游離脂肪酸對畜禽腸道內分泌細胞作用的研究進展
2021-11-09 14:18:06陳文靜楊曉靜
中國畜牧雜志 2021年9期
陳文靜,楊曉靜
(南京農業大學動物醫學院,江蘇南京 210095)
腸道是機體最重要的消化器官,也是與外界物質接觸最為密切的部位。營養物質包括脂肪酸在胃腸道的消化與吸收影響著胃腸運動和分泌,進而影響機體代謝等生理過程。這些重要的生理功能都離不開腸道上皮層不同類型的細胞,如吸收性腸上皮細胞、杯狀細胞、內分泌細胞、潘氏細胞和M 細胞等。其中,腸內分泌細胞(Enteroendocrine Cells,EECs)所分泌的多種激素參與調節腸道自身的運動和分泌功能、食欲和攝食行為、胃腸化學感應、營養物質代謝等過程[1]。因此,脂肪酸和腸道內分泌之間相互作用,在調控機體營養吸收和能量代謝等方面具有重要作用。本文綜述了脂肪酸對畜禽腸道EECs 的作用,為其調控畜禽腸道健康、生長發育以及相關疾病的防治提供參考。
1 脂肪酸
脂肪酸是一類由碳、氫、氧3 種元素組成的化合物。不同鏈長的脂肪酸有不同的代謝途徑,脂肪酸鏈長是脂肪消化和吸收的重要決定因素。根據碳鏈長度可將其分為短鏈脂肪酸(碳鏈上碳原子小于6,Short-Chain Fatty Acids,SCFAs)、中鏈脂肪酸(碳鏈上碳原子為6~12,Medium-Chain Fatty Acids,MCFAs)、長鏈脂肪酸(碳鏈上碳原子數大于12,Long-Chain Fatty Acids,LCFAs)。
1.1 SCFAs 及其在腸道的功能 SCFAs 又稱為揮發性脂肪酸,主要包括乙酸、丙酸、丁酸、異丁酸和異戊酸等。大多數SCFAs 是由腸道微生物(如乳酸桿菌、雙歧桿菌等)發酵分解不易消化的碳水化合物(包括抗性淀粉和可溶性低聚糖和多糖)而產生的,其他SCFAs 則是由飲食攝入和蛋白質等代謝產生[2]。腸道SCFAs 中乙酸、丙酸和丁酸含量較高,它們的比例約為3:1:1,占總SCFAs 的90%~95%,其余僅占很小一部分[3]。發酵產生的SCFAs 在體內參與不同組織的代謝,它們可被腸上皮細胞吸收并在線粒體內被氧化,為機體提供5%~10%的能量[4]。SCFAs 除了是一種瞬時能源外,還被認為是局部和遠端生理功能中的關鍵信號分子,如跨上皮陰離子分泌、腸道運動和全身能量代謝等[5]。
SCFAs 主要通過抑制組蛋白去乙酰基酶(HDACs)和激活SCFAs 受體2 種方式對細胞產生影響進而促進腸道穩態。SCFAs 作為配體誘導HDACs 抑制作用,抑制NF-κB 途徑,下調促炎細胞因子的表達,進而調控免疫反應[6]。目前已經發現的SCFAs 受體有FFAR2(也被稱為GPR43)、FFAR3(GPR41)以及GPR109A。這些受體分布廣泛,不僅大量分布于腸上皮細胞,在腸內分泌細胞、免疫細胞、肝細胞、脂肪細胞以及神經細胞中也都有分布[7]。
在SCFAs 中,丁酸在畜禽上的應用最為廣泛,但是丁酸易揮發、有濃烈的酸臭味并且較難傳輸到腸道后段,因此目前生產上常用丁酸鹽和丁酸甘油酯。Sun 等[8]研究表明,在生長育肥豬日糧中添加丁酸鈉可降低耗料增重比,增加胴體重量并增加豬盲腸中擬桿菌的相對豐度,改善腸道健康。在經產母豬日糧中添加丁酸鈉顯著提高了仔豬出生窩重、新生仔豬體重和斷奶體重[9]。而斷奶仔豬日糧中添加丁酸鈉顯著增加了腸道絨毛高度和絨毛高度/隱窩深度比,抑制了肥大細胞活化和炎性介質產生[10],增強腸黏膜緊密連接蛋白的表達[11],增強空腸和結腸閉鎖蛋白的表達,改善細菌群落[12],降低了斷奶后腹瀉發病率。在肉雞日糧中添加丁酸鹽可以促進腸道發育,誘導抗炎作用以及調節肉雞腸道微生物群落[13-15],并有效控制了腸道中沙門氏菌的繁殖。劉麗華等[16]研究表明,在14 日齡肉仔雞飼糧中添加不同丁酸鈉可顯著提高其采食量和體重,顯著降低耗料增重比。
1.2 中、長鏈脂肪酸及其在腸道的功能 MCFAs 包括己酸(C6)、辛酸(C8)、葵酸(C10)和月桂酸(C12),被甘油脂化后形成中鏈甘油三酯(Medium Chain Triglycerides,MCT)。MCFAs 主要存在于椰子油和棕櫚油等植物油中,在反芻動物的乳脂和椰子油中含量較高,但在黃油中MCFAs 只有不到6%,玉米油中不含任何MCFAs[17]。
LCFAs 又稱為高級脂肪酸。在碳原子個數為14~24的LCFAs 中,LCFAs 的碳原子個數多為偶數,因為自然條件下脂肪酸的生物合成總是以2 個碳原子為擴增單位,如豆蔻酸(C14)、軟脂酸(C16)、硬脂酸(C18)以及花生酸(C20)等飽和脂肪酸。LCFAs 可根據其碳鏈的飽和程度分為飽和脂肪酸、單不飽和脂肪酸以及多不飽和脂肪酸。根據不飽和脂肪酸雙鍵所連碳原子的位置不同,LCFAs 又可分為n-3、n-6、n-9 等,包括亞麻酸(LA)、油酸、亞油酸(LOA)、花生四烯酸(AA)和二十二碳六烯酸(DHA)等。LCFAs 在機體的生長發育、保護腸黏膜屏障以及刺激腸道內分泌細胞分泌激素等方面起著重要作用。
MCFAs 和LCFAs 主要通過激活受體發揮作用,從而對細胞產生影響調節腸道穩態。不同研究都表明,FFAR1(GPR40)和FFAR4(GPR120)可同時被MCFAs 和LCFAs 激活[18],但是GPR84 只能被MCFAs激活[19]。FFAR1 和FFAR4 分布廣泛,LCFAs 直接和間接誘導了胰島素分泌證明FFAR1 和FFAR4 在腸道中也大量分布[20-21]。GPR84 在腦、結腸、肝臟和脾臟等都有分布[18]。
仔豬對MCFAs 的消化吸收率很好,除了供能作用,還可提高仔豬斷奶后的消化率和飼料效率[22],緩解腸道損傷。研究表明,斷奶前飼喂MCFAs 提高了仔豬的生長性能,促進了腸道發育[23],降低了弱仔豬的死亡風險[24]。日糧中添加魚油(富含ω-3 多不飽和脂肪酸)能夠抑制LPS 對斷奶仔豬的刺激,緩解LPS 對仔豬的生長抑制[25]。在經產母豬日糧中添加魚油和大豆油顯著提高了哺乳仔豬的生長性能[26]。在肉雞日糧中添加MCFAs 顯著降低了腹部脂肪等,防止脂肪過度堆積,提高飼料利用率和胴體品質[27]。早期研究表明,飼糧中添加魚油可以有效減少柔嫩艾美耳球蟲對雛雞增重的不利影響以及腸道病變的程度[28]。在肉仔雞和蛋雞日糧中添加多不飽和脂肪酸能一定程度改善肉品質[29],產出富含多不飽和脂肪酸的雞蛋[30]。
2 EECs
EECs 是由腸道干細胞分化而來,是腸上皮細胞中重要的一員,雖然在整個胃腸腔上皮細胞中EECs 僅占不到1%,但是其數量超過了機體其他器官所有內分泌細胞的總和,并且分泌超過20 種激素,因此EECs 被認為是機體內最大、最復雜的內分泌器官[31]。EECs 散布在腸管黏膜的上皮組織中,大多為“開放型”,其頂端表面覆蓋著一層微絨毛,與管腔成分直接接觸,感受管腔食物刺激和腸道pH 變化等;還有很大比例的EECs 為“封閉型”,包括腸嗜鉻細胞(EC 細胞)、腸嗜鉻樣細胞(ECL 細胞)、X/A 細胞和D 細胞,它們不直接接觸管腔,通過旁分泌感受內容物等變化來調節其內分泌狀態[32]。
2.1 EECs 的分類 EECs 通常根據其產生的主要激素來進行分類(表1)[33]。在十二指腸中,EECs 中的K 細胞分泌葡萄糖依賴性促胰島素多肽(Glucose-Dependent Insulinotropic Polypeptide,GIP),具有抑制胃酸分泌、促進胰島素釋放的作用[34]。I 細胞分泌膽囊收縮素(Cholecystokinin,CCK),具有抑制采食、促進胰酶分泌和膽囊收縮的作用[35]。S 細胞分泌腸促胰液素(Secretin,SCT),可以抑制胃酸分泌、促進胰腺分泌[33]。位于小腸遠端和結腸的L 細胞產生胰高血糖素樣肽1 和2(GLP-1 和GLP-2),也產生肽YY(PYY),具有促進胰島素分泌、抑制采食等作用[33]。EC 細胞分布在腸道各處,存儲了胃腸道中90%以上的5-羥色胺(5-Hydroxytryptamine,5-HT),分泌的5-HT可以刺激腸道平滑肌收縮,調節胃腸道運動[36]。
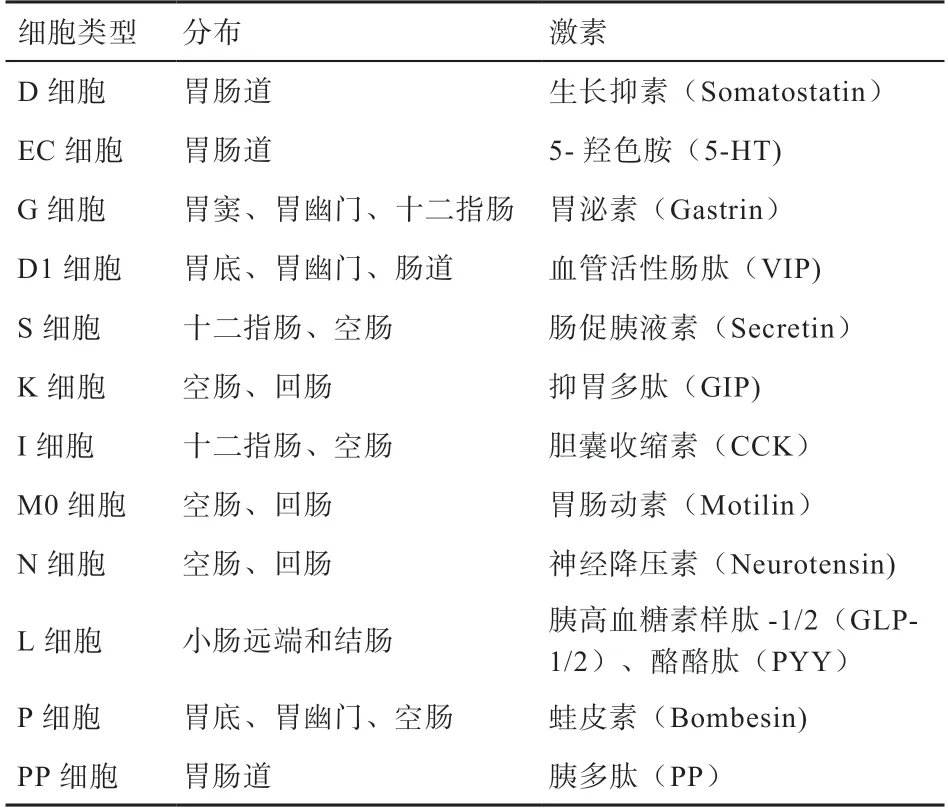
表1 主要腸道內分泌細胞及其分泌的激素[32]
2.2 腸內分泌細胞的分布 EECs 的分布存在物種間差異,比如小鼠的L 細胞主要分布于回腸和結腸,豬的L細胞主要分布于十二指腸[37],而雞的L 細胞則沿著小腸分布[38]。在豬的腸黏膜中,S 細胞和I 細胞主要分布在十二指腸黏膜[39]。同時EECs 的分布還受到外界環境的影響,當飲食中含有高脂飼料時,小鼠腸道的L 細胞和K 細胞的數量均顯著減少,且表達量也顯著下降[40]。
EECs 釋放的激素大多數通過內分泌的方式進入血液循環作用于靶細胞;有一些激素通過旁分泌進入細胞間隙,擴散到鄰近靶細胞發揮作用;還有一部分釋放的激素通過自分泌的方式作用于靶細胞膜上。EECs 釋放的激素與靶細胞上的受體結合發揮相應的生理作用,如調節采食活動和胃酸分泌、調節胰島素分泌、膽囊收縮和胃腸蠕動等。
3 脂肪酸對EECs 作用的影響
游離脂肪酸不僅是必需的營養元素,還參與調節許多細胞功能。游離脂肪酸通過EECs 表面的G 蛋白偶聯受體(GPCRs)對細胞產生影響,進而影響腸道功能。許多研究表明,GPCRs 作為營養物質傳感器,其在整個胃腸道中都有表達[41]。Symonds 等[41]研究表明,短鏈脂肪酸受體FFAR2 在結腸近端高表達,FFAR3 在空腸遠端表達最強。中鏈脂肪酸受體GPR84 和中、長鏈脂肪酸受體FFAR1 在回腸中表達最高,而FFAR4 在結腸近端中表達水平最高[41]。這種分布與其生物學效應有關,中、長鏈脂肪酸來源于飲食,相比之下,腸道中SCFAs 主要由微生物發酵產生,因此,FFAR2 和FFAR3 在腸道前段表達較低,主要存在于腸道后段。
3.1 SCFAs 對EECs 分泌的影響 在SCFAs 中,FFAR2和FFAR3 都可以識別SCFAs,但其優先識別的SCFAs不同,FFAR2 優先識別乙酸和丙酸,而FFAR3 優先感應丙酸、丁酸和其他SCFAs[42]。腸腔內SCFAs 濃度增加可誘導L 細胞和EC 細胞增殖,乙酸和丙酸通過Gq/11 而不是Gi 增加細胞內鈣水平,從而誘導L 細胞分泌GLP-1[5]。相反,在一項在體研究中,丁酸(而不是乙酸和丙酸)增加了野生型小鼠的GLP-1 和PYY水平,而在FFAR3-/-鼠中未觀察到這一現象,并且丁酸和丙酸對K 細胞分泌GIP 的刺激作用也不依賴于FFAR3[43]。Moquet 等[44]研究表明,在幼齡肉仔雞日糧中添加丁酸改變了消化過程和氨基酸的生物利用度,可能與腸道內分泌L 細胞分泌活動的改變有關。
3.2 LCFAs 對EECs 分泌的影響 研究表明,FFAR1 敲除鼠在灌胃LCFAs 后,CCK、GLP-1 和GIP 分泌減少[45]。在腸內分泌細胞K 細胞中,FFAR1 在小腸下部K 細胞中高表達,FFAR4 在小腸上部的K 細胞中高表達,當FFAR4 被敲除后,K 細胞的GIP 分泌量減少,FFAR4 在脂肪酸誘導的GIP 分泌中起著重要作用,有利于在高脂引發的肥胖中誘導胰島素分泌維持正常血糖水平[34]。FFAR4 可以與α-亞麻酸(ALA)和二十二碳六烯酸(DHA)結合,使胞外Ca2+通過L 型通道進入刺激STC-1 細胞株釋放CCK、GLP-1 和PYY[46]。雖然LCFAs 的這些作用與FFAR1 和FFAR4 的激活有著密切關系,但這些效應可能還涉及其他機制。有研究表明,ALA 仍然能刺激FFAR1/4 雙敲除鼠建立的混合結腸上皮細胞分泌GLP-1[47]。ALA 和DHA 也可以刺激L細胞上的FFAR1 產生GLP-2,從而改善DSS 誘導的小鼠結腸炎[48]。
除了FFAR1 和FFAR4 外,CD36 和脂肪酸轉運蛋白(FATP4)與LCFAs 直接結合作用,促進腸腔脂肪酸的吸收[45,49]。橄欖油灌胃后,與野生型小鼠相比,CD36 敲除小鼠CCK 和促胰液素分泌減少;在體外,脂肪酸促進表達CD36 的細胞釋放促胰液素和CCK,對脂肪消化和內環境穩定具有重要意義[49]。
4 EECs 中脂肪酸受體下游信號通路
LCFAs 受體FFAR1 和FFAR4 均與Gq偶聯,通過激活三磷酸肌醇受體和內質網Ca2+的釋放,激活胞吐作用的其他相關信號分子[40]。SCFAs 受體FFAR2 和FFAR3 中,FFAR2 被證實可以通過Gq和Gi/o參與調節cAMP 途徑,而FFAR3 對Gs具有選擇性[50],cAMP 和下游效應蛋白Epac2 和PKA 都可能參與胞吐機制。
LCFAs 與FFAR1 和FFAR4 結合,使得FFAR1 和FFAR4 與Gq/11偶聯,激活下游信號通路PLC-β,生成IP3 和DAG,IP3 調節細胞內鈣信號,Ca2+分泌增加,而DAG 調節PKC,共同調控GLP-1 等激素的分泌。但是,FFAR1 的激活調節信號Ca2+并不是從細胞的鈣庫中釋放的,而是通過激活TRPC3、膜去極化和電壓依賴的Ca2+進入原代L 細胞中的[51]。有研究證實,EC細胞中存在α2-腎上腺素能受體下游的百日咳毒素敏感刺激機制(瞬時受體電位TRPC4 通道的激活)[52]。在STC-1 細胞中,LA 可激活STC-1 細胞的瞬時受體電位通道M5(Trpm5),干擾降低Trpm5的mRNA 表達后,LA 誘導的細胞內鈣和CCK 分泌的增加明顯減弱,FFAR4 通過Trpm5 誘導STC-1 細胞CCK 分泌[53]。在原代L 細胞和GLUtag 細胞中,TRPA1 激動劑使細胞內Ca2+濃度升高,促進GLP-1 的分泌,當把TRPA1敲除或用TRPA1 抑制劑后,這些作用都顯著減弱[54]。FFAR2 優先與2~3 碳的SCFAs 結合,通過Gq-磷脂酰肌醇途徑增加細胞內Ca2+濃度,也可通過G 蛋白亞基Gi/o途徑減少細胞內cAMP,而FFAR3 只能通過Gi/o途徑優先與剩下的SCFAs 結合[55]。
除了FFARs,CD36 在十二指腸和空腸絨毛細胞的刷狀膜緣處表達,脂肪酸通過CD36 增加細胞內鈣和cAMP,進而調節EECs 的分泌[57]。LCFA 與CD36 結合后引起Src-PTKs 的磷酸化作用,激活PLC 途徑后產生IP3,隨后Ca2+從內質網中釋放出來;同時鈣池調控鈣離子通道(SOC)打開,細胞外的Ca2+內流。Ca2+濃度增加和cAMP 的增加促進了EECs 的分泌。CD36作為LCFAs 的轉運載體,還可能作為GPR120 的共受體,誘使和轉運LCFA 至低親和力受體GPR120[58]。
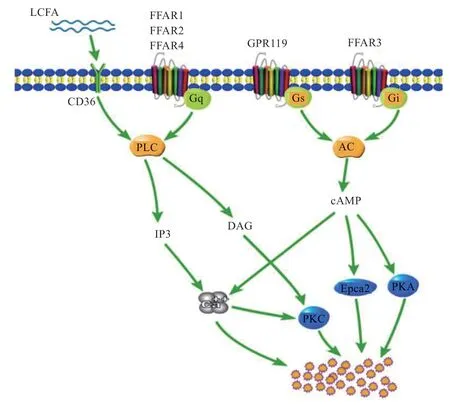
圖1 腸內分泌細胞感應脂肪酸機制[56]
5 小 結
游離脂肪酸與EECs 的相互作用形成了穩定的腸道環境。游離脂肪酸作為飼料添加劑常用于提高肉雞和仔豬的生長性能,但作用方式還不完全清楚。長鏈脂肪酸受體FFAR1 和FFAR4、短鏈脂肪酸受體FFAR2和FFAR3 都分布于腸內分泌細胞上,LCFAs 和SCFAs通過激活這些受體刺激EECs 分泌胃腸激素(CCK、GLP-1、5-HT 和GIP 等)。這些激素通過腸道作用于全身,如EC 細胞分泌的5-HT 可以刺激腸道平滑肌收縮,也可以通過腦-腸軸調節機體大腦活動;L 細胞分泌的GLP-1 在調節胰島素分泌中起著重要作用。因此,了解脂肪酸對畜禽EECs 的作用,深入研究信號途徑,有利于全面了解腸道消化吸收功能的過程。通過調整飼糧的脂肪酸成分,調控胃腸道化學感受器,優化腸道激素的分泌,刺激食欲和調節采食量,對改善畜禽腸道健康、調控生長發育具有重要意義。
