基于粒子圖像測速技術(PIV)的自由游泳草魚動力學特征分析
2021-11-17 04:29:40楊國黨胡曉張奔龍澤宇張永年石小濤
大連海洋大學學報 2021年5期
關鍵詞:效率
楊國黨,胡曉,張奔,龍澤宇,張永年,石小濤
(三峽大學 水利與環(huán)境學院,湖北省魚類過壩技術國際科技合作基地,湖北 宜昌 443002)
相關研究成果和結論可對魚類物理模型及仿生機器魚研究提供科學參考。
許多水生動物,特別是魚類,通常會通過改變游泳狀態(tài)來適應環(huán)境變化,這種現(xiàn)象引起了科學界廣泛的關注。魚類能夠準確感應身體周圍的流動,通過將身體運動的動量傳遞給周圍的流體,從而有效地控制推力的產(chǎn)生[1]。對于魚類來說,它們傳遞給周圍流體的動量和能量會以漩渦的形式包裹起來,研究者通過量化流體中的循環(huán)量,從而估計魚體產(chǎn)生的推力和推進效率[2-4]。
魚類在水體中運動時的瞬態(tài)流場變化是分析魚類游泳動力形成的關鍵因素,而粒子圖像技術(particle image velocimetry,PIV)能在不影響魚類運動的情況下顯示瞬態(tài)流場整體結構并保證測量的分辨率和精度[5]。近年來,隨著科技的不斷發(fā)展,PIV應用也愈發(fā)普遍,Stamhuis等[6]在測量鯔魚幼魚周圍流場時首次引入該技術,PIV技術開始成為魚類運動力學研究中的重要工具,并逐漸為研究者所接受。
目前,國內(nèi)外學者主要通過對活魚進行生物試驗,以及對仿生魚模型進行物理試驗及數(shù)值模擬等手段研究魚類游泳的運動力學機理。Tytell等[7]分析了美洲鰻鱺在穩(wěn)定游動下的尾流結構并測量了其推進效率;Mwaffo等[8]研究了斑馬魚Daniorerio在機動和巡游兩種運動模式下的渦結構和水動力壓力分布;Akanyeti等[9]通過仿生學模型試驗,驗證了加速期間魚尾部通過改變渦環(huán)幾何結構來提高推進效率;Borazjani等[10]通過數(shù)值模擬研究了雷諾數(shù)和尾拍頻率系統(tǒng)變化下鰻魚運動的水動力學;Kazakidi等[11]通過數(shù)值模擬發(fā)現(xiàn)尾鰭的形狀會影響魚體的推進效率。研究魚類在不同游泳狀態(tài)下的推進機制與動力學不僅能為高效率的仿魚類推進裝置設計提供參數(shù)依據(jù),還對理解水生動物的進化過程及其生物學作用具有一定的科學價值。前期相關研究大多集中于魚類常見的游泳狀態(tài)(如直線等),而對魚類轉彎和后退游泳狀態(tài)的研究較少。本研究中,采用PIV試驗結合渦量分析原理對草魚Ctenopharyngodonidellus幼魚在直線、轉彎及后退3種游泳狀態(tài)下的受力和推進效率進行了研究,分析了草魚幼魚在不同游泳狀態(tài)轉變過程中魚體周身的渦量分布特征,通過對正向力和反向力的定量分析,得到了上述3種游泳狀態(tài)下幼魚高效的行為方式,
1 材料與方法
1.1 材料
試驗用魚為常見的淡水魚草魚幼魚,購自湖北省宜昌市某養(yǎng)殖場,共15尾,平均體長為4~10 cm,暫養(yǎng)在圓形養(yǎng)魚池中,水溫保持在(20±1)℃,每周投喂兩次,試驗前24 h禁止攝食[12]。
試驗裝置如圖1所示,包括一套PIV系統(tǒng)(光源系統(tǒng)、同步系統(tǒng)、圖像拍攝系統(tǒng)、圖像分析系統(tǒng))和透明的有機玻璃水槽(長×寬×高為0.46 m×0.20 m×0.25 m)。

圖1 試驗裝置圖
1.2 方法
1.2.1 試驗設計 試驗于三峽大學生態(tài)水利學實驗室進行,試驗時水槽內(nèi)水深為0.1 m,示蹤粒子選用隨水性良好的空心玻璃微珠(粒徑為20~30 μm),在水槽前后兩側分別安裝激光器(型號為NQ-506,波長為532 nm,功率為100 mW)1和2,在四周分別架設一部高速攝像機(Redlake Motionscope,分辨率為640 pixel×480 pixel,幀率為120 fps),視場范圍為12 cm×9 cm。整個試驗在無光和無外界干擾的條件下進行,為了防止粒子圖像可見度減弱和兩側氬離子激光器發(fā)生散射,將一塊貼黑紙的薄玻璃隔板放在試驗區(qū)域的正下方。試驗選用澄清的自來水(試驗前靜置48 h),通過攝像機捕捉草魚幼魚在靜水環(huán)境下的自主前進、轉彎及后退行為,每條魚單獨進行試驗,每次試驗持續(xù)時間為30 min。根據(jù)實驗生物學要求,同一試驗魚不再重復試驗,避免對試驗結果造成影響[13]。
1.2.2 數(shù)據(jù)處理 試驗獲得的草魚運動圖片采用開源程序PIVlab獲取原始的流速場(有完整魚體),提取魚體周身的渦量和速度,導入Surfer軟件中將整個流場進行細分,獲得渦環(huán)在二維水平面上的投影面積及渦量,推導計算各部分的作用力(利用Matlab對一個擺尾周期內(nèi)的魚體周身進行作用力計算)。由于魚體在5次擺尾后尾拍幅度和游動速度不斷下降,直到第7次擺尾后,草魚幼魚保持身體筆直,開始滑行,所以本研究中選取1到5次擺尾間的數(shù)據(jù)進行數(shù)字化圖像處理[14]。為便于分析魚的運動,定義草魚幼魚向前波動的過程中,魚從幾乎順直狀態(tài)開始擺尾一個來回,這個過程的歷時定義為一個周期T,選取的直線、轉彎和后退游泳期間完成一個擺尾的運動周期T分別為300、575、400 ms。整個過程由于粒子數(shù)據(jù)較多,前期主要通過人工手段提取魚體周身的粒子,用于具體分析每一部分受力變化,本試驗中結合Triantafyllou等[15]的研究理論,通過周身渦流的變化強度將魚體劃分為3部分,如圖2 所示,將吻端至胸鰭間的區(qū)域定義為頭部,將胸鰭至臀鰭間的區(qū)域定義為中部,將臀鰭至尾鰭后部的區(qū)域定義為尾部,然后分析每一幀下不同部位數(shù)據(jù)的變化,由于數(shù)據(jù)過多,每張粒子圖像下的數(shù)據(jù)變化較小,故將其在時間上進行劃分。
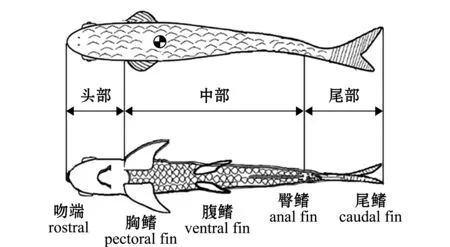
圖2 魚體部位劃分示意圖
1.2.3 相關力學計算原理 魚體的每次波動都會產(chǎn)生渦環(huán),通過渦環(huán)可以獲取施加于魚體上的作用力。首先,獲取不同粒子的速度大小及分布,計算魚體周身渦量分布和沿渦環(huán)的速度環(huán)量,其計算公式[16]為

其中:Ω為渦量(1/s);u、v分別為粒子的水平速度和豎直速度(m/s);Γ為速度環(huán)量(m2/s);v′為粒子的速度(m/s);l為渦環(huán)的周長(m)。
根據(jù)單位時間速度環(huán)量面積的變化,作用在幼魚魚體上的瞬時作用力計算公式[17]為

(3)
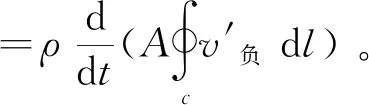
(4)
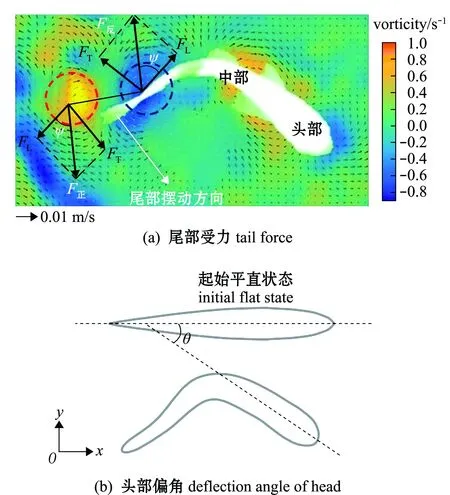
圖3 魚體受力示意圖
由于魚體的每次波動都會產(chǎn)生作用力,每次產(chǎn)生的作用力又會隨射流角度的變化而發(fā)生改變,根據(jù)其力的作用效果不同,可將其分為沿魚體切向方向的推力(FT)及法向方向的側向力(FL),并通過渦環(huán)在不同平面上的投影面積計算相應的作用力,公式[19]可以改寫為
其中:Axy為渦環(huán)在xy平面上的投影面積(m2);Ayz為渦環(huán)在yz平面上的投影面積(m2);Γ為速度環(huán)量(m2/s);當魚體波動前行時,尾部通常伴隨有大振幅,頭部會存在一定的小振幅,頭部在游泳時大致是剛性的[20],因此,幼魚每一時刻頭部中線與起始平直狀態(tài)中線間都會形成一定的夾角θ,定義為頭部偏角[8,21](圖3(b))。
通過計算流體動力學得到魚類游泳的推進效率(η),其計算公式[22]為
η=FTuaxial/(FTuaxial+Plateral),
(7)
Paxial=FLulateral。
(8)
其中:FT為推力(N);FL為側向力(N);uaxial為幼魚的水平游泳速度(魚體質(zhì)心的水平位移與相應時間的比值,m/s);ulateral為幼魚的側向游泳速度(魚體質(zhì)心的豎直位移與相應時間的比值,m/s);Plateral為幼魚側向運動損耗的功率(W);Paxial為幼魚水平向運動損耗的功率(W)。
2 結果與分析
2.1 不同游泳狀態(tài)下草魚幼魚周身的受力變化
2.1.1 直線游泳狀態(tài)下的受力變化 結合圖4的云圖和圖5的曲線可以看出,草魚幼魚完成一個擺尾周期(T)的直線游動過程,魚體周身的正、負渦作用力之比可達1.6∶1(圖5(a)),其中幼魚尾部主要聚集正渦,頭部聚集負渦(圖4),上述兩個部位一個運動周期內(nèi)所形成的正、反向力平均比例分別為43%和34%,如圖5(d)、(b)所示。

圖4 直線游泳時魚體渦量云圖
在運動的前1/8周期,幼魚尾部是正渦的主要聚集區(qū)域,該階段尾部由正渦所產(chǎn)生的作用力占魚體總正向力的比例接近60% (圖5(d)),之后正渦逐漸向魚體胸鰭部位聚集,魚體中部正向力的比例開始上升并在1/2周期達到最高點,如圖5(c)所示,尾部由正渦產(chǎn)生的作用力開始下降,此后正渦逐漸由中間部位向尾部擴散。在整個直線游泳過程中,魚體有63%(33.74 mN)的正向力被抵消不做功,只有37%(19.88 mN)的正向力對魚體做功,其中大部分集中在魚體尾部(圖5(a))。

圖5 直線游泳時魚體各部分受力
2.1.2 轉彎游泳狀態(tài)下的受力變化 草魚幼魚在轉彎時,魚體的彎曲方向與魚頭轉彎的方向剛好相反,幼魚的擺尾幅度增大,擺尾頻率降低,試驗結果顯示,在頭部偏角相同的情況下,轉彎時草魚幼魚前進的距離要遠小于直線游泳狀態(tài)。
從圖6和圖7(a)可知,幼魚完成一個周期的轉彎游動過程,魚體周身的正、負渦作用力之比為2.2∶1,整個轉彎過程中,有45%(29.4 mN)的正向力被反向力所抵消不做功,實際作用于魚體的力只有55%(36.3 mN),其中,尾部為正渦的主要聚集區(qū)域,一個運動周期內(nèi)由尾部正渦所形成的正向力占總正向力的比例平均為55%。
圖6的渦量云圖顯示,在運動的前1/16周期,幼魚頭部聚集的正、負渦呈上升趨勢,此時頭部渦體產(chǎn)生的正向力所占比例高達60%(圖7(b)),之后正渦逐漸向魚體中部和尾部擴散;在1/2周期時刻,幼魚魚體的彎曲程度最大,魚體尾部的正向力、反向力所占比例分別達到最高值64%、80%(圖7(d)),而魚體中部則聚集了較多負渦(圖7(c)),該部位由負渦所形成的反向力比例平均可達32%。

圖6 轉彎游泳時魚體渦量云圖

圖7 轉彎游泳時魚體各部分受力
2.1.3 后退游泳狀態(tài)下的受力變化 草魚的后退游泳行為,在生物力學上表現(xiàn)出不同差異(當草魚遇到狹小空間或無法繼續(xù)前進時,就會出現(xiàn)后退游泳行為)。整個后退游泳過程中渦量云圖如圖8所示,渦量首先在幼魚的頭部聚集,然后沿魚體長度方向逐漸向幼魚的中部、尾部擴散,之后再次聚集和擴散,導致幼魚各部位的正向力、反向力呈現(xiàn)出周期性的波動。如圖9(a)所示,草魚幼魚完成一個周期的后退游動過程,幼魚尾部聚集正渦和負渦的區(qū)域逐漸減小;相對直線游泳和轉彎狀態(tài)而言,魚體尾部的正向力和反向力所占比例有一定程度下降(圖9(d));魚體中部主要聚集負渦,該部位在一個運動周期內(nèi)產(chǎn)生的反向力所占比例平均可達41%(圖9(c));雖然幼魚頭部的正向力、反向力波動較大,但其平均值仍維持在較高水平(圖9(b))。
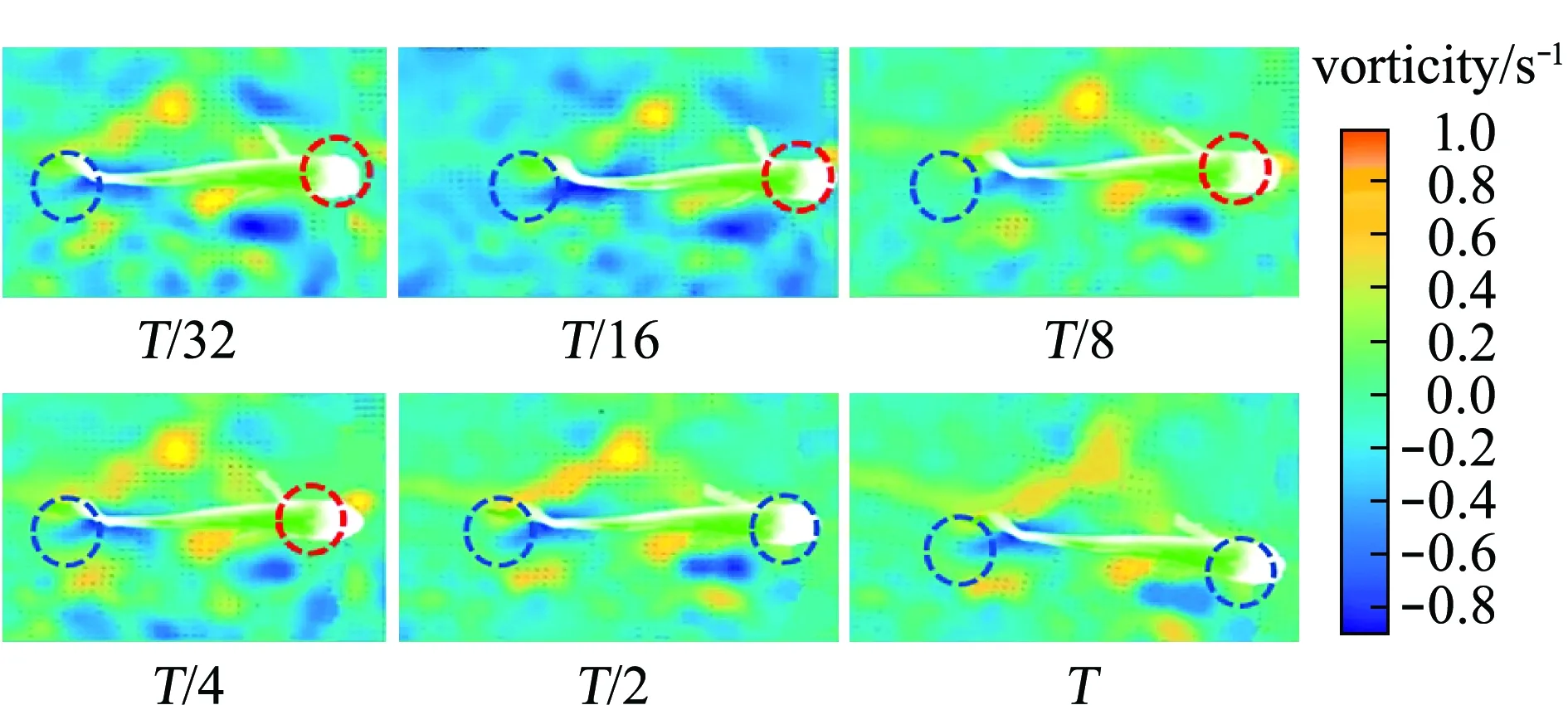
圖8 后退游泳時魚體渦量云圖

圖9 后退游泳時魚體各部分的受力
2.2 不同游泳狀態(tài)下草魚幼魚的推力和側向力
通過對試驗數(shù)據(jù)進行分析發(fā)現(xiàn),草魚幼魚在直線游泳狀態(tài)時,有高達73%的正、負渦合作用力轉化為推力,且該推力主要產(chǎn)生在幼魚尾部,剩下的渦合作用力則轉化為側向力,主要用于尾部的橫向擺動和頭部的偏轉;轉彎游泳的幼魚將52%的渦合作用力轉化為推力,相較于直線游泳,幼魚頭部推力的比例在逐漸降低,僅為直線游泳的50%,此外,由于轉彎時草魚幼魚的整體偏轉角度增大,使得魚體的推力分量大幅下降(圖10(a)、(b));相對前兩種游泳狀態(tài),后退游泳產(chǎn)生的作用力相對較小,其中有66%的渦合作用力轉化為推力,其余的轉化為側向力,雖然推力的利用率整體增大,但由于正、負渦的合作用力相對較小,故后退時的幼魚推力較小(圖10(a)、(b))。

圖10 不同游泳狀態(tài)下魚體周身的推力及側向力
2.3 不同游泳狀態(tài)下草魚幼魚的推進效率
本研究中結合草魚幼魚周身的受力特征,對其頭部、中間部位和尾部的推進效率進行了分析比較,結果如圖11(a)~(c)所示,發(fā)現(xiàn)直線游泳和轉彎時的幼魚尾部推進效率最高,其次是中部和頭部;當幼魚后退狀態(tài)時,魚體的高效推進在中部,頭部推進效率最低。
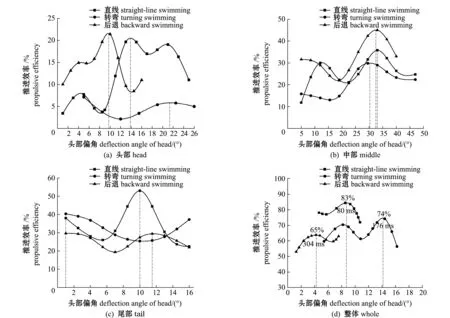
圖11 草魚幼魚不同部位的推進效率
通過對幼魚直線、轉彎和后退3種狀態(tài)下的整體推進效率比較發(fā)現(xiàn):直線游泳的幼魚整體推進效率最高,其次分別是轉彎和后退游泳狀態(tài);當幼魚的頭部偏角為9°~10°時,直線游泳的整體推進效率最高,為83%;對于轉彎的草魚幼魚,其整體推進效率在頭部偏角為14°~15°時最高,為74%;后退游泳時,草魚幼魚的頭部偏角為3°~4°時,其整體推進效率達到最大值65%。
3 討論
3.1 不同游泳狀態(tài)下的動力學差異
在不同的水環(huán)境中,魚類為了特定目的進化出各種各樣的運動策略。魚類通過肌肉變形作用于流體,帶動周圍流體的運動,同時受到流體的反作用力而推動自身前進[23]。Müller等[24]研究發(fā)現(xiàn),魚體周身渦結構是沿體表下移直至尾部脫落,這與本研究中的分析結果一致。一些研究發(fā)現(xiàn),魚體整個身體對推力產(chǎn)生都有一定貢獻,另有研究提到尾鰭在產(chǎn)生推力方面的重要性。當尾鰭產(chǎn)生的渦強度較大,其產(chǎn)生的推力更大[25]。本研究中發(fā)現(xiàn),草魚幼魚在近似直線運動過程中,正、負渦合作用力對推力的貢獻作用高達73%,且該推力主要產(chǎn)生在幼魚尾部。相比之下,草魚幼魚轉彎時有較高的推力和側向力,但是其轉彎時的整體偏轉角度增大,使得正、負渦合作用力轉化成推力的分量大幅度下降。另外,魚體轉彎時脫落的渦環(huán)的動量可以促使魚加速運動[26]。本研究中發(fā)現(xiàn),草魚幼魚后退時主要靠胸鰭劃動后退,魚體周身渦量變化主要與其中部的胸鰭劃動有關,相比直線、轉彎時魚體借助尾部的主要動力來源,魚體后退過程中正、負渦的合作用力整體相對較小,因此,后退時呈現(xiàn)較小的推力。由于魚體和流體相互作用的復雜性,未來應開展三維渦量研究,進一步闡明草魚幼魚產(chǎn)生的尾流渦結構,這有助于對魚尾渦結構進行分類和作用機理的認識。
3.2 魚體頭部偏轉角度對推進效率的影響
在自由游泳中,魚體有角度的偏轉與流體的相互作用對其整體推進效率有重要的影響[27]。魚類通過周期性激活肌肉來控制頭部偏轉角度,以補償側滑加速[28]。這說明魚體頭部壓差降至最低時,可減少流體阻力,并提高推進效率。另外,Nair等[27]研究發(fā)現(xiàn),斑馬魚幼魚尾部偏轉角度與頭部偏角呈線性關系。這說明魚體頭部控制對于通過轉向、身體旋轉和側向平移來提高游泳效率至關重要[29]。本研究中發(fā)現(xiàn),3種不同游泳模式下魚頭偏轉角度不盡相同,與轉彎頭部偏轉角度相比,直線游泳頭部偏角為9°~10°時就可以達到83%的推進效率。這說明轉彎頭部偏轉幅度增加并不意味著渦環(huán)產(chǎn)生的動力成本低于直線游泳產(chǎn)生的動力成本。這主要是由于試驗幼魚轉彎時尾部渦動力產(chǎn)生與游泳方向相反的力,需要較大外力矩實現(xiàn)轉彎,最終導致游泳速度大大降低,因此,運動效率相對較低。張冰冰[30]也發(fā)現(xiàn),魚體轉彎比“S”型波動前進會產(chǎn)生較小的力矩和推力,故前進角度大但推進效率小。實際中,除了魚體頭部偏轉以外,魚體常表現(xiàn)出空間俯仰角度和橫搖角度變化,需進一步研究魚體這些行為引起渦動力的變化情況。
3.3 不同游泳狀態(tài)下的推進效率差異
推進效率是魚類動力學研究的重要指標, 是衡量魚類游泳能力的主要評判標準之一。王睿等[31]對仿生魚研究發(fā)現(xiàn),仿生魚的推進效率遠低于自然界中的魚類,其原因在于仿生魚的尾流結構并未根據(jù)身體其他部位的變化進行整體性協(xié)調(diào)。自然界的鯉科魚類,魚游泳前期主要是通過尾部產(chǎn)生推力,后期利用頭部和胸鰭的偏轉來抵抗周圍水體阻力,而仿生魚只完成了魚體前期運動,這也是仿生機器魚的水力學特性高于實際魚體,但游泳效率卻低于實際魚的主要原因。本文結合草魚幼魚的游泳速度對其推進效率進行研究,可以得到不同游泳狀態(tài)下幼魚的最佳游泳方式。
本研究中發(fā)現(xiàn),在直線和轉彎中魚體尾部相比頭部和中部具有較高的推進效率,而兩者隨體長變化不盡相同。這也證明了魚體尾部是其前進的主要動力來源。Chen等[32]研究魚在流體中高效推進機理時發(fā)現(xiàn),推進效率隨著游動波速的升高而增大。
本試驗中幼魚在后退時中部推進效率貢獻高于頭部和尾部,推測可能與魚體中部波速有關。Wardle等[33]通過研究魚體體波時序和肌電圖(electromyogram,EGM)的區(qū)別時發(fā)現(xiàn),肌肉時斷時續(xù)產(chǎn)生額外能量,部分時間做負功甚至不做功,這可能是魚類具有高推進效率的原因。實際中,在具有來流的條件下,魚類常表現(xiàn)出頂流前進、頂流后退、趨流轉換等行為,因此,需進一步研究以提高對魚類推進機理方面的認識。
4 結論
本研究中利用PIV技術結合渦量理論定量分析了草魚幼魚在直線、轉彎及后退游泳狀態(tài)下的動力學特征,主要得出以下結論:
1)草魚幼魚在直線和轉彎游泳時主要依靠尾部聚集的正渦產(chǎn)生推力,而后退時則主要依靠魚體中間部位的負渦形成反向力,來推動自身前進。
2)對不同游泳狀態(tài)下草魚幼魚所受推力和側向力分析發(fā)現(xiàn),直線游泳的幼魚推力轉化率最高可達73%,轉彎時的正、負渦合作用力有48%用于頭部和胸鰭部位的偏轉,后退時的幼魚渦合作用力轉化率為66%,但是對應的推力卻相對較小。
3)對草魚幼魚不同游泳狀態(tài)下的整體推進效率進行對比分析得到,直線游泳的幼魚整體推進效率最高可達83%,其次是轉彎和后退游泳狀態(tài),幼魚最高推進效率分別為74%和65%,試驗數(shù)據(jù)顯示,較小的頭部偏角更有利于幼魚的高效推進。
猜你喜歡
瘋狂英語·初中天地(2021年5期)2021-07-21 02:24:28
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數(shù)理化(高中版.高考數(shù)學)(2020年5期)2020-06-02 09:19:08
商周刊(2017年9期)2017-08-22 02:57:49
遼寧經(jīng)濟(2017年6期)2017-07-12 09:27:16
中國衛(wèi)生(2016年9期)2016-11-12 13:27:54
時代英語·高二(2015年1期)2015-03-16 00:08:11
中國洗滌用品工業(yè)(2015年7期)2015-02-28 19:02:38
電子設計工程(2015年12期)2015-02-27 12:06:10
中國衛(wèi)生(2014年11期)2014-11-12 13:11:32
