綿羊OSBPL11、PAEP、ALDH1A2 基因多態性與胸椎數關聯分析
2021-11-09 14:18:10郭思武鐘英杰劉秋月劉玉芳儲明星
中國畜牧雜志 2021年9期
郭思武,鐘英杰,劉秋月,陶 林,劉玉芳,儲明星*
(1.中國農業科學院北京畜牧獸醫研究所,農業農村部動物遺傳育種與繁殖重點實驗室,北京 100193;2.河北工程大學生命科學與食品工程學院,河北邯鄲 056001)
哺乳動物中,亞種或家畜的品種之間經常出現脊椎數的差異,即使在同種內個體間也存在差異。胸椎數的變異在不同的物種中有著千差萬別,如在人類中發生脊椎數目的變異會造成嚴重的遺傳性疾病,影響正常生活[1];在家畜中胸椎數的增加卻是一個非常有利的經濟性狀,家畜胸椎和腰椎數目增加,胴體長度也會隨之增加,這就給養殖企業帶來更高的經濟效益[2-3]。在豬[4-5]、牛[6]、羊[7-8]等物種中均已有多個基因調控脊椎數變異的相關報道。本研究選取的小尾寒羊(STH)是我國肉裘兼用型綿羊品種,具有耐粗飼、發育快、繁殖性能強、適應性強等特點[9],而蘇尼特羊(SNT)屬蒙古綿羊系統中的一類,經過長期的自然選擇和人工選擇而形成了具有耐寒、抗旱、生長發育快、生命力強的特性,是最能適應荒漠半荒漠草原的肉用地方良種之一[10]。這2 種羊均存在高比例的多脊椎數變異,含有14 個胸椎(T14,正常綿羊為T13)的個體比例高達20%~30%,同時其肉質細嫩呈大理石紋狀,富含人體必需的各種氨基酸、維生素、礦物質等營養物質,具有繁殖選育的潛力[11]。研究綿羊胸椎數相關基因已持續數年,通過分子生物學技術選育多脊椎數性狀的綿羊品種已成為當前階段的重要研究內容。因此,本研究基于先前實驗的基因組重測序結果,對得到的21 個候選基因中的OSBPL11、PAEP、ALDH1A2基因進行胸椎數關聯分析研究,以期獲得新的多胸椎數相關分子標記。
OSBPL11、PAEP、ALDH1A2基因基于綿羊基因組重測序被篩選出來,但目前研究進展表明這3 個基因的功能在胸椎數變異中并無明顯的關聯性。其中,OSBPL11是氧甾醇結合蛋白(Oxysterol-Binding Protein,OSBP)家族的成員之一,OSBP 在20 世紀80 年代作為幾種氧甾醇的細胞質親和力受體被分離出來[12-14]。與OSBP 具有序列同源性的蛋白質家族存在于整個真核生物界,在哺乳動物中,OSBP 家族由12 個成員組成[15-17]。孕激素相關子宮內膜蛋白(Progestagen Associated Endo metrial Protein,PAEP)長約5 050 bp,包括7 個外顯子和6 個內含子,該基因由180 個氨基酸構成,是一種分子大小為28 kDa 的糖蛋白,屬于核脂鈣蛋白家族[18]。PAEP是子宮內膜分泌的主要蛋白質之一,其中在雄性精囊中表達最高,PAEP還在生殖組織的其他上皮細胞中表達。醛脫氫酶1A2(ALDH1A2)是醛脫氫酶(ALDH1)家族中的一員,ALDH1 由ALDH1A1、ALDH1A2 和ALDH1A3 3 個亞科成員組成[19]。其中ALDH1A1參與視網膜氧化、乙醛代謝和環磷酰胺解毒,而ALDH1A2和ALDH1A3參與視網膜氧化為維甲酸(RA)[20]。因此,本研究利用分子遺傳學手段檢測了蘇尼特羊和小尾寒羊OSBPL11、PAEP、ALDH1A2基因的多態位點,并將這些位點與胸椎數性狀進行關聯分析,以揭示這3 個基因的遺傳變異與多脊椎數性狀之間的關系,初步推測OSBPL11、PAEP、ALDH1A2基因影響蘇尼特羊和小尾寒羊胸椎數的具體分子機理,為提高綿羊胸椎數、改良綿羊生長發育提供理論基礎。
1 材料與方法
1.1 樣品采集 實驗采集了188 只來自內蒙古巴彥淖爾烏拉特中旗的蘇尼特羊的組織樣品(樣品組織為背最長肌)和195 只來自內蒙古巴彥淖爾及山東鄆城的小尾寒羊的組織樣品,采樣綿羊健康狀態良好。綿羊屠宰后,快速用2 mL 凍存管采集新鮮肌肉組織,并用液氮冷凍保存帶回實驗室轉移到-80℃冰箱保存備用。記錄綿羊胴體長度部分數據,其中蘇尼特羊胸椎數為13 個及14個的個體分別為122 只和66 只,小尾寒羊胸椎數為13個和14 個的個體分別為137 只和58 只。
1.2 DNA 提取 使用DNA 提取試劑盒(天根生物科技有限公司,北京)進行組織DNA 提取,對提取后的組織樣品用NanoDrop 2000 和1.5%瓊脂糖凝膠電泳進行濃度和質量的評估,合格的DNA 樣品用于后續實驗。
1.3 引物設計 根據小尾寒羊和蘇尼特羊OSBPL11、PAEP、ALDH1A2基因的SNP 位點序列信息通過Assay design 3.1 軟件進行引物設計,具體信息見表1。引物設計及合成由康普森生物技術有限公司完成。
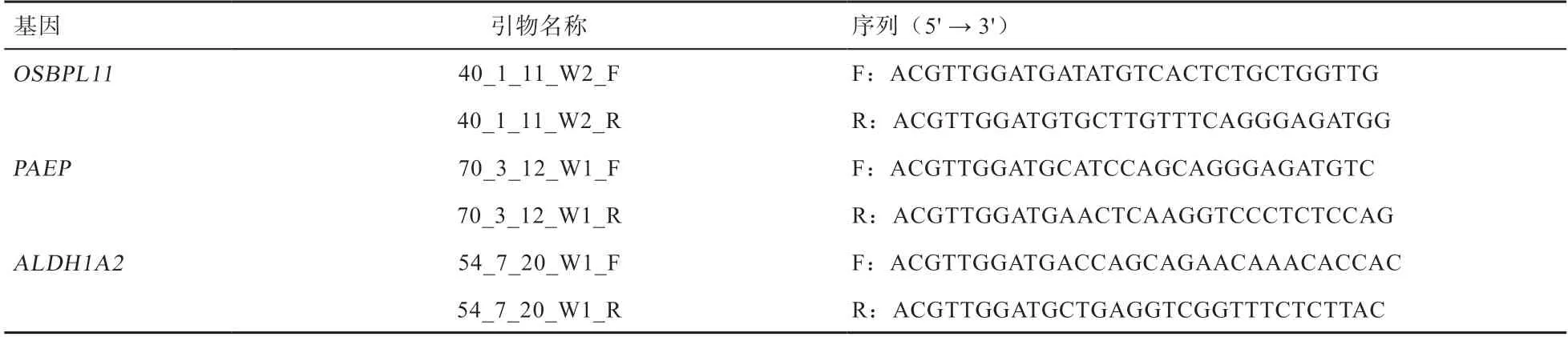
表1 小尾寒羊與蘇尼特羊OSBPL11、PAEP 和ALDH1A2 基因的引物序列
1.4 基因分型 采用Sequenom MassARRAY?SNP[21]分型技術對OSBPL11基因的g.188064535A>G、PAEP基因的g.3541777A>G 和ALDH1A2基因的g.49249275G>A進行分型檢測,分型樣品為DNA,每個樣品需要量為20 μL,DNA 濃度為40~80 ng/μL。分型的具體過程見Zhang 等[22]所示方法。
1.5 數據統計分析 利用Microsoft Excel 2019 軟件計算各位點的基因頻率、基因型頻率、有效等位基因數、多態信息含量(Polymorphism Information Content,PIC)和雜合度(Heterozygosity,He),并進行哈代溫伯格平衡檢驗。計算群體遺傳學參數的方法詳見Zhang 等[22]。利用SPSS 19.0 采取一般線性模型單變量分析與單因素方差分析LSD 對OSBPL11、PAEP、ALDH1A2不同基因型的小尾寒羊和蘇尼特羊進行胸椎數關聯分析。模型為yijn=μ+Pi+Gj+IPG+eijn。式中:yijn代表表型值(胸椎數);μ 代表群體均值;Pi代表品種效應(i=1,2);Gj代表j 種基因型的影響(j=1,2,3);IPG表示品種與基因型的互作效應;eijn代表隨機誤差。胸椎數用平均值±標準誤表示。
2 結果與分析
2.1OSBPL11、PAEP、ALDH1A2基因多態性分析 測序結果顯示,383 只綿羊群體基因型及分型分別是:OSBPL11基因g.188064535A>G 位 點AA 型350 只、AG型30 只、GG(不計入統計分析)型1 只;PAEP基因g.3541777A>G 位點AA 型216 只、AG 型143 只、GG 型21 只;ALDH1A2基因g.49249275G>A 位點GG 型362 只、AG 型18 只(圖1~3)。
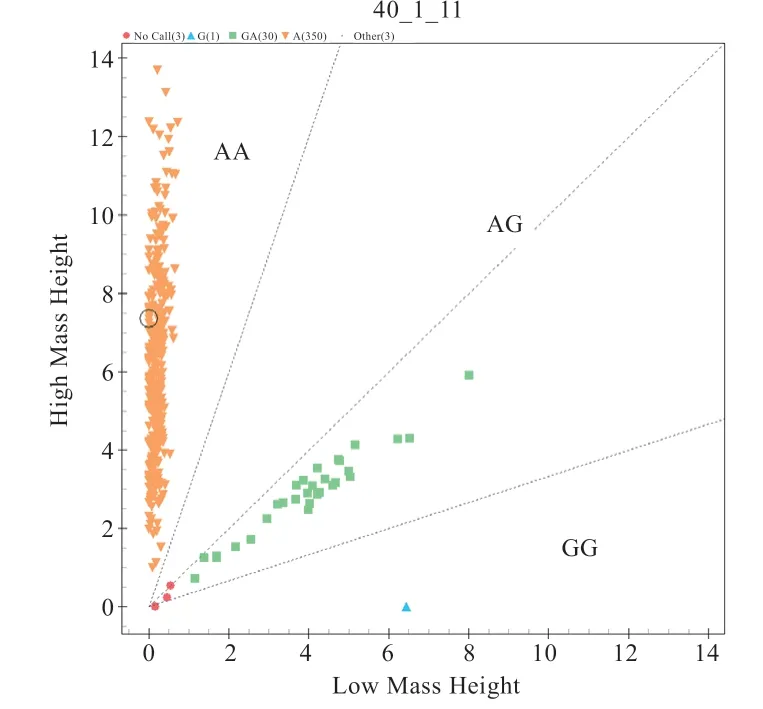
圖1 OSBPL11 基因g.188064535A>G 位點分型結果
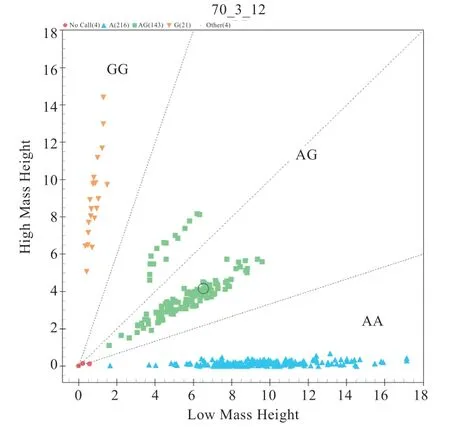
圖2 PAEP 基因g.3541777A>G 位點分型結果
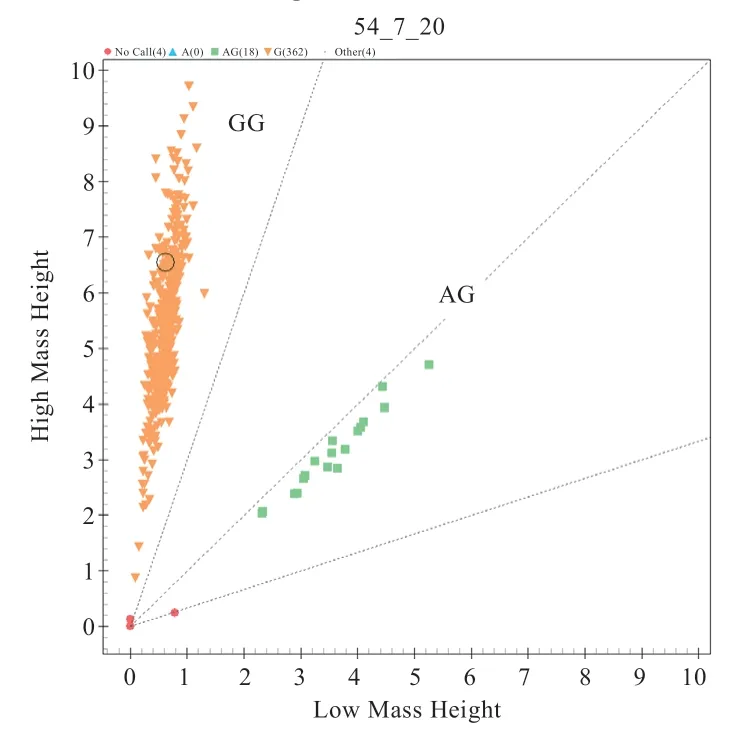
圖3 ALDH1A2 基因g.49249275G>A 位點分型結果
PIC:OSBPL11基因g.188064535A>G位點中小尾寒羊和蘇尼特羊PIC 分別是0.06 和0.09,均屬于低度 多 態(PIC<0.25);PAEP基 因g.3541777A>G 位點中小尾寒羊和蘇尼特羊PIC 分別是0.32 和0.27,均屬于中度多態(0.25<PIC<0.50);ALDH1A2基因g.49249275G>A 位點中小尾寒羊和蘇尼特羊PIC 分別是0.05 和0.04,均屬于低度多態(PIC<0.25)(表2)。在卡方檢驗值方面,OSBPL11基因g.188064535A>G位點小尾寒羊與蘇尼特羊的卡方檢驗值分別為0.631 和0.439;PAEP基因g.3541777A>G 位點小尾寒羊與蘇尼特羊的卡方檢驗值分別為0.427 和0.930;ALDH1A2基因g.49249275G>A 位點小尾寒羊與蘇尼特羊的卡方檢驗值分別為0.685 和0.793。結果表明,小尾寒羊和蘇尼特羊中3 個SNPs 位點均處于哈代溫伯格平衡狀態(P>0.05)。

表2 3 個位點在小尾寒羊和蘇尼特羊中的群體遺傳學分析
2.2OSBPL11、PAEP、ALDH1A2基因3個位點與胸椎數的關聯分析 由表3 可知,OSBPL11、PAEP、ALDH1A2基因的3 個突變位點均與胸椎數不顯著相關。對188 只蘇尼特羊和195 只小尾寒羊2 個種群單獨進行胸椎數關聯分析,結果發現,ALDH1A2基因g.49249275G>A 位點在蘇尼特羊中野生型個體(GG)的胸椎數顯著低于突變雜合個體(AG),其他位點與胸椎數均不顯著相關;而在小尾寒羊中OSBPL11、PAEP、ALDH1A2基因的3 個突變位點與胸椎數均不顯著相關,表明ALDH1A2基因g.49249275G>A 位點與蘇尼特羊的胸椎數變化有關(表4)。
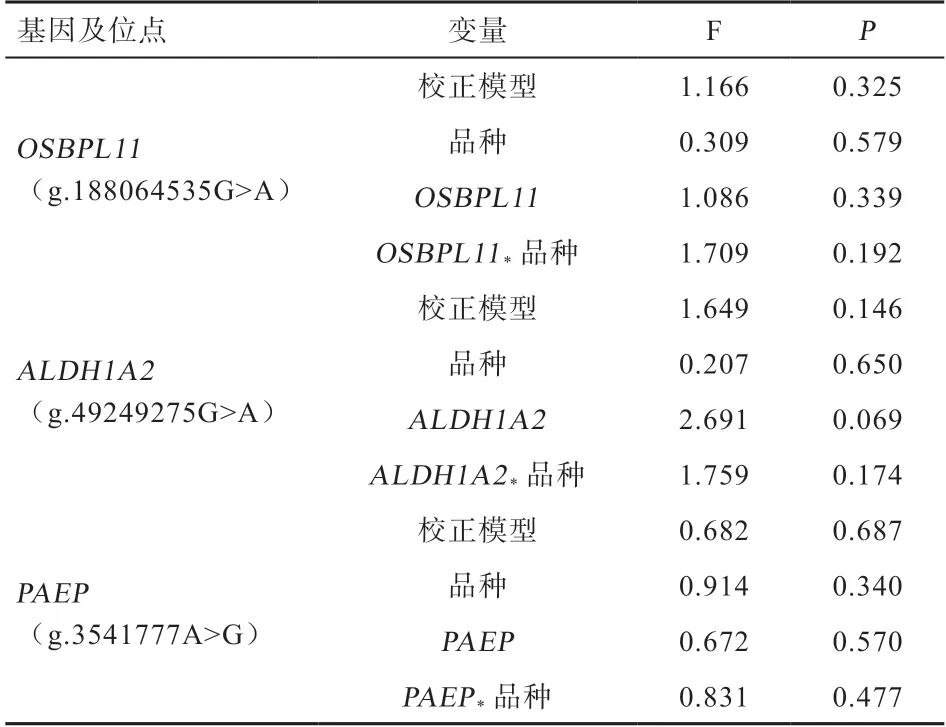
表3 OSBPL11、PAEP 及ALDH1A2 基因位點與綿羊胸椎數關聯分析結果

表4 3 個位點各基因型與小尾寒羊和蘇尼特羊胸椎數最小二乘均值及標準誤
2.3OSBPL11、PAEP、ALDH1A2基因3 個位點與小尾寒羊胴體長的關聯分析 由表5 可知,這3 個突變位點各基因型與小尾寒羊胴體長均不顯著相關。

表5 各位點基因型與小尾寒羊胴體長的關聯分析(平均值±標準誤)
3 討論
哺乳動物脊椎是由胚胎發育到原腸胚時中胚層分化而來,而控制胸椎生長的初級神經胚的發育變化主要受到骨形成蛋白(BMP)的濃度控制。其中發育的每個階段都是極為復雜的工程,任何地方出錯都有可能導致胸椎的異常分化[23-24]。家畜多脊椎性狀是指家畜胸椎數和腰椎數比正常個體增多的現象。早在20 世紀中期,人們就發現家畜的胸腰椎數目存在變異。脊椎數的增多可以間接地提高家畜的體長、體重、產肉量和肋骨數等性狀,從而提高其經濟效益。所以家畜脊椎數變異的遺傳改良也在近年來得到了深入的研究。
豬多脊椎性狀候選基因的挖掘始于QTL 定位。由于豬的繁殖速度較其他家畜快,使得在利用家系進行多脊椎性狀相關QTL 定位研究中較其他單胎家畜具有明顯的優勢,這也是豬繁殖研究中獲得成果較其他家畜多的原因之一。早在2000 年,Wada 等[25]首次將脊椎數相關QTL 位點定位在豬1 號染色體和2 號染色體上。其中1 號染色體上脊椎數相關的QTL 與Rohrer 等定位在1 號染色體與胴體長度相關的QTL 在位置上很接近[26],提示脊椎數與胴體長可能存在相關性。在此基礎上,該研究團隊后期研究又發現在豬1 號染色體和7 號染色體各存在1 個與脊椎數相關的QTL 位點[27],該結果后來也被在其他不同豬品種上的研究所證實[28-29]。與豬相比,羊脊椎數變化范圍要小很多。張立嶺等[3]研究發現,蒙古羊每增加1 枚胸椎或腰椎,脊柱就加長約2.40 cm或3.50 cm。無論多1 枚胸椎還是腰椎,綿羊的活重、胴體重、凈肉重和眼肌面積均明顯增加,其中凈肉平均增加4 kg 以上。該研究結果與豬的研究結果基本一致。最早在大白豬群體中發現控制脊椎數性狀的VRTN基因首次被證實為影響脊椎數性狀的主效基因[30],隨后又發現了更多影響胸椎數性狀的基因,如NR6A1[7]、Hoxc[31]和TGFβ3[32]等,而在蘇尼特羊中已被證實VRTN對胸椎數的影響仍然存在[33]。
查閱OSBPL11基因相關文獻發現,該基因的表達在各物種間可能存在差異,人OSBPL11基因的功能與脂肪形成有關,綿羊中卻在肺臟和淋巴組織中高度表達。目前OSBPL11所在家族中的其他成員OSBP 相關(ORP)蛋白或OSBP 樣(OSBPL)蛋白更多的研究主要聚焦在哺乳動物和酵母系統中,主要參與控制細胞脂質代謝、囊泡運輸和細胞信號傳導[34-35],在脊椎數變異等骨相關的研究鮮少。查閱PAEP基因相關文章發現,還沒有發現PAEP在綿羊胸椎數變異研究的報道,已知的PAEP 蛋白亞型有4 種,分別為glycodelin A(來自羊水、內膜和母體血清)、glycodelin S(來自雄性精漿和精液囊泡)、glycodelin F(來自卵泡液和輸卵管)和glycodelin C(來自卵丘),這些亞型的蛋白骨架完全相同,區別僅在糖基結構部分,表現出不同甚至相反的功能,提示糖基化對蛋白功能具有重要的調節作用[36]。且在NCBI 中發現PAEP主要在綿羊乳腺中高度表達,其原因是由于PAEP的表達受孕酮/孕激素、松弛素和組蛋白去乙酰化酶抑制劑的調控[37]。
ALDH1A2是醛脫氫酶1(ALDH1)家族的一員,而ALDH1 是通過氧化全跨視網膜(all-trans-retinal)和9-順視網膜(9-cis-retinal)產生維甲酸(RA)的主要酶,主要參與細胞分化、細胞周期阻滯最終凋亡等生物學功能[38-40]。ALDH1A2的功能與胸椎數性狀沒有直接關系,但是本研究結果表明,ALDH1A2基因在存在品種效應的條件下雖與胸椎數呈不顯著關系,但在單獨群體中ALDH1A2基因的突變位點可以顯著提高蘇尼特羊的胸椎數,其機理有待進一步研究。
4 結論
在小尾寒羊中OSBPL11基因g.188064535A>G、PAEP基因g.3541777A>G 和ALDH1A2基 因g.49249275G>A位點與胸椎數和胴體長無顯著關聯。OSBPL11基因g.188064535A>G 和PAEP基 因g.3541777A>G 位 點與蘇尼特羊胸椎數性狀無顯著關聯;ALDH1A2基因g.49249275G>A 位點突變與蘇尼特羊胸椎數呈顯著相關。ALDH1A2基因可作為潛在的多胸椎數的分子標記。
