脂肪酸的生理功能及其在畜禽生產中的應用研究進展
2021-11-09 14:18:06王華凱馬永喜
中國畜牧雜志 2021年9期
關鍵詞:研究
張 楠,王華凱,馬永喜*,楊 博
(1.中國農業大學動物科學技術學院,北京 100193;2.華南理工大學生物科學與工程學院,廣東廣州 510006)
抗生素自1920 年被發現以來,以其抗病、促生長的作用在畜禽生產中得到廣泛應用,給畜牧業帶來了巨大價值。但隨著抗生素的長期大量使用,抗藥性、殘留等問題日益凸顯,嚴重威脅了畜產品的質量和公眾健康。2006 年開始歐盟禁止在飼料中添加促生長類抗生素,我國也自2020 年7 月起,飼料生產企業停止生產含有促生長類藥物飼料添加劑(中草藥除外)的商品飼料。因此,研發抗生素替代品成為養殖業的必然選擇。脂類是油、脂肪、類脂的總稱,是由甘油和脂肪酸組成的三酰甘油酯。脂類具有提供能量、改善飼料適口性、促進脂溶性維生素吸收等多種作用。脂類的生理作用與其脂肪酸的組成和比例密切相關,但目前對不同類型脂肪酸的作用、適宜添加比例等研究還有所欠缺。因此,本文介紹了不同類型脂肪酸的生理功能,總結了脂肪酸在畜禽生產中的應用研究進展,以期為開發功能性脂肪酸產品提供理論依據。
1 短鏈脂肪酸(SCFAs)
SCFAs 又稱揮發性脂肪酸,是指碳原子數≤6 的脂肪酸,內源性SCFAs 是由結腸微生物利用抗性淀粉、非淀粉性多糖、低聚糖等難以消化的結構性碳水化合物發酵產生,以乙酸、丙酸、丁酸含量最高。乙酸和丙酸吸收后經門靜脈運送到肝臟,主要參與能量代謝,丁酸則主要是為腸黏膜細胞供能[1]。此外,丁酸還具有維護腸上皮屏障、調節腸道微生態平衡、抗炎等多種生理功能。
1.1 SCFAs 的生理功能
1.1.1 提供能量和電解質平衡 結腸內發酵產生的SCFAs可為結腸上皮細胞提供60%~70%的能量,SCFAs 為細胞供能的順序為丁酸>丙酸>乙酸,其中丁酸含量占腸道SCFAs 總量的85%[2]。除供能以外,SCFAs 還可以刺激結腸Na+吸收,抑制環磷酸腺苷(cAMP)和環磷酸鳥苷(cGMP)介導的Cl-分泌來調節電解質平衡[3]。
1.1.2 腸黏膜屏障作用 腸黏膜屏障由機械屏障、免疫屏障、化學屏障和生物屏障組成,共同維持畜禽的腸道健康,SCFAs 對維持腸黏膜屏障至關重要。SCFAs 可以通過促進黏液層分泌黏液、增加緊密連接蛋白的數量來加強機械屏障。丁酸鹽可以通過上調緊密連接蛋白claudin-1 的轉錄進而增強腸上皮屏障功能[4]。此外,SCFAs 可促進上皮細胞內一些黏膜免疫相關細胞的增殖、免疫球蛋白的分泌來提高腸黏膜免疫屏障。研究發現,丁酸鈉不僅可以增加仔豬腸道黏膜中漿細胞IgA+的數量,還可以促進斷奶仔豬小腸上皮內淋巴細胞、肥大細胞和杯狀細胞的增殖,改善小腸黏膜上皮細胞的形態結構,維持正常的腸道黏膜屏障功能[5-6]。
1.1.3 抗炎、抗菌作用 SCFAs 可以通過抑制炎癥信號通路絲裂原活化蛋白激酶(MAPK)介導的信號轉導,阻斷炎癥信號通路,進而抑制炎癥反應。Xu 等[7]研究發現,給新生豬每日注射7~13 mL 丁酸鈉,連續注射7 d后,白細胞介素6(IL-6)、白細胞介素8(IL-8)、γ-干擾素(IFN-γ)、腫瘤壞死因子β(TNF-β)的表達顯著降低,同時回腸組蛋白去乙酰化酶抑制劑(HDACi)含量也降低。這表明SCFAs 還可作為HDACi 的抑制劑,進而抑制下游相關促炎因子的mRNA 表達從而起到抗炎效果。此外,動物試驗研究表明,在斷奶仔豬日糧中添加450 mg/kg 丁酸鈉降低了空腸脫粒肥大細胞中組胺、類胰蛋白酶的濃度以及腫瘤壞死因子-α(TNF-α)及IL-6 mRNA 的表達量[8]。Jiang 等[9]研究發現,在脂多糖應激下,添加丁酸鈉可以降低十二指腸黏膜過氧化物酶活性和TNF-α的mRNA 表達量。
腸道菌群平衡是腸道健康的重要特征之一,SCFAs進入細菌細胞內可被分解為酸根離子和H+,H+濃度升高,腸道pH 降低,從而抑制大腸桿菌、沙門氏菌等有害菌的生長,促進乳酸菌等有益菌的生長,進而維持動物腸道健康。丁瑩[10]研究發現,在飲水中添加0.15%SCFAs 可以顯著降低42 日齡肉雞盲腸中大腸桿菌數量。鈕海華[6]研究發現,丁酸鈉可以顯著增加仔豬空腸內乳酸菌數量,顯著降低大腸桿菌數量。此外,丁酸還可通過促進黏蛋白分泌黏液從而減少有害菌的黏附[11]。
1.1.4 抑制癌細胞生長 在細胞核中,組蛋白乙酰化和去乙酰化過程處于動態平衡,并由組蛋白乙酰化酶(HAT)和組蛋白去乙酰化酶(HDAC)共同作用,丁酸作為HDACi,可以抑制HDAC 的活性,進而可以調節基因的表達起到抗癌作用[12]。
1.2 SCFAs 在畜禽上的應用 SCFAs 及其衍生物在畜禽生產中的應用研究見表1,SCFAs 及其衍生物在豬方面的研究多集中在斷奶仔豬上。SCFAs 及其衍生物可以提高斷奶仔豬對粗蛋白質、粗脂肪等營養物質的消化率,提高飼料轉化效率、平均日增重和平均日采食量,進而提高生長性能。SCFAs 及其衍生物還可提高斷奶仔豬腸道絨毛高度、絨毛隱窩比值,降低隱窩深度,維持腸黏膜形態結構的完整,促進腸道健康。在母豬妊娠后期及哺乳期添加SCFAs 及其衍生物可以提高母豬的繁殖性能及后代仔豬的生長性能。在家禽方面,SCFAs 及其衍生物可以降低肉雞血清中IL-6、TNF-α等抗炎因子含量,提高超氧化物歧化酶等抗氧化酶活性,進而提高其免疫應答和抗氧化能力。
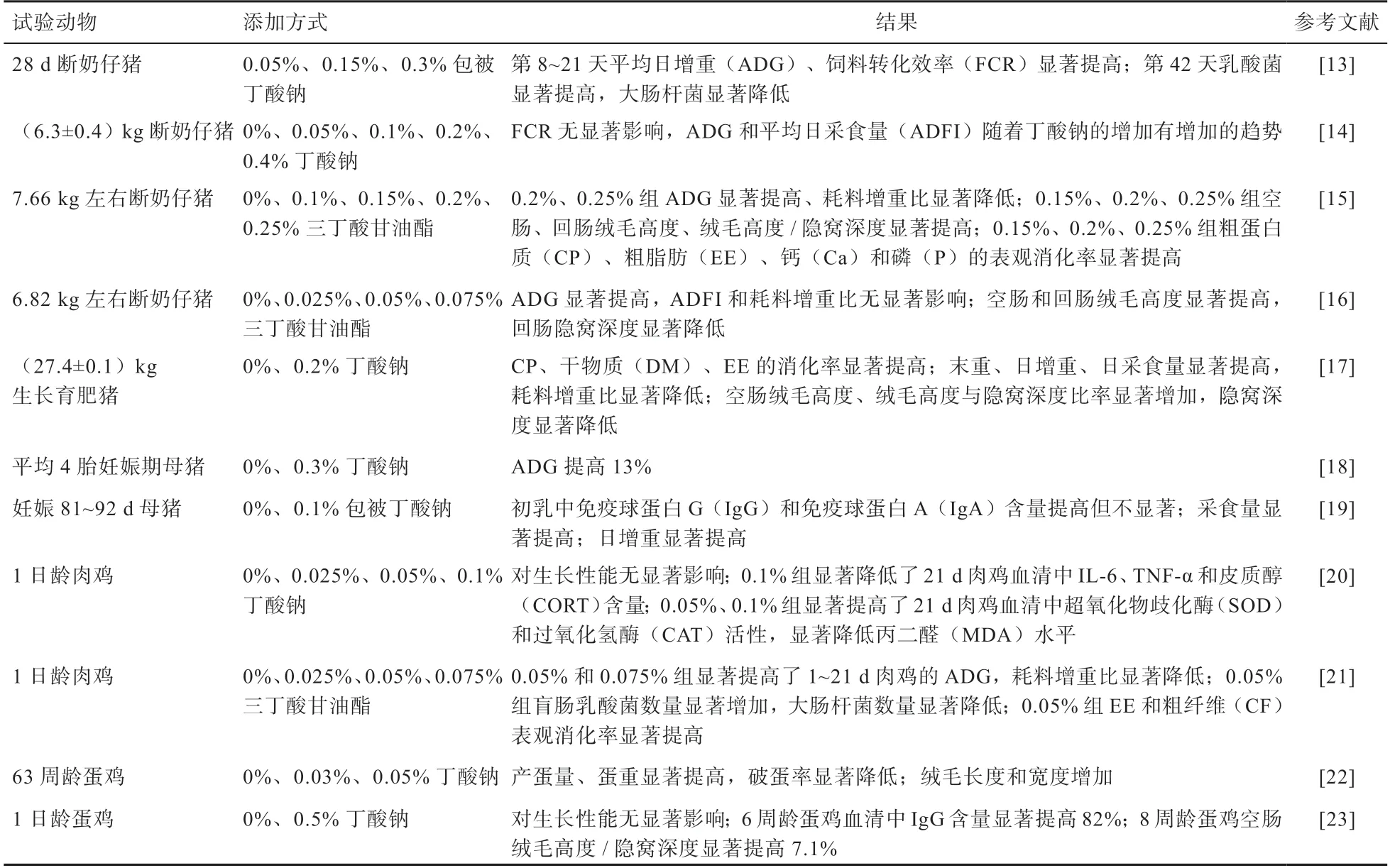
表1 SCFAs 及衍生物在畜禽生產中的應用
綜上所述,在畜禽飼糧中添加0.2%~0.3% SCFAs效果最佳。SCFAs 除具有上述生理功能外,還具有無殘留、易吸收等優點。目前以丁酸及丁酸鈉研究較多,但丁酸鈉存在臭味大、難以為后腸提供丁酸、包被丁酸鈉成本高、含量低等缺陷。丁酸甘油酯是一種丁酸衍生物,其具有在腸道各部分均可釋放丁酸且無臭味等優勢[1]。今后,可加強丁酸甘油酯對不同生理階段畜禽的使用效果研究,還要重視乙酸、丙酸等多種SCFAs 混合作用的效果,以及SCFAs 不同添加方式(如飼糧或飲水中)的應用效果。
2 中鏈脂肪酸(MCFAs)
MCFAs 是指碳原子數為6~12 的飽和脂肪酸,包括己酸(C6:0)、辛酸(C8:0)、癸酸(C10:0)和月桂酸(C12:0)。MCFAs 與甘油發生酯化反應生成相應的中鏈甘油三酯(MCT),從而在自然界中以MCT 的形式存在于動物的乳脂和一些飼料原料中,其中以椰子油、棕櫚油和萼距花屬植物種子油含量較為豐富。MCFAs與長鏈脂肪酸(LCFAs)的性質比較見表2[24]。MCFAs因其特殊的化學及物理特性,在消化、吸收及轉運等方面與LCFAs 存在較大差異。MCFAs 具有調節代謝、提高免疫力、抑菌等多種作用,是最具有發展前景的抗生素替代品之一。
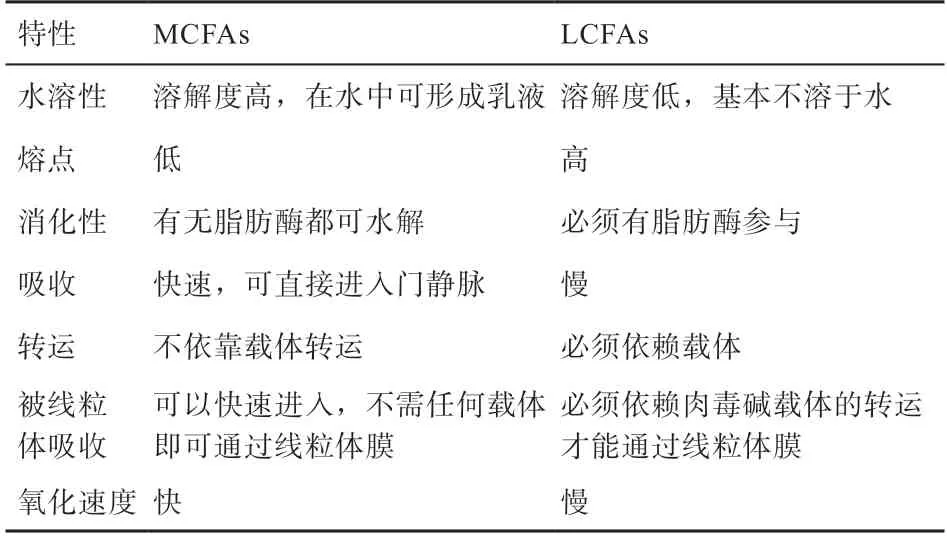
表2 MCFAs 與LCFAs 的性質比較
2.1 MCFAs 的生理功能
2.1.1 調節代謝 MCFAs 可以通過抑制脂肪酸合成酶、過氧化物酶體增殖物激活受體(PPARγ)、增強子結合蛋白(CCAAT)等脂肪酸合成相關基因來調節脂類代謝。MCFAs 還可通過提高動物胰島素敏感性和葡萄糖耐受性,從而參與糖代謝的改善[25]。
2.1.2 調節腸道微生態平衡 MCFAs 具有很強的抑菌作用,其機制可能是MCFAs 通過誘導一種自溶酶的分泌,進而引起細菌溶解和死亡。游離的MCFAs 還可以通過病原體細胞膜進入細胞內并產生酸化作用,引起細菌死亡,同時分解的MCFAs 會抑制病原菌DNA 的復制,進而殺死病原菌[26]。
2.1.3 抗病毒 MCFAs 的單甘脂具有抗病毒的功能,以月桂酸單甘油酯(GML)作用最為突出。GML 通過插入病毒的囊膜,使病毒的膜蛋白外泄,從而使病毒失去感染力或是通過降低相關促炎細胞因子、趨化因子的表達,間接減少病毒的傳播[27-28]。Li 等[27]研究表明,GML 可以降低IL-8 和巨噬細胞炎性蛋白-3α(MIP-3α)的表達,抑制CD4+T 細胞的募集,從而有效阻止恒河獼猴感染猴免疫缺陷病毒(SIV)。在畜禽生產中,GML 通過降低豬繁殖與呼吸綜合征病毒重組N 蛋白(PRRSV-N)在猴胚胎腎上皮細胞-145(MARC-145)中的表達來抑制PRRSV 的增殖,從而起到抗PRRSV的作用[28]。
2.2 MCFAs 在畜禽上的應用 MCFAs 及其衍生物在畜禽生產中的應用研究見表3,MCFAs 及其衍生物因其具有快速供能的特點,多應用在幼小動物生產中。除作為畜禽的能源物質,MCFAs 及其衍生物還具有很強的抑菌作用,可以抑制和殺滅病原微生物,同時還可以改善腸道形態,進而促進畜禽的腸道健康。但MCFAs 及其衍生物對畜禽飼料轉化效率、采食量、增重等方面研究結果不一致,大部分研究表明無顯著影響。這與MCFAs 及其衍生物的種類、添加量、畜禽所處的能量狀態等因素有關。
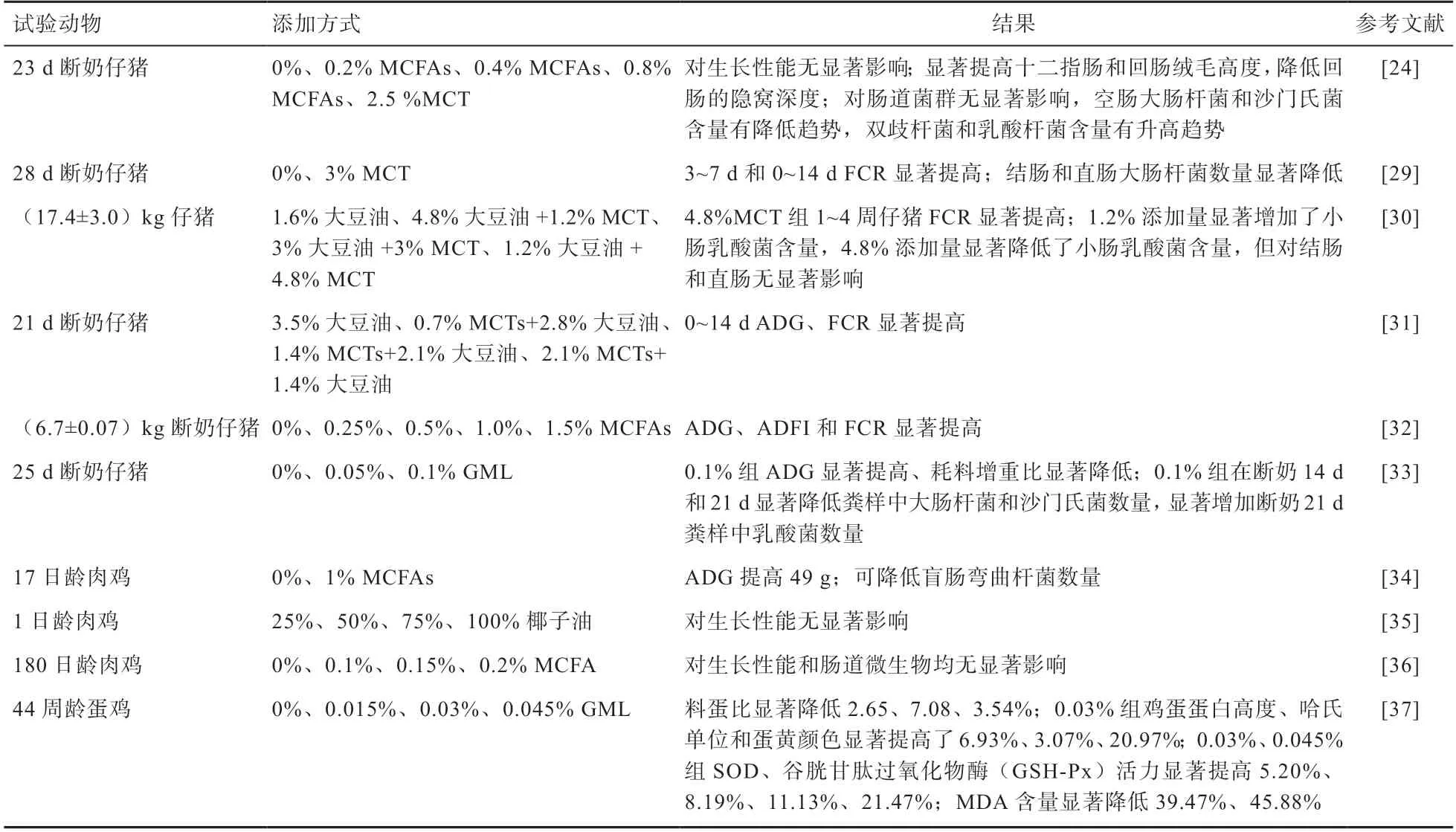
表3 MCFAs 及衍生物在畜禽中的應用
綜上所述,在畜禽飼糧中添加0.2%MCFAs 效果最佳。在今后的研究中應重視MCFAs 與其他產品(如益生菌、植物精油等)聯合使用,以達到畜禽的生長性能最大化。
3 多不飽和脂肪酸(PUFAs)
PUFAs 指含有2 個或2 個以上雙鍵且碳鏈長度為18~22 個碳原子的直鏈脂肪酸,通常分為ω-3 系列和ω-6系列。從脂肪酸的甲基端算起,第3 個碳原子上開始出現雙鍵就是ω-3 脂肪酸,第6 個碳原子上開始出現雙鍵就是ω-6 脂肪酸。ω-3PUFAs 主要包括α-亞麻酸(ALA)、二十碳五稀酸(EPA)、二十二碳六稀酸(DHA),ω-6PUFAs 主要包括亞油酸(LA)、γ-亞麻酸(GLA)、花生四烯酸(AA)。
ω-3PUFAs 和ω-6PUFAs 的功能不同,某些活性甚至相反,例如ω-6PUFAs 中的花生四烯酸是合成促炎類二十烷酸類激素的前體,而ω-3PUFAs 中的EPA 則合成抗炎類二十烷酸類激素。因此,注重二者的平衡才能有利于動物的健康[1]。
3.1 PUFAs 的生理功能 AA 產生的前列腺素E2(PGE2)是促炎因子,而ω-3PUFAs 可通過競爭5-環氧合酶抑制AA 的產生,減少PGE2 濃度,從而減緩炎癥反應[38]。ω-3PUFAs 可以降低TNF-α、核因子-κB(NF-κB)等促炎因子的表達水平、提高白細胞介素10(IL-10)等抗炎因子表達而抑制炎癥反應。此外,ω-3PUFAs 的活性代謝物(如消退素可以刺激巨噬細胞等)分泌產生抗炎性細胞因子來起到抗炎作用[39]。
PUFAs 可以降低血液中甘油三酯和膽固醇濃度,通過抑制血小板凝集防止形成動脈血栓減少心血管疾病[40]。PUFAs 也可通過影響轉錄因子和信號轉導,改變細胞膜功能,通過細胞中脂質過氧化等途徑來預防和治療癌癥。
3.2 PUFAs 在畜禽上的應用 PUFAs 在畜禽生產中的應用研究見表4。在妊娠后期和泌乳期飼糧中添加ω-3PUFAs可以提高母豬初乳中免疫球蛋白的數量,減少仔豬腸道黏膜抗炎因子的mRNA 表達量,減緩母豬和仔豬的應激,進而提高母豬的繁殖性能和仔豬的生長性能,對于斷奶仔豬而言,ω-6/ω-3 的比值會顯著影響斷奶仔豬的生長性能和免疫能力,比值為10 時效果最佳。在家禽方面,PUFAs 可以提高肉雞的生長性能,添加量為150~250 mg/kg 時可以提高蛋雞血清中免疫球蛋白數量,從而提高免疫力。
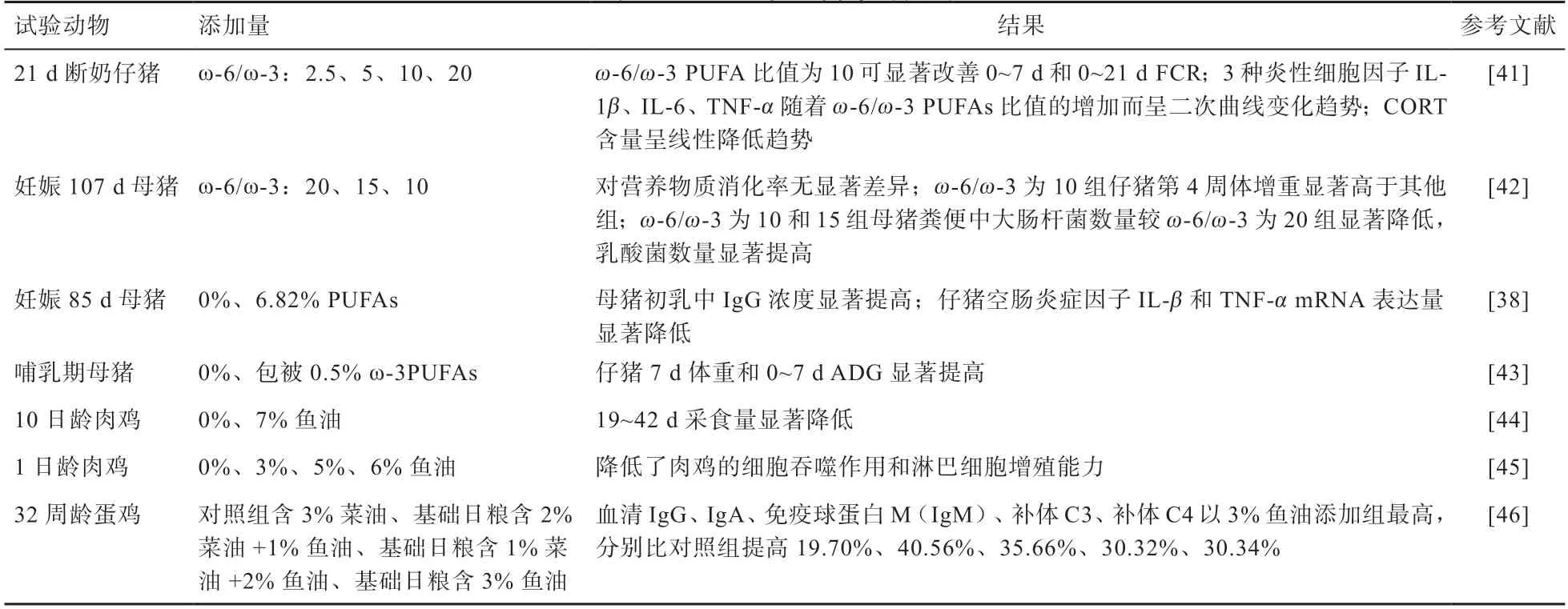
表4 PUFAs 在畜禽中的應用
綜上所述,在飼糧中添加PUFAs 可以抑制畜禽腸道有害菌的增殖,改善腸道形態,提高免疫力,從而提高其生長性能。在自然界中,ω-6PUFAs 含量非常豐富,但只有少數油脂中含有ω-3PUFAs,在飼料中應用大豆油較多,應當在飼料中適當的添加魚油等富含ω-3PUFAs 的油脂。只有當二者的比例處于合理的范圍內,才能使脂肪酸的價值最大化。
4 共軛亞油酸(CLA)
CLA 是指一系列含有共軛雙鍵、具有位置和幾何異構的十八碳二烯酸的統稱。CLA 的異構體種類眾多,其中以c9,t11-CLA 和t10,c12-CLA 應用最為廣泛。天然CLA 主要存在于反芻動物的奶制品和肉制品中。但由于天然CLA 含量較少,人們開始通過人工合成CLA的途徑來獲取更多的CLA 以滿足需求。目前人工合成CLA 的方法有光敏異構法、化學合成法、微生物合成法等,但人工合成成本高、工藝復雜,生產工藝還需進一步優化。CLA 具有抗氧化、調節免疫、抑制脂肪積累、降低膽固醇防止動脈粥樣硬化、抗癌等作用。
4.1 CLA 的生理功能
4.1.1 調節免疫 CLA 通過降低類二十烷酸的合成,特別是前列腺中的PGE2 來調節免疫,PGE2 能夠抑制細胞免疫和細胞因子的分泌,對生物機體免疫功能發揮著重要作用[47]。Zhang 等[48]研究表明,與對照組相比,在雞日糧中添加10.0 g/kg CLA,系統和外周血淋巴細胞中PGE2 的合成量顯著降低57% 和42%。CLA 也可通過細胞核PPARγ途徑抑制TNF-α、NF-κB 等因子的產生,從而起到調節免疫的作用。
4.1.2 防止動脈粥樣硬化及調節脂類代謝 血液中膽固醇含量過高是導致動脈粥樣硬化的主要原因,CLA 能夠阻止脂肪和血小板在粥樣病變的動脈壁上沉積,從而起到抗動脈粥樣硬化的作用[49]。CLA 在以兔子、倉鼠、老鼠為動物模型上的研究較多[50]。Lee 等[51]研究表明,在兔子日糧中每天添加0.5 g CLA,低密度脂蛋白和三酰甘油減少,對主動脈進行檢查,動脈粥樣硬化較少。田敏等[52]研究表明,CLA 可以顯著降低Apo E-/-小鼠血清中總膽固醇、甘油三酯水平,從而起到抗動脈粥樣硬化炎癥反應的作用。
CLA 通過影響脂肪細胞增殖和分化、相關基因的表達及脂代謝中關鍵酶來影響脂肪沉積。陸春瑞等[53]研究表明,與對照組相比,CLA 腹部脂肪參數下降20.93%,腹部脂肪組織中生長激素受體(GHR)和PPARγ的mRNA 表達量分別下降24.74% 和66.52%。Jiang 等[54]研究表明,CLA 降低了背部脂肪△9去飽和酶、脂肪酸合成酶和脂蛋白脂肪酶的活性。
4.1.3 抗氧化 CLA 通過提高TSOD、GSH-Px、CAT 等含量,降低MDA 含量,清除自由基,起到抗氧化的作用。Zhang 等[55]研究表明,10 g/kg CLA 可以提高肉雞肝臟、血清和雞肉中TSOD 的活性及肝臟中CAT 活性,10 g/kg和5 g/kg CLA 可降低肉雞肝臟、血清和雞肉中MDA含量,這與吳國玲[56]的研究結果一致。
4.2 CLA 在畜禽上的應用 CLA 在畜禽生產中的應用研究見表5。在妊娠、哺乳期母豬日糧中添加CLA 可以提高母豬初乳和新生仔豬血清中免疫球蛋白的數量,從而提高新生仔豬的免疫力。在家禽方面,CLA 對肉雞生產性能的研究結果不一致。有的研究表明,CLA 會降低肉雞的ADFI 和ADG,這可能與肉雞的生長階段、CLA 添加量等有關。但可以提高蛋黃中多不飽和脂肪酸含量,降低單不飽和脂肪酸含量,使蛋黃中富集CLA。
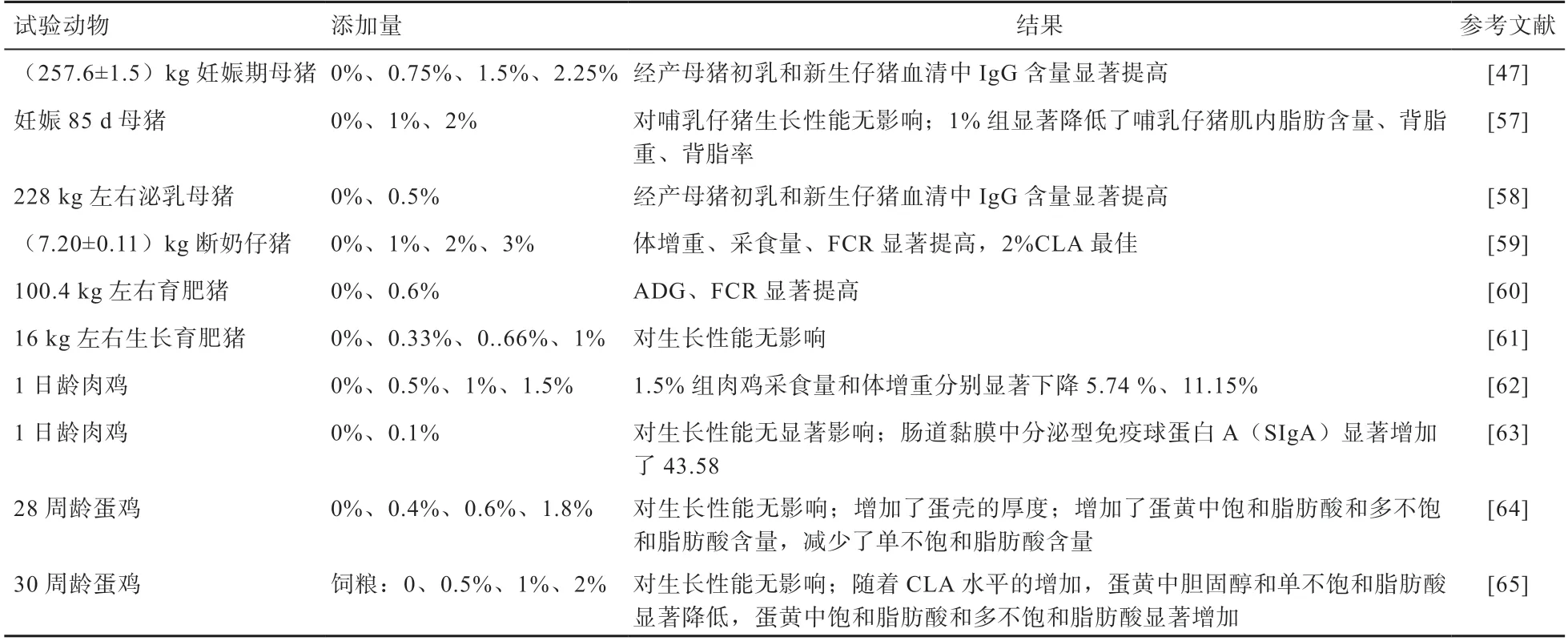
表5 CLA 在畜禽中的應用
綜上所述,CLA 作為一種功能性脂肪酸,在畜禽日糧中以2% 的添加量效果最佳。但CLA 產品現多為游離脂肪酸型,其容易氧化產生對人體有害的過氧化物,且口感不好。與CLA 相比,共軛亞油酸甘油酯穩定性好,在人和動物機體內消化吸收速度更快,具有更好的親脂性,可防止脂質的積累[66],具有廣闊的市場前景。
5 小 結
綜上所述,MCFAs 具有無殘留、易吸收、快速供能等優點,但存在異味、難以為后腸道利用等問題,可以通過與其他飼料添加劑聯合使用或優化添加方式來克服其缺點;合理的ω-6/ω-3PUFAs 比值可以調控畜產品的品質,但富含ω-6PUFAs 和ω-3PUFAs 的產品容易氧化,使用時需要確保飼糧當中的抗氧化劑水平;CLA具有抗動脈粥樣硬化、促免疫等多種生理功能,但目前CLA 制品存在口感不佳、穩定差、易氧化變質等缺點,可以對CLA 的結構進行改造,合成CLA 酯類衍生物來提高CLA 的穩定性。在畜禽飼糧中添加適量脂肪酸可以調節畜禽腸道微生態平衡,增強畜禽的免疫及抗氧化能力,從而提高其生長性能。但不同種類、生理階段畜禽的適宜添加量、添加形式以及聯合的使用效果還需進一步研究。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
