基于測序基因分型技術的3種石斛蘭遺傳關系分析
2021-11-10 08:28:12王趁芳黃守程周玉麗王曉鵬劉愛榮侯北偉葉梅榮
中國野生植物資源 2021年10期
關鍵詞:分析
王趁芳,黃守程,周玉麗,王曉鵬,劉愛榮,侯北偉,葉梅榮*
(1.安徽科技學院,安徽鳳陽233100;2.南京野生植物綜合利用研究所,江蘇南京211100)
測序基因分型技術(Genotyping by sequencing,GBS)是基于二代測序技術的簡化基因組測序,可以檢測大量單核苷酸(Single nucleotide polymor?phisms,SNP)變異位點,并且成本低[1]。SNP為第三代分子標記、具有密度高、分布范圍廣、遺傳穩定、分型簡單、可自動化等優點[2],SNP多態性是指由于單個核苷酸變異所引起的基因組上DNA序列的多態性[3]。目前GBS-SNP標記已廣泛用于動植物群體的進化、育種、親緣關系、遺傳結構、物種鑒定等研究[4]。如周萍萍等[5]利用該標記研究栽培六倍體燕麥(Avena sativaL.)的起源;Talaveraet al.[6]應用該標記研究牛油果(Persea americanaMill.)的遺傳多樣性和群體結構。
霍山石斛(Dendrobium huoshanenseC.Z.Tang and S.J.Cheng)、鐵皮石斛(Dendrobium officinaleKimura et Migo)和 細 莖 石 斛(Dendrobium monili?forme(L.)Sw.)均為蘭科(Orchidaceae)石斛屬(Den?drobiumSw.)多年生附生草本植物,是我國傳統名貴的藥用植物[7]。霍山石斛俗稱米斛、霍石斛等,僅分布在我國大別山區,如湖北英山、河南南召、安徽霍山等地區[7];而鐵皮石斛和細莖石斛分布范圍廣泛,從我國云南最南部的熱帶到北部的河南省均有分布[8-9]。霍山石斛的莖桿加工成的楓斗被稱為“金霍斗”,為“楓斗之王”;而鐵皮石斛和細莖石斛的莖桿被加工成著名的中藥“鐵皮楓斗”和“銅皮楓斗”[10-11]。中國約有40多種石斛作為傳統的中藥藥材,具有重要的藥用和保健功能,據《神農本草經》記載“石斛,一名林蘭。味甘,平,無毒。主傷中,除痹,下氣,補五藏虛勞贏瘦,強陰。久服厚腸胃,輕身延年”[12]。現代藥理研究表明,石斛具有養胃潤肺、清音明目、增強免疫力、抗氧化、抗腫瘤、降血糖、防衰老和延年益壽等功效[13]。因此石斛楓斗被當作高檔保健飲品,價格昂貴,市場需求極大。雖然有近40多種石斛被作為藥用植物,而歷代本草及現在被公認的優質“石斛”應該指“鐵皮石斛”與“霍山石斛”[14]。目前關于霍山石斛、鐵皮石斛與細莖石斛的遺傳關系及其鑒別的研究報道已有不少,如羅宇琴等[15]從含量測定、指紋圖譜、藥理作用等方面分析霍山石斛和鐵皮石斛的異同。這3種石斛的遺傳關系一直是石斛研究者感興趣的問題,因此該文基于GBS標記的SNP位點研究3種石斛的遺傳關系,為進一步促進3種石斛的研究提供參考。
1 材料和方法
1.1 試驗材料
該研究于2019年9月在安徽霍山地區只收集到栽培3年以上的霍山石斛樣本8份和栽培鐵皮石斛樣本7份。因霍山石斛、鐵皮石斛及細莖石斛的野生樣本稀缺,所以未能收集到霍山石斛和鐵皮石斛的野生樣本。而細莖石斛除收集到霍山地區栽培的3份樣本外,又收集到2份廣西偉江的野生樣本。所有石斛樣本均由王曉鵬教授鑒定并保存在安徽科技學院里。將樣本的新鮮葉片收集于密封袋,然后迅速投入液氮里速凍,取出密封袋埋藏于干冰里,運送到菲莎基因有限公司進行DNA提取和GBS分析。
1.2 DNA提取、文庫構建和GBS分析
用改良的十六烷基三甲基溴化銨法(Cetyltri?methylammonium bromide,CTAB)提取樣本的DNA,利用瓊脂糖凝膠電泳檢測樣品DNA的完整性及DNA片段長度,DNA純度用Nanodrop微量分光光度計檢測,DNA的濃度用Qubit 3.0精確定量,只有單個樣本的DNA濃度≧10μg才能用于文庫構建;將檢測合格的DNA樣品用Covaris超聲波破碎儀隨機打斷,用Msel酶降解,末端修復后加A尾和測序接頭、純化、聚合酶鏈式反應(Polymerase chain reaction,PCR)擴增等步驟完成構建整個文庫,用Illumina測序儀PE150對構建好的文庫進行測序。
1.3 序列處理和比對到參考基因組
測序平臺獲得的原始圖像數據文件用堿基識別分析轉化為原始測序序列即Raw data[所有Raw data數據已上傳美國國家生物技術信息中心(Na?tional center for biotechnology information,NCBI),登錄號為PRJNA659117],用Trimmomatic Version 0.38軟件對Raw data過濾,去除含有接頭、兩端連續質量小于20堿基對(base pair,bp)及堿基長度小于50 bp的測序片段,成對的測序片段被保留,即最后獲得干凈的測序片段稱為clean data或clean reads。用Burrows-wheeler alignment tool(BWA)[16-17]將clean data比對到黃石斛(Dendrobium catenatumLindl.)[18]參考基因組上。
1.4 變異檢測
Clean data的SNP數目用The genome analysis toolkit(GATK)[19]軟件進行檢測和過濾,Minor allele frequency(MAF)<0.01和20%樣本沒有的SNP被去除掉,得到的SNP可用作遺傳結構等分析。
1.5 居群的遺傳結構特征和遺傳距離分析
居群遺傳結構用Admixture軟件[20]進行分析,交叉驗證誤差最小的值為最適的K值,鄰接樹(Neigh?bor-joining tree,NJtree)用TreeBeST[21]構建,為探討3種石斛的遺傳結構關系用Genome-wide complex trait analysis(GCTA)[22]軟件進行主成分分析。用GenAlEx軟件計算3種石斛的遺傳距離。
2 結果與分析
2.1 數據過濾和比對到參考基因組
霍山石斛、細莖石斛和鐵皮石斛平均每個樣本產生的clean reads是霍山石斛>細莖石斛>鐵皮石斛,分別為1 535 753、1 380 009和1 208 693個。平均每個樣本產生的clean bases也是霍山石斛>細莖石斛>鐵皮石斛的,分別為436 387 047 bp、391 592 914 bp和344 268 426 bp。3種石斛比對到黃石斛參考基因組的reads數占比最高的是鐵皮石斛,為99.06%,霍山石斛和細莖石斛相差不明顯,分別為97.00%和96.98%;測序長度符合一定閾值且雙端的reads都比對上參考基因組的reads數占比最高的也是鐵皮石斛,為91.35%,霍山石斛、細莖石斛之間差異不明顯,分別為79.79%和80.08%(表1)。
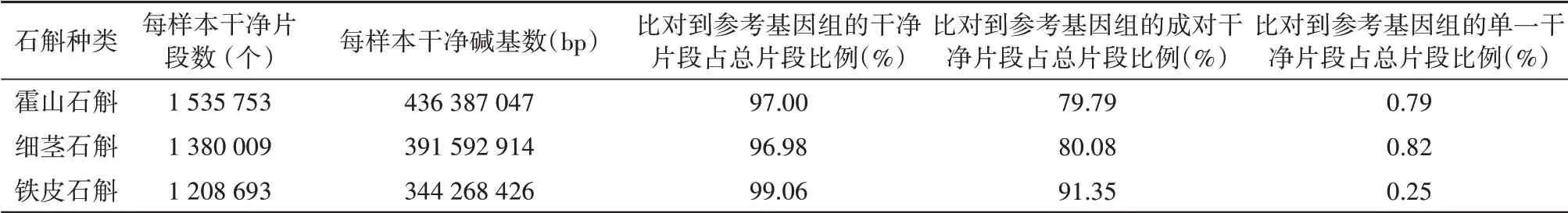
表1 每樣本產生干凈片段的數目和平均比對到參考基因組的比例Table 1 Summary of clean reads per specimen detected by GBSand percentage of alignment to reference genome sequence
2.2 SNP的變異檢測數目
霍山石斛、細莖石斛和鐵皮石斛的平均每個樣本產生的SNP數目霍山石斛>細莖石斛>鐵皮石斛,分別為1 509 648、1 364 605和903 183個。其轉換和顛換的SNP數目也是霍山石斛>細莖石斛>鐵皮石斛,分別為899 064和6 108 125,810 614和551 968,552 704和347 449個,鐵皮石斛的轉換和顛換的比例是1.59,其余的都是1.47。霍山石斛、細莖石斛和鐵皮石斛平均每個樣本產生的雜合和純合SNP數分別為204 940和1 304 709,168 830和1 195 774,216 436和686 747個,雜合SNP數占總SNP比最高的是鐵皮石斛為0.24,霍山石斛和細莖石斛差異不明顯分別為0.14和0.12(表2)。
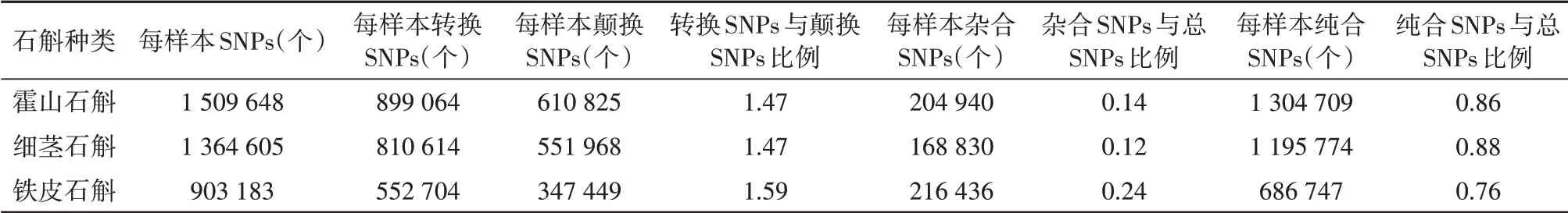
表2 GBS檢測獲得的每個樣本SNP數目Table 2 Summary of average SNPs per specimen detected by GBS
2.3 群體結構和遺傳距離分析
Clean reads過濾后得到用于群體結構分析的總SNP數為187 143個,其中霍山石斛、細莖石斛和鐵皮石斛分別有120 734、97 441和73 787個。根據Tree BeST軟件計算遺傳距離陣,然后用鄰接法構建系統NJ進化樹(圖1),從圖1中看出明顯分為2大支,鐵皮石斛聚為一支,細莖石斛和霍山石斛聚為另一支,細莖石斛和霍山石斛又被嚴格分為2支。根據Admixture軟件分析的遺傳結構(圖2A和圖2B)顯示:最佳的K值是2,鐵皮石斛聚為一支,霍山石斛和細莖石斛聚在一起,暗示其遺傳關系更接近;當K=3時,分為3支,3種石斛被嚴格分開與NJ樹相似。主成分分析(Principal component anal?ysis,PCoA)與Structure和NJ樹分析類似(見圖3),也是3種石斛被嚴格分開,總的遺傳變異是38.21%,而PC1和PC2分別為26.99%和11.22%。遺傳距離分析發現:霍山石斛與鐵皮石斛間的遺傳距離最大為0.3052,其次是細莖石斛與鐵皮石斛間的為0.2792,最低為霍山石斛與細莖石斛間的為0.2141。

圖1 基于20個石斛植物樣本的187 143 SNPs-GBS的遺傳距離矩陣構建鄰接樹Fig.1 NJ tree based on pairwise distance matrix representing the grouping of the 20 Dendrobium specimens obtained from 187 143 SNPs-GBS
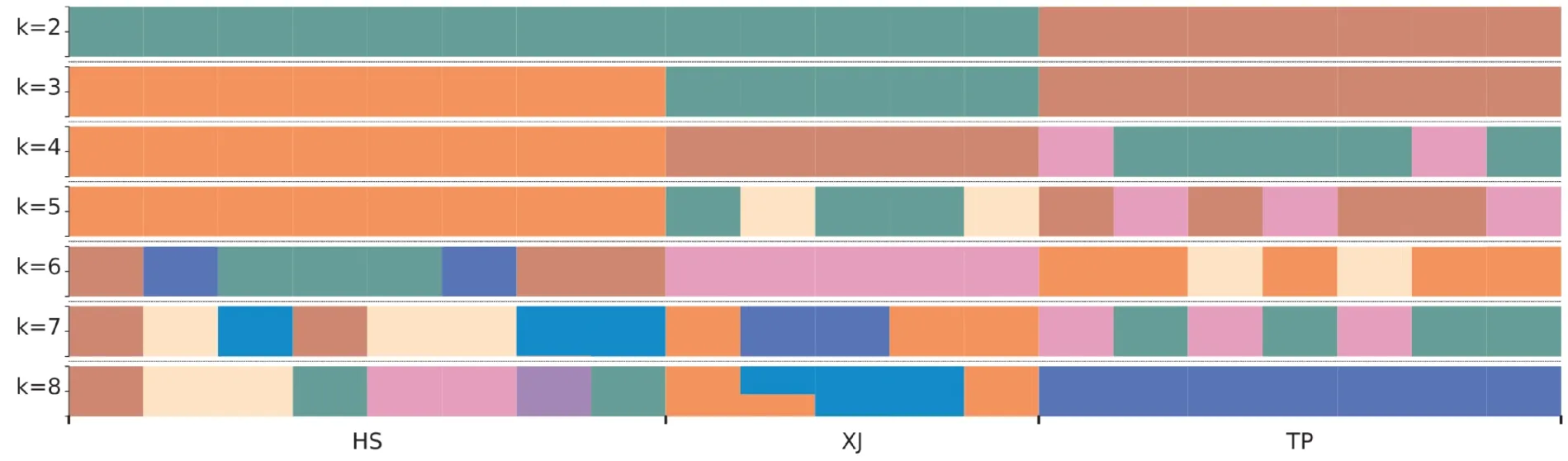
圖2A 根據Admixture軟件構建的20個石斛樣本的遺傳結構圖Fig.2A Genetic structure of 20 Dendrobium specimens for K=2-8 based on the Admixture software
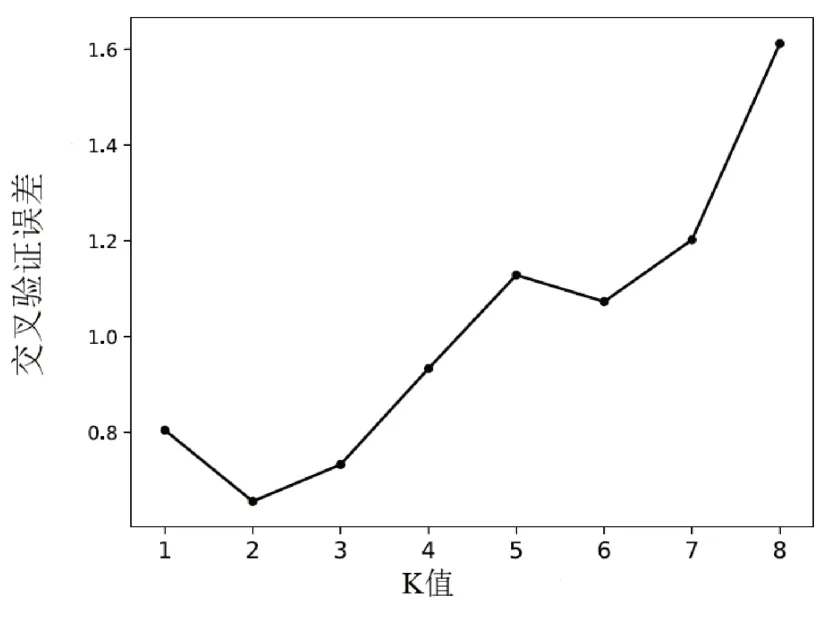
圖2B 交叉驗證誤差和K值Fig.2B Cross-validation error and K value
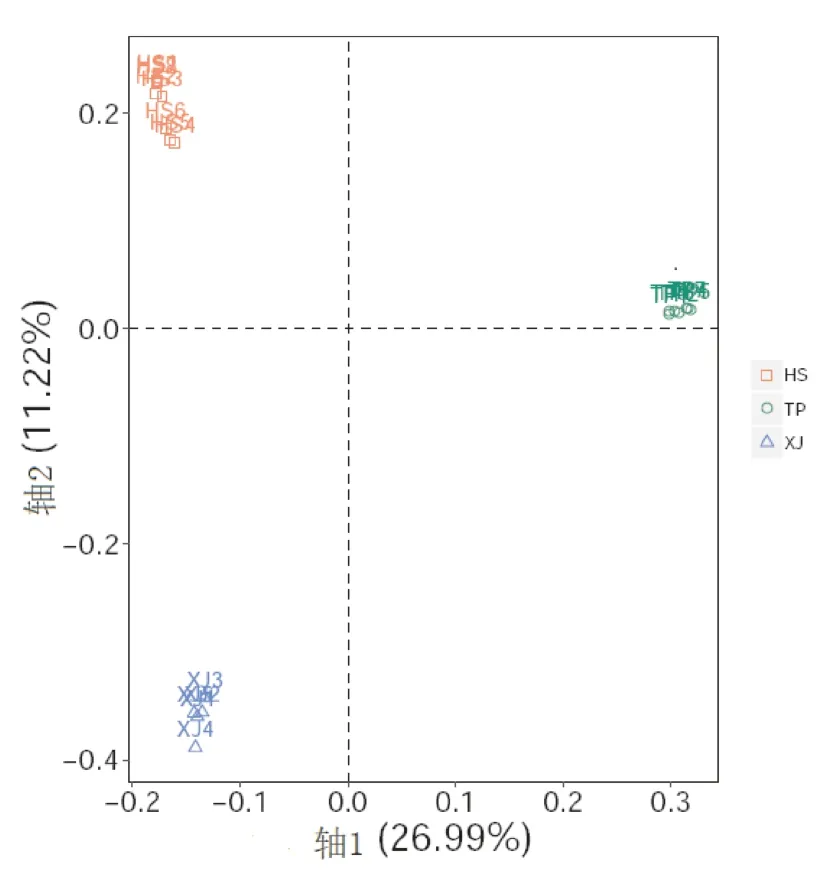
圖3 根據樣本間匹配相似性構建的主成分圖(PCoA)Fig.3 Principal coordinates analysis(PCoA)of pairwise sim?ple matching dissimilarities between specimens
3 討論
霍山石斛、鐵皮石斛與細莖石斛都是我國傳統的名貴藥用植物,而關于三種石斛的遺傳關系一直是石斛研究者感興趣的問題。如鄧輝等[23]、劉明珍等[24]分別用重復序列聚合酶鏈式反應(Inter-simple sequence repeat,ISSR)技術證明了霍山石斛與相似種鐵皮石斛、細莖石斛等存在較大的、穩定的差異,認為霍山石斛不屬于鐵皮石斛,應為一獨立的種;樊洪泓等[25]根據隨機擴增多態性DNA(Random am?plified polymorphic DNA,RAPD)分子標記研究認為霍山石斛與細莖石斛親緣關系更近,而與鐵皮石斛的親緣關系相對較遠;而其根據相關序列擴增多態性(Sequence-related amplified polymorphism,SRAP)標記研究認為鐵皮石斛與霍山石斛也具有較近的親緣關系。陸安靜等[26]通過內轉錄間隔序列(Inter?nal transcribed spacer,ITS)分析比較認為細莖石斛與鐵皮石斛的親緣關系較近;李國良等[27]通過ITS、Nad intron2和psbA-trnH系列分析發現,霍山石斛與鐵皮石斛在這些序列上具有明顯的堿基差異,而與細莖石斛差異不明顯,霍山石斛和鐵皮石斛之間的遺傳距離大于其和細莖石斛之間的遺傳距離。這些研究結果與該文研究結果類似,即這三種石斛的遺傳親緣關系都比較近,而霍山石斛與細莖石斛的遺傳關系比霍山石斛與鐵皮石斛的遺傳關系更近。
探討不同種石斛的遺傳關系,一般野生樣本更具有說服力,人工栽培因人為的選擇可能會造成遺傳多樣性的丟失或者遺傳背景的復雜,如Hou[28]利用三核苷酸微衛星(Trinucleotide microsatellite)標記分析發現異位收藏保護的鐵皮石斛和其野生居群相比基因多樣性并沒有顯著的差異,但Structure分析揭示了其在遺傳組成上具有明顯的差異;張君毅等[29]采用目標區域擴增多態性(Target region ampli?fication polymorphism,TRAP)和特異序列擴增區域(Sequence-characterized amplified region,SCAR)標記研究得出鐵皮石斛野生類群遺傳多樣性水平最高,而仿野生類群和栽培類群基本一致;而魏丹紅和徐紅[30]用ISSR標記研究認為金釵石斛栽培居群的遺傳多樣性和野生相比基本上沒有喪失;吳永輝等[31]利用保守DNA衍生多態性(Conserved DNA-de?rived polymorphism,CDDP)分子標記研究表明鐵皮石斛人工栽培種具有豐富的遺傳多樣性;該研究因資金的有限和石斛野生資源的稀缺未能完成野生和栽培石斛遺傳關系的比較研究,而所選的細莖石斛栽培和野生樣本量太少,無法準確判斷兩者差異,希望未來的研究能比較野生樣本和栽培樣本的遺傳差異。
總之,根據GBS-SNP標記分析發現霍山石斛與鐵皮石斛遺傳關系較遠,而其和細莖石斛遺傳關系較近;遺傳結構、NJ樹及主成分分析結果類似,3種石斛被明顯分開,證明GBS-SNP分子標記可用于該3種石斛的鑒定研究。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
