封袋帶回實(shí)驗(yàn)室風(fēng)干,風(fēng)干后的土樣放入95 ℃烘箱烘至恒重,研磨過(guò)100 目篩,稱取100 mg 待測(cè)樣品,用錫箔紙包裹,用元素分析儀CN 模式分析測(cè)定土壤全碳量(soil total carbon content,SCC)、土壤全氮量(soil total nitrogen content,SNC)。
1.3 數(shù)據(jù)分析
相關(guān)性采用psych 包[42]中的Pearson 相關(guān)檢驗(yàn)分析。7 種性狀標(biāo)準(zhǔn)化后(減平均值,除以標(biāo)準(zhǔn)差)通過(guò)Vegan 包[43]進(jìn)行主成分分析。基于TukeyHSD 檢驗(yàn)對(duì)4種生境中的檵木幼苗主成分1(PC1)、各葉經(jīng)濟(jì)性狀、群落郁閉度進(jìn)行多重比較,并采用獨(dú)立樣本T檢驗(yàn)比較群落郁閉度在兩兩生境間的差異。對(duì)生境因子做Pearson 相關(guān)性分析后,因土壤全碳量與土壤全氮量之間相關(guān)系數(shù)大于0.5,故分析時(shí)剔除土壤全碳量。剩余自變量經(jīng)標(biāo)準(zhǔn)化后作為固定效應(yīng)因子,不同島嶼和大陸點(diǎn)做隨機(jī)效應(yīng),用lme4 包[44]和lmerTest 包[45]擬合線性混合效應(yīng)模型(最大似然估計(jì))觀察生境因子對(duì)性狀PC1 及其他各性狀的影響。通過(guò)MuMIn 包[46]完成模型選擇。保留模型權(quán)重之和達(dá)0.95 的子模型,各參數(shù)的相對(duì)重要性由其出現(xiàn)在子模型的模型權(quán)重相加得到。然后進(jìn)行模型平均得到參數(shù)估計(jì)值和95%置信區(qū)間。此外,線性混合效應(yīng)模型分析顯示,幼苗高度對(duì)性狀無(wú)顯著影響,加之高度非本研究的側(cè)重點(diǎn),故后續(xù)部分未涉及幼苗高度。
以上所有分析均于R 3.6.2 軟件[47]中完成。
2 結(jié)果
2.1 葉經(jīng)濟(jì)性狀相關(guān)性
7 個(gè)葉經(jīng)濟(jì)性狀的相關(guān)性分析顯示,21 對(duì)相關(guān)性中有12 對(duì)呈顯著相關(guān)(表1)。其中,Chl 與LDMC、C∶N 極顯著負(fù)相關(guān),而與SLA、N 極顯著正相關(guān)。LDMC 與SLA 極顯著負(fù)相關(guān),與N 顯著負(fù)相關(guān),且與C∶N 顯著正相關(guān)。SLA 與N 極顯著正相關(guān),與C∶N 極顯著負(fù)相關(guān)。N 與C 極顯著正相關(guān),與C∶N 極顯著負(fù)相關(guān)。

表1 檵木(Loropetalum chinense)幼苗不同葉經(jīng)濟(jì)性狀之間的相關(guān)性Table 1 Correlations between economic traits of Loropetalum chinense seedlings on the intraspecific level
主成分分析得到的PC1 和PC2 的累計(jì)方差貢獻(xiàn)率為62.46%(圖2)。其中,PC1 的解釋度為43.02%,該軸與Chl、SLA、N 呈高度正相關(guān),與C∶N、LDMC 呈高度負(fù)相關(guān)(表2)。PC1 體現(xiàn)了檵木幼苗的種內(nèi)葉經(jīng)濟(jì)譜,PC1 值越大,植株的資源利用越偏向于快速投資-收益策略。PC2 的解釋度為19.44 %(圖2),該軸主要與SLA 呈正相關(guān),與LDMC 和C 呈負(fù)相關(guān)(表2)。

圖2 檵木(Loropetalum chinense)幼苗葉經(jīng)濟(jì)性狀的主成分分析結(jié)果Fig.2 Results of principle component analysis of the leaf economic traits of Loropetalum chinense seedlings

表2 檵木(Loropetalum chinense)幼苗葉經(jīng)濟(jì)性狀與主成分的相關(guān)性Table 2 Correlations between economic traits and the principle components of Loropetalum chinense seedlings
2.2 葉經(jīng)濟(jì)性狀隨生境片段化的變化
多重比較結(jié)果(圖3)顯示,小島、大島邊緣以及大島內(nèi)部的檵木幼苗葉經(jīng)濟(jì)性狀PC1 顯著低于大陸。大島邊緣的檵木幼苗葉經(jīng)濟(jì)性狀PC1 顯著低于大島內(nèi)部,而小島與大島邊緣、大島內(nèi)部差異不顯著。同時(shí),大陸檵木幼苗的SLA、Chl、N 顯著高于大島內(nèi)部、大島邊緣、小島,C∶N 顯著低于大島內(nèi)部、大島邊緣、小島;大島內(nèi)部的Chl 顯著高于大島邊緣,C∶N 略低于大島邊緣(附表2)。

附表2 不同生境中檵木(Loropetalum chinense)幼苗葉經(jīng)濟(jì)性狀(平均值±標(biāo)準(zhǔn)偏差)差異Table S2 Difference of the leaf economic traits(mean ± SD)of Loropetalum chinense seedlings among different habitats

圖3 不同生境下檵木(Loropetalum chinense)幼苗葉經(jīng)濟(jì)性狀主成分1 的差異Fig.3 The difference of PC1 of Loropetalum chinense seedlings among different habitats
2.3 生境因子對(duì)葉經(jīng)濟(jì)性狀的影響
線性混合效應(yīng)模型分析顯示,檵木幼苗性狀PC1 受各因數(shù)的影響,從大到小依次為群落郁閉度、土壤全氮量、土壤最大持水量(圖4)。其中,PC1 受群落郁閉度顯著正向影響,受土壤全氮量和土壤最大持水量的影響不顯著。生境因子對(duì)各葉經(jīng)濟(jì)性狀的影響主要表現(xiàn)為郁閉度對(duì)LA、SLA、Chl 的顯著正效應(yīng)和對(duì)LDMC 的顯著負(fù)效應(yīng)(附表3)。同時(shí),大陸的群落郁閉度顯著高于大島內(nèi)部和小島,而大島內(nèi)部、大島邊緣與小島間的群落郁閉度無(wú)顯著差異(圖5)。T檢驗(yàn)結(jié)果顯示,大陸的群落郁閉度也高于大島邊緣(表3)。

表3 不同生境之間群落郁閉度對(duì)比Table 3 The comparison of canopy closure between different habitats
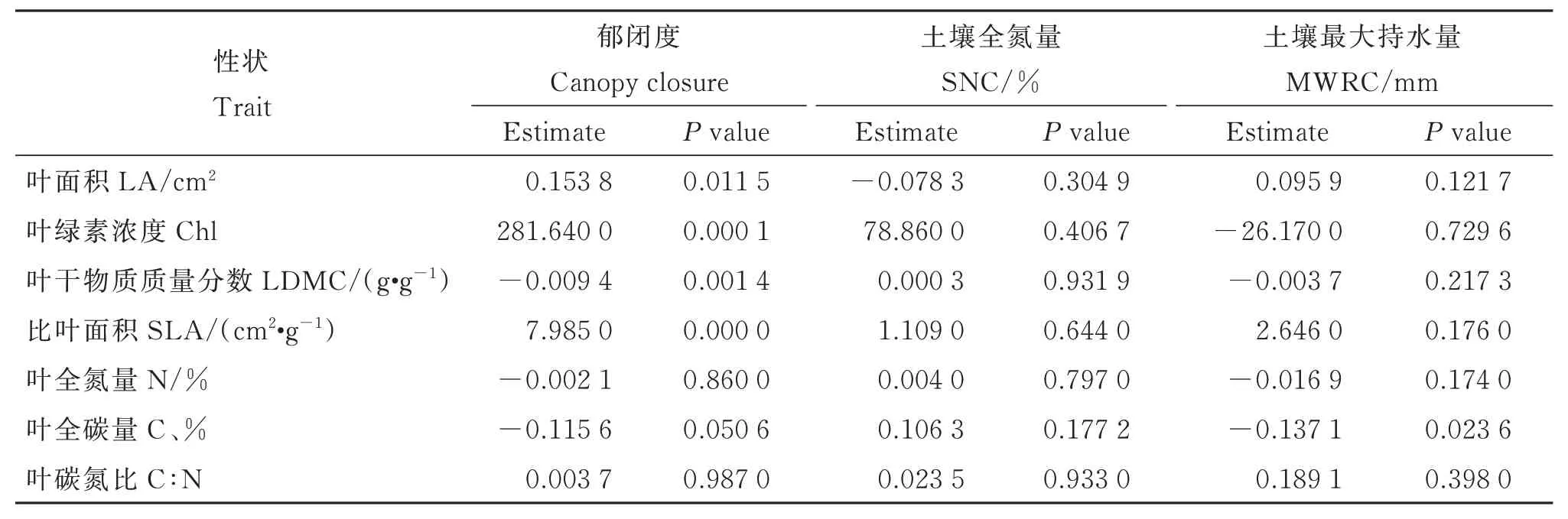
附表3 檵木(Loropetalum chinense)幼苗葉經(jīng)濟(jì)性狀與生境因子的關(guān)聯(lián)性Table S3 Correlations between leaf economic traits of Loropetalum chinense seedlings and the habitat conditions
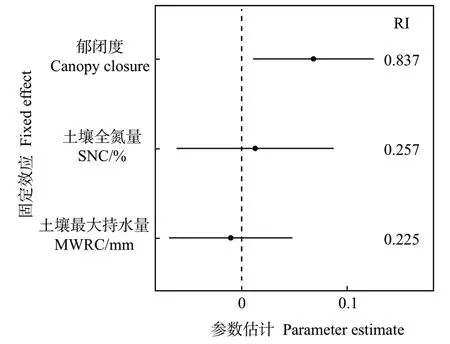
圖4 固定效應(yīng)因子的模型平均參數(shù)估計(jì)及95%置信區(qū)間Fig.4 Model averaged coefficient estimates and 95%confidence intervals for fixed effects explaining PC1

圖5 不同生境下群落郁閉度的差異Fig.5 The difference of canopy closure among different habitats(P<0.05)。
3 討論
3.1 種內(nèi)葉經(jīng)濟(jì)譜
具有高SLA 的檵木幼苗光捕獲面積大,因而具有較強(qiáng)的捕獲光資源能力[48],Chl 和N 均為光合作用的重要物質(zhì),其中Chl 是植物光合作用的主要光合色素,可在一定程度上反映植物的光合能力[39],N是光合酶不可或缺的成分,通過(guò)影響酶含量和酶活性等影響光合作用[49],高的光合速率通常伴隨高的SLA、Chl 和N[2,50],三者呈正相關(guān)關(guān)系,出現(xiàn)在葉經(jīng)濟(jì)譜的一端,代表較強(qiáng)的光合能力,即更側(cè)重于生長(zhǎng)。而高的光合速率往往伴隨短的葉片壽命,在一定程度上降低了植物對(duì)植食昆蟲(chóng)和物理脅迫的耐受能力。相反,在經(jīng)濟(jì)譜的另一端,具有較高的LDMC、C∶N。LDMC 高的葉片相對(duì)堅(jiān)韌,對(duì)植食動(dòng)物的適口性較低,且對(duì)物理脅迫的抵抗力更強(qiáng)[51]。葉片中C∶N 較高表明其具有較多的防御物質(zhì)(如纖維、固醇類(lèi))和較少的營(yíng)養(yǎng)成分(如蛋白質(zhì)等)。有研究表明,資源利用策略較保守的植物,通常具有較長(zhǎng)的葉片壽命和較低的光合速率,即更側(cè)重于生存[1]。
檵木幼苗種內(nèi)葉經(jīng)濟(jì)性狀變異間的顯著協(xié)同性,主要呈現(xiàn)為PC1,表明存在一定程度的種內(nèi)葉經(jīng)濟(jì)譜。具體地,PC1 與各性狀的關(guān)系表明,PC1 越大,則SLA 大、N 和Chl 高,而LDMC 和C∶N 減少,植物將資源更多地用于光合生產(chǎn),以使其快速生長(zhǎng),即具有更大的最大光合效率,更傾向于快速投資-收 益的資源利用策略;PC1 越小,則SLA 小、N 和Chl 低,而LDMC 和C∶N 增大,更有利于幼苗葉片對(duì)外界的防御,可能也具有更長(zhǎng)的葉壽命,傾向于緩慢投資-收益的資源利用策略[2,23,52]。與以往研究相比[15,19],本研究檢測(cè)出了種內(nèi)葉經(jīng)濟(jì)性狀間相對(duì)較強(qiáng)的關(guān)聯(lián)性,這可能與生境片段化帶來(lái)的較大的生境異質(zhì)性有關(guān)。
3.2 片段化對(duì)生境條件及種內(nèi)葉經(jīng)濟(jì)性狀的影響
千島湖庫(kù)區(qū)的生境片段化,對(duì)馬尾松林內(nèi)生境條件和檵木幼苗葉經(jīng)濟(jì)性狀具有一定影響。片段化可能通過(guò)影響生境條件進(jìn)而影響幼苗性狀。相對(duì)于大陸森林,島嶼森林內(nèi)的檵木幼苗更傾向于采取緩慢投資-收益的資源利用策略。群落郁閉度對(duì)檵木幼苗最大光合效率具有顯著正效應(yīng),大陸森林的林下光照較少,檵木幼苗具有較大的SLA 和Chl 等性狀特征,以提高對(duì)光照的利用效率,達(dá)到更高的同化率,這與以往的研究結(jié)果一致[27]。相反,島嶼森林的林下光照更充足,可能也伴隨較大的溫度浮動(dòng)等不利生境條件,在此情況下,較小的SLA(較厚的葉片)有利于減少水分散失等以抵御高溫干旱脅迫。這一系列的性狀特征改變,導(dǎo)致島嶼生境中的檵木幼苗形成了較為保守的資源利用策略,葉片可能較為結(jié)實(shí)耐久[38],進(jìn)而更好地適應(yīng)片段化環(huán)境,在經(jīng)濟(jì)譜中偏向于緩慢投資-收益的資源利用策略。
以上結(jié)果符合葉經(jīng)濟(jì)譜中的環(huán)境因子主導(dǎo)假說(shuō)[23]。
島嶼中,大島內(nèi)部的最大光合效率更高,表明大島內(nèi)部的檵木幼苗更傾向于快速投資-收益的資源利用策略,而大島邊緣更傾向于緩慢投資-收益的資源利用策略。這可能與檵木幼苗對(duì)昆蟲(chóng)植食的防御有關(guān)。GONZALEZ 等[53]在探究片段化森林中邊緣效應(yīng)對(duì)節(jié)肢動(dòng)物的影響時(shí)指出,邊緣的植食昆蟲(chóng) 豐富度更高;GUIMARAES 等[54]通過(guò)整合分析片段化對(duì)植食昆蟲(chóng)多度、豐富度和植食的作用指出,森林邊緣具有較高的植食昆蟲(chóng)多度和豐富度,且邊緣植物比內(nèi)部植物的植食率高70%;而MALDONADO-LOPEZ 等[55]也發(fā)現(xiàn),森林邊緣的癭蜂具有較高的多度和豐富度。大島邊緣的檵木幼苗可能更多地將資源分配到防御性狀上,為更好地存活而采取緩慢投資-收益的資源利用策略。小島與大島間檵木幼苗葉經(jīng)濟(jì)性狀差異不顯著,表明邊緣效應(yīng)可能較面積效應(yīng)對(duì)植物葉經(jīng)濟(jì)性狀的影響更大。
與以往研究不同[25,56],土壤全碳量、土壤全氮量和水分條件對(duì)千島湖片段化島嶼及周邊連續(xù)森林中的檵木幼苗性狀無(wú)顯著影響,這可能與千島湖片段化景觀基質(zhì)是水有關(guān),使水分不再是限制性因子。而HAN 等[57]通過(guò)對(duì)753 種植物的氮磷比研究指出,我國(guó)陸地植物生長(zhǎng)主要受土壤磷的限制,這可能是導(dǎo)致檵木幼苗葉經(jīng)濟(jì)性狀不受土壤全碳量、全氮量影響的原因。
4 結(jié)論
檵木幼苗種內(nèi)葉經(jīng)濟(jì)性狀并不獨(dú)立存在,面對(duì)片段化森林中的多變生境,檵木在更新階段形成了一套能適應(yīng)特定環(huán)境的葉經(jīng)濟(jì)性狀組合,從而調(diào)整其資源利用策略,形成了種內(nèi)葉經(jīng)濟(jì)譜。片段化可能通過(guò)影響生境條件進(jìn)而影響植物的葉經(jīng)濟(jì)性狀,而隨著生境片段化的加劇,檵木幼苗葉逐漸傾向于緩慢投資-收益策略。本研究一定程度上豐富了植物葉經(jīng)濟(jì)譜的理論,在開(kāi)展基于功能性狀的物種共存研究時(shí)對(duì)是否需要考慮種內(nèi)變異具有一定的指導(dǎo)作用。也提示在研究片段化生境中群落構(gòu)建和生物多樣性維持機(jī)制時(shí)應(yīng)考慮種內(nèi)葉經(jīng)濟(jì)譜的影響。
后續(xù)研究需同時(shí)測(cè)定葉脈密度、氣孔密度等水力性狀以確定主成分2 是否與植物的水分利用策略等相關(guān),且需同時(shí)考慮生物因子對(duì)葉經(jīng)濟(jì)性狀的影響;并在條件允許的情況下盡可能增大取樣量,以使結(jié)果更有說(shuō)服力。
貴州師范大學(xué)金毅,浙江大學(xué)張愛(ài)英、巫東豪對(duì)論文提出了修改建議,淳安縣新安江生態(tài)開(kāi)發(fā)集團(tuán)有限公司在項(xiàng)目實(shí)施過(guò)程中給予配合與支持,當(dāng)?shù)卮迕駥?duì)野外工作給予幫助,在此一并致以誠(chéng)摯的謝意!
猜你喜歡
今日農(nóng)業(yè)(2022年14期)2022-09-15 01:44:56
民生周刊(2020年13期)2020-07-04 02:49:22
華人時(shí)刊(2018年23期)2018-03-21 06:26:00
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
西部大開(kāi)發(fā)(2017年7期)2017-06-26 03:14:00
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
大社會(huì)(2016年6期)2016-05-04 03:42:05
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
 浙江大學(xué)學(xué)報(bào)(理學(xué)版)2021年6期
浙江大學(xué)學(xué)報(bào)(理學(xué)版)2021年6期
- 浙江大學(xué)學(xué)報(bào)(理學(xué)版)的其它文章
- 中國(guó)出境旅游的外交效應(yīng)
- 1980—2018 年鎮(zhèn)賚縣濕地景觀格局變化與人為干擾的關(guān)系
- 中國(guó)自然災(zāi)害時(shí)空變化及其社會(huì)經(jīng)濟(jì)效應(yīng)
- 中國(guó)歷史災(zāi)害研究的特點(diǎn)與態(tài)勢(shì)管窺
——基于博、碩士學(xué)位論文的計(jì)量分析 - 基于NPP-VIIRS 夜間燈光數(shù)據(jù)的山西省GDP 空間化模擬
- 基于ISCCP 數(shù)據(jù)集的梅雨期影響浙江省及周邊地區(qū)的對(duì)流系統(tǒng)特征分析