轉基因甘蔗BtG-2的T-DNA側翼序列分析及其轉化事件特異性檢測
2021-11-08 01:03:08馮翠蓮萬玥馮小艷王俊剛趙婷婷王文治沈林波張樹珍
熱帶作物學報 2021年9期
馮翠蓮 萬玥 馮小艷 王俊剛 趙婷婷 王文治 沈林波 張樹珍
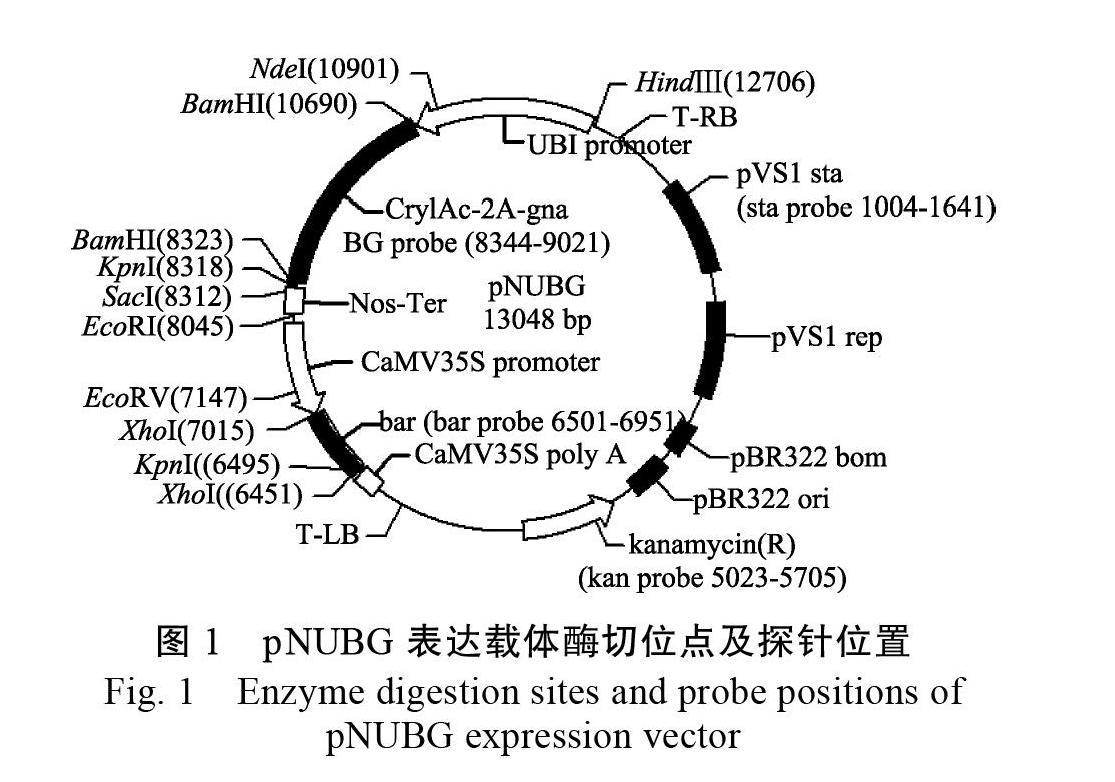

摘? 要:轉基因甘蔗BtG-2是利用農桿菌介導法把Cry1Ac-2A-gna融合抗蟲基因導入‘新臺糖22號的轉基因甘蔗株系,具有良好的抗蟲特性和農藝性狀。為了明確轉基因甘蔗BtG-2的分子特征及其檢測方法,推進其生物安全性評價工作,以BtG-2的T2代為研究材料,利用Southern雜交檢測外源基因在轉基因甘蔗基因組內的拷貝數;利用染色體步移技術分離外源基因在甘蔗基因組中插入位點的側翼序列,并建立了該轉化體高效靈敏的特異性PCR檢測方法。結果表明:Southern雜交檢測證明外源T-DNA以單拷貝方式插入BtG-2株系;經過3次的熱不對稱巢式PCR擴增,獲得外源基因T-DNA左邊側翼序列984 bp和右邊側翼序列705 bp;以這2個序列和相應的T-DNA的左右端序列分別設計3對檢測引物對,建立了BtG-2株系的轉化事件特異性PCR檢測方法,擴增效率最高的引物對LS011/LA451和RS160/RA588分別擴增到440 bp和428 bp的特異片段。其中T-DNA左側設計的LS011/LA451檢測引物對擴增的靈敏度高、特異性強,能夠在甘蔗BtG-2基因組DNA相對含量為0.1%的模板中檢測出轉基因目的成分,相當于9個單倍體基因組拷貝數。本研究完成了轉基因株系BtG-2的分子特征及其轉化事件特異性檢測,為該轉基因甘蔗及其衍生產品的檢測和身份識別提供技術依據。
關鍵詞:抗蟲轉基因甘蔗;T-DNA側翼序列;轉化事件特異性檢測;染色體步移
中圖分類號:S961.6????? 文獻標識碼:A
Analysis of the T-DNA Flanking Sequence and Event-specific Detection for Insect-resistant Transgenic Sugarcane BtG-2
FENG Cuilian1, WAN Yue2, FENG Xiaoyan1, WANG Jungang1, ZHAO Tingting1, WANG Wenzhi1, SHEN Linbo1, ZHANG Shuzhen1,2*
1. Institute of Tropical Bioscience and Biotechnology, Sugarcane Research Center, Chinese Academy of Tropical Agricultural Sciences / Key Biotechnology Laboratory for Tropical Crops, Ministry of Agriculture and Rural Affairs, Haikou, Hainan 571101, China; 2. College of Life Science, Nanjing Agriculture University, Nanjing, Jiangsu 210095, China
Abstract: Sugarcane BtG-2 is an insect resistance transgenic sugarcane strain, developed by introducing the Cry1Ac- 2A-gna fusion gene into ‘ROC22 with the Agrobacterium-mediated method. It has strong insect resistance and excellent agronomic traits. In order to clarify the molecular characteristics and detection of transgenic sugarcane BtG-2, and promote biological safety evaluation, the T2 generation of BtG-2 was selected, and the copy number of foreign genes in the transgenic sugarcane genome was detected by Southern hybridization. The flanking sequence of the insertion site of the foreign gene was isolated using the chromosome walking technology, and an efficient specific PCR detection method of the strain was established. The results showed that the foreign T-DNA insertion of BtG-2 strain was a single copy. After three times amplifications of thermal asymmetric interlaced PCR, 984 bp of the left flanking sequence and 705 bp of the right flanking sequence of the foreign gene T-DNA were obtained. According to the flanking sequences, three pairs of detection primers were designed respectively, then the event-specific PCR detection for transgenic sugarcane BtG-2 was established. The primer pairs with the highest amplification efficiency were LS011/LA451 and RS160/RA588, with 440 bp and 428 bp specific amplified fragments respectively. Among them, the pair of primers LS011/LA451 designed on the left side of T-DNA had high sensitivity and specificity for detection, and this method could detect the genetically modified ingredients in samples containing 0.1% genomic DNA of sugarcane BtG-2. This study completed the molecular characteristics and event-specific detection of the transgenic strain BtG-2, which provided a technical basis for the detection and identification of the transgenic sugarcane and its derivatives.
Keywords: Insect-resistant transgenic sugarcane; T-DNA flanking sequence; event-specific detection; chromosome walking
DOI: 10.3969/j.issn.1000-2561.2021.09.005
甘蔗(Saccharum spp.)是全球第五大宗農作物,是世界上最重要糖料和生物燃料作物。中國糖業協會數據顯示:2020年全國種植甘蔗119.1萬hm2,產糖量841.9萬t,農業直接年產值約480億人民幣[1],因此甘蔗產業對國民經濟具有重要的意義。然而甘蔗受各種螟蟲的為害,嚴重影響甘蔗的產量和質量,僅以湘桂蔗區為例,2018—2019年,該區的甘蔗蟲害發生極為嚴重,平均螟害株率高達46.54%,平均產量損失為14.87%,平均蔗糖分損失高達1.12個百分點,農業損失4.67億元,工業損失8.56億元,國家財政損失0.51億元[2]。因此培育抗蟲甘蔗新種質一直是甘蔗育種的一個重要目標,然而甘蔗是高度雜合的無性繁殖作物,其龐大的基因組、復雜的高多倍體以及同源異源雜交品種等復雜的遺傳背景[3],給育種工作帶來很大的盲目性。基因工程可定向改變作物的某些性狀,通過轉基因技術提高甘蔗抗蟲性將是甘蔗抗蟲育種的重要新途徑。
在轉基因作物中,T-DNA在受體植物基因組中的整合位置都是隨機的,然而每個轉基因事件的T-DNA左右端序列與受體基因組序列拼接而成的T-DNA插入位點側翼序列是唯一的,是該轉基因事件身份的特異性標識[4]。因此,分離轉基因植物的T-DNA側翼序列,以及依據該側翼序列而建立的特異性檢測方法,是準確識別不同轉基因作物,實現轉基因作物及其產品產權保護、檢測和有效監督管理的一個重要依據。
目前分離外源T-DNA側翼序列主要以PCR技術為基礎,包括反向PCR、外源接頭介導PCR、半隨機引物PCR、全基因組重測序技術等,其中半隨機引物PCR中的熱不對稱交錯PCR(TAIL-PCR)、高效熱不對稱PCR(hiTAIL-PCR)或染色體步移法(Genome Walking)是當前使用最廣泛的方法。利用此類技術已經成功分離了轉基因水稻[4-8]、大豆[9-11]、棉花[12-13]、玉米[14-17]、小麥[18-20]、馬鈴薯[21-22]、油菜[23]和木瓜[24]等T-DNA側翼序列,并建立其相應的轉化事件特異性檢測技術;全基因組重測序是近年發展起來的新技術,是對已知基因組序列的物種進行不同個體的基因組測序,并在此基礎上對個體或群體進行差異性分析,主要用于檢測單核苷酸多態性位點(SNP)、插入缺失位點(InDel)、結構變異位點(SV)位點和拷貝數變異位點(CNV)[25]。近兩年該技術也成功應用到分離轉基因作物大豆[26-29]、水稻[30]的T-DNA側翼序列上,并建立其相應的轉化事件特異性檢測技術。轉基因作物轉化事件特異性檢測技術是以其轉基因T-DNA側翼序列的部分序列為靶標進行擴增,擴增產物是T-DNA 5端或3端序列與受體基因組的拼接序列,因此具有高度特異性,可準確識別不同的轉基因作物品系。
轉基因甘蔗的研究始于20世紀90年代初,經過近30年的努力,取得了很大的成果。截至目前,據國際農業生物技術應用服務組織(ISAAA)的數據,獲得安全證書的甘蔗轉基因事件僅有6例,分別是巴西2017年獲批的轉Cry1Ab基因的抗蟲轉基因甘蔗CTB141175/01-A和2018年獲批的轉Cry1Ac基因的CTC91087-6和CTC93209-4,印度尼西亞2011年獲批的轉大腸桿菌的膽堿脫氫酶(EcBetA)基因耐旱轉基因甘蔗的NXI-1T,以及2013年獲批的轉苜蓿根瘤菌的膽堿脫氫酶(RmBetA)基因耐旱轉基因甘蔗的NXI-4T、NXI-6T[31]。由此可見,轉基因甘蔗的研究遠遠落后于其他作物。目前轉基因甘蔗的研究重點仍在遺傳轉化等前期工作上,轉基因甘蔗的生物安全評價以及檢測、監督等后期工作的研究極少有報道,尤其轉化事件特異性檢測國內外尚未有報道。
本研究組前期通過農桿菌介導把Cry1Ac- 2A-gna抗蟲融合基因轉入甘蔗新臺糖22號,經過2個無性繁殖世代的抗蟲性和遺傳穩定性的評價和深入農藝性狀鑒定(待發表),已獲得抗蟲性好、農藝性狀優良的BtG-2株系,正準備進行環境釋放。為了明確轉基因甘蔗BtG-2的分子特征及便于其檢測,推進其生物安全性評價工作,本研究以其無性繁殖T2代為植物材料,首先利用Southern雜交檢測外源T-DNA在BtG-2中的插入拷貝數;然后利用染色體步移技術分離其T-DNA側翼序列,并根據T-DNA左右端序列和左右側翼序列設計檢測引物對,建立轉基因甘蔗BtG-2的轉化事件特異性檢測方法,同時檢驗該方法的特異性與靈敏度,為該轉基因甘蔗及其衍生產品的檢測和身份識別提供技術依據。
1? 材料與方法
1.1? 材料
植物材料為轉Cry1c-2A-gna融合抗蟲基因和bar抗除草劑基因的甘蔗BtG-2株系以及BtG-1、BtG-4、BtG-17、BtG-32、BtG-35、BtG-36和BtG-41是通過農桿菌介導轉化甘蔗‘新臺糖22號(‘ROC22)胚性愈傷而獲得,由本實驗室保存,BtG-2株系已申請在海南省的中間試驗。植物表達載體以pCambia3300為骨架。PCR DIG Probe Synthesis Kit、Dig nucleic acid detection kit購于德國羅氏公司;HyboodTM-N+尼龍膜購自美國GE Amersham;試劑LA Taq酶、Genome Walking Kit、pMDTM18-T Vector Cloning Kit、T4 DNA Ligase、部分Marker購于TaKaRa公司;質粒提取、膠回收試劑盒及純化試劑盒購于Axygen公司;氨芐青霉素、瓊脂糖購于Sigma公司;2×Taq plus MasterMix購于Biosharp公司。PCR引物合成及基因測序由上海生工生物工程技術服務有限公司完成。其他試劑均為進口或國產分析純。
1.2? 方法
1.2.1? 甘蔗BtG-2基因組DNA的提取? 選擇轉基因甘蔗BtG-2的無性繁殖T2代植株的幼嫩葉片,采用改良CTAB法大量提取并純化基因組總DNA[32],同時提取非轉基因甘蔗ROC22基因組總DNA作為對照。取2 μL于Thermo Scientific Nano Drop One生物核酸定量檢測儀測定DNA的濃度和純度,另取2 μL于1.0%瓊脂糖凝膠電泳檢測DNA的完整性。
1.2.2? Southern雜交檢測? 以轉化質粒pNUBG為模板,利用高效PCR地高辛標記法分別制備BG(包括Cry1Ac-2a-gna中Cry1Ac基因下游355 bp,2a和gna全長)、bar、sta、kan四種不同的探針,擴增引物、退火溫度、產物長度等詳見表1。通過分析T-DNA區的酶切位點,結合各探針在T-DNA上的位置(圖1),分別選擇2種限制性內切酶對30 μg基因組DNA進行酶切(表1),然后轉移至HyboodTM-N+尼龍膜,再利用相應的探針進行Southern雜交檢測,具體步驟參照試劑盒說明書。雜交溫度為40 ℃,預雜交時間為2 h,雜交時間為16 h,室溫黑暗條件下用BCIP/NBT進行化學顯色,待雜交信號帶清晰后終止顯色,拍照并分析[33]。
1.2.3? 轉基因甘蔗BtG-2的T-DNA左右側翼序列的獲取? 按照Genome Walking Kit中的要求,在表達載體pNUBG的T-DNA左右端序列分別設計3條退火溫度約為65 ℃的嵌套特異性引物(表2),分別命名為Lsp1、Lsp2、Lsp3和Rsp1、Rsp2、Rsp 位置見圖2。按照操作說明,分別選取試劑盒中提供的隨機簡并引物AP1、AP2、AP3、AP4其中之一,與嵌套特異性引物組合配對,進行3輪巢式PCR擴增,分離該株系的T-DNA左右側翼序列。第1輪PCR反應體系中,BtG-2基因組DNA模板為200 ng,特異性引物為Lsp1或Rsp 簡并引物為AP1-AP4其中之一。第2輪PCR反應的擴增模板為上一輪PCR產物稀釋50倍,特異性引物為Lsp2或Rsp2,簡并引物為第1輪所用的AP引物,第3輪同第2輪,特異引物換成Lsp3或Rsp 每一輪反應的退火溫度詳見表2,擴增程序參照試劑盒說明書。3輪擴增產物用1.0%瓊脂糖凝膠電泳分離,第3輪特異擴增片段利用DNA膠回收試劑盒回收。連接T載體,轉化大腸桿菌DH5α,選取陽性菌株送至上海生工生物工程技術服務有限公司測序。
1.2.4? 轉基因甘蔗BtG-2事件特異性PCR檢測及其靈敏度? BtG-2事件特異性PCR檢測的建立:根據T-DNA左右兩側翼序列的測序結果,以及已知的T-DNA左右端載體序列分別設計3對左、右側特異性檢測引物:LS060/LA451、LS011/LA451、LS060/LA585和RS160/RA477、RS129/RA477、RS160/RA588,對轉基因甘蔗BtG-2進行擴增,分別篩選一組左、右特異性檢測擴增效率最好的,并回收純化特異性片段及測序,比對是否與預期序列完全一致,最終建立事件特異性檢測方法。特異性檢測引物對的一條引物位于甘蔗基因組序列,另一條引物則位于T-DNA左、右端載體序列,引物序列詳見表3。分別利用上述篩選的2對引物和bar基因引物,以包括BtG-2在內的轉Cry1Ac-2a-gna基因的不同甘蔗株系和非轉基因甘蔗DNA為模板,以表達載體pUNBG質粒為對照,進行轉化事件特異性PCR檢測。DNA模板量為200 ng,反應循環數35,退火溫度見表3。PCR產物取5 μL于1.0%瓊脂糖凝膠電泳檢測擴增效率和擴增特異性。
檢測轉化事件特異性PCR的靈敏度:轉基因甘蔗BtG-2與受體甘蔗ROC22的基因組DNA分別稀釋至100 ng/μL,根據不同的比例配制BtG-2的DNA相對含量為100%、50%、10%、1%、0.5%、0.1%、0.05%和0%(V/V)的樣品。混合樣品各取1 μL作為PCR模板,分別利用上述已篩選出來的擴增效率最高的左、右特異性引物對,進行PCR擴增。取5 μL PCR產物,用1.0%瓊脂糖凝膠電泳,檢查各濃度DNA模板下的擴增效果,檢測各引物對的事件特異性PCR的特異性和靈敏度。
2? 結果與分析
2.1? 抗蟲轉基因甘蔗BtG-2外源T-DNA插入拷貝數檢測
轉基因甘蔗BtG-2基因組DNA分別用BamHⅠ、NdeⅠ、EcoRⅤ限制性內切酶進行酶切、電泳和轉膜后,利用地高辛標記的外源基因BG和bar片段的探針分別進行Southern雜交檢測。
結果表明,不同限制性內切酶酶切后的甘蔗基因組DNA經BG和bar探針雜交后,均只顯示一條雜交帶,并且雜交帶的大小與預測理論大小一致,而對照非轉基因植株中無雜交信號,說明外源基因已整合到甘蔗BtG-2基因組中,并以單拷貝的方式整合(圖3);用載體骨架序列制備的sta和kan探針進行Southern雜交,結果表明轉基因甘蔗基因組中沒有來源于轉化載體的其他元件以及骨架序列。
2.2? T-DNA左、右側翼序列的獲得及分析
3條左側巢式特異性Lsp引物分別與試劑盒中AP1-AP4四條簡并引物組合,經過3輪熱不對稱巢式PCR反應后,電泳結果顯示,特異性引物與簡并引物AP3擴增出的片段更長且特異性更好(圖4A)。回收3rd產物,測序獲得984 bp的序列(圖5A),其中838 bp屬于甘蔗基因組序列,此部分GC百分含量為48.69%;剩余146 bp與T-DNA左邊載體序列重合,并且最左邊缺失了包括LB border在內的115 bp(圖5B)。
3條右側巢式特異性Rsp引物同樣分別與試劑盒中AP1-AP4四條簡并引物組合擴增右側翼序列,結果顯示,簡并引物AP1擴增出的片段特異性更好(圖4B)。測序獲得705 bp的序列(圖6A),其中566 bp為甘蔗基因組序列,GC百分含量為50.71%,余下139 bp屬于T-DNA右邊界載體序列,T-DNA插入BtG-2株系基因組時,右邊比較完整,僅缺失了2 bp(圖6B)。由此可見,T-DNA序列插入時,左右兩側均有不同程度的缺失,且左邊相對容易缺失。
2.3? 建立轉基因甘蔗BtG-2事件特異性檢測方法
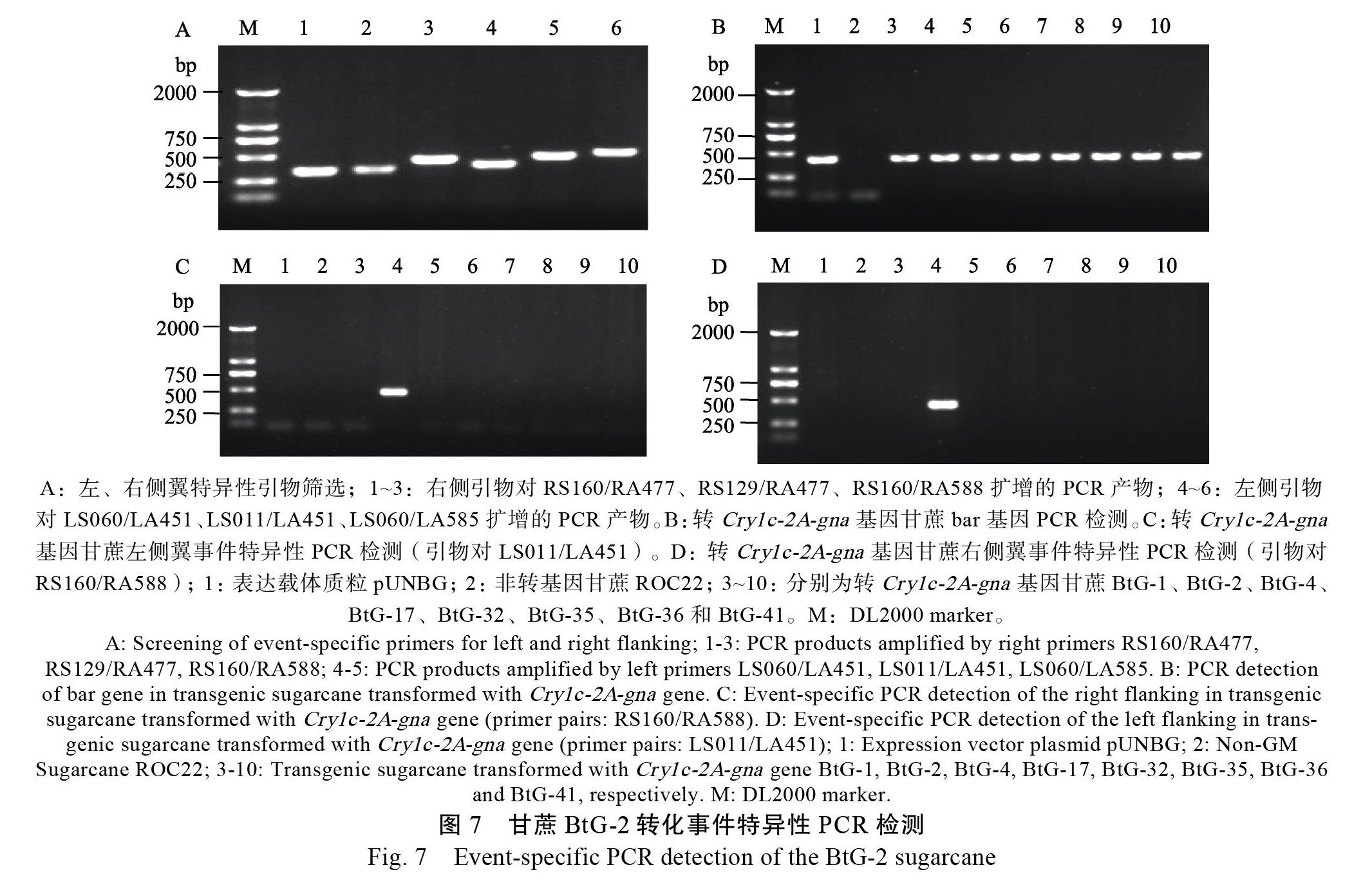
使用左、右端3對特異性檢測引物LS060/ LA451、LS011/LA451、LS060/LA585和RS160/ RA477、RS129/RA477、RS160/RA588,對轉基因甘蔗BtG-2進行擴增,擴增效率最好的左、右特異性檢測引物對分別是LS011/LA451和RS160/ RA588(圖7A)。以轉相同基因的轉基因甘蔗株系和非轉基因甘蔗為模板,分別利用上述2對擴增效率最好引物和bar基因引物進行事件特異性檢測。結果顯示,使用特異性檢測引物對的擴增產物中,只在轉基因甘蔗BtG-2可擴增到大小為440 bp(左側翼)和428 bp(右側翼)的特異性條帶,其他轉基因株系和非轉基因材料擴增均為陰性(圖7C、D);而使用bar基因引物的擴增產物中,表達載體質粒和轉基因各株系均能擴增到約423 bp的特異性片段,只有非轉基因甘蔗擴增結果為陰性(圖7B)。BtG-2擴增到的440 bp和428 bp的特異性片段回收后,經測序比對證實與預期序列完全一致。證明引物對LS011/LA451和RS160/RA588可特異的識別轉基因甘蔗事件BtG-2。
2.4? 事件特異性PCR檢測的靈敏度
利用引物對LS011/LA451和RS160/RA588檢測事件特異性PCR在不同BtG-2的DNA含量混合樣品中的擴增限度。結果發現,以左側引物對LS011/LA451擴增時,當BtG-2的DNA含量降低至0.1%時,仍然可以特異地擴增到目的片段(圖8A);以右側引物對RS160/RA588擴增時,當BtG-2的DNA含量降低至1%和0.5%時,除了目的片段,還出現了非特異片段的擴增,當BtG-2的DNA含量降低至0.1%及更低時,只有非特異片段的擴增(圖8B)。以上結果表明,以右側引物對RS160/RA588的特異性PCR檢測的特異性和靈敏度較差,而以左側引物對LS011/LA451的特異性PCR檢測的特異性好、靈敏度高,最低檢出值達每100 ng DNA模板量的0.1%,按甘蔗基因組為10 Gb計算,相當于9個單倍體基因組拷貝數,該靈敏度可滿足事件特異性檢測的要求。
3? 討論
近年來,轉基因作物種植及應用持續保持穩定增長,2018年,全球26個國家種植了1.917億hm2轉基因作物,比2017年增長1%。2019年,全球共有43項關于轉基因作物的批準,有9個新的轉基因作物品種獲得批準,包括油菜、棉花、豇豆、大豆和甘蔗。而國內農業農村部公示了192個擬頒發的“農業轉基因生物安全證書”植物品種,其中包括189個棉花、2個玉米和1個大豆品種[34]。然而轉基因作物的快速發展,亟需對轉基因作物進行有效的評估與監管。每一個轉化事件中,T-DNA在受體植物的插入都是隨機的,因此,T-DNA側翼序列的克隆及定位對于轉因植株的身份驗證具有重要意義,并有助于對外源基因的表達和外源基因在受體植物中所能產生的影響進行研究,從而有利于對轉基因植株的安全性做出充分評估[5]。
檢測外源基因旁側序列及在受體基因組中的整合位點的方法有很多種,當前應用最廣泛最成功的的是熱不對稱交錯PCR或染色體步移法,以及近幾年發展起來的全基因組重測序新技術。后者是對已知基因組序列的物種進行不同個體的基因組測序,并在此基礎上對個體或群體進行差異性分析,由此可見,應用該技術的前提條件是已知受體植物的基因組序列信息。截至目前,國內外并未對甘蔗ROC22開展基因組測序工作,因此本研究仍然選擇操作比較繁瑣的染色體步移法。染色體步移法的原理和熱不對稱交錯PCR是一致的。本研究中,根據已知T-DNA左右序列設計3條同向Tm值約為65 ℃的特異性引物,與Genome Walking Kit中的4條簡并引物分別進行熱不對稱PCR反應,通過3次巢式PCR反應后,電泳檢測發現,4條簡并引物跟相同的特異性引物組合的擴增效率差異很大,只有簡并引物AP1和右側特異性引物組合、AP3和左側特異性引物組合經過退火溫度、模板濃度等調整可擴增到特異片段,其他組合經過多次調整仍無法得到特異片段,由此推測AP1和AP3簡并引物比較適用于甘蔗。
通過上述染色體步移法獲得了BtG-2的T-DNA側翼序列,上傳至NCBI的核酸數據庫中進行比對,結果顯示沒有任何的同源序列,原因是甘蔗栽培種全基因組測序尚未完成,NCBI的核酸數據庫只有少量的轉錄組測序序列和一些EST序列,因而無法對抗蟲轉基因甘蔗T-DNA插入位點進行精準的定位。另一方面甘蔗割手密種AP85-441基因組測序于2018年完成并發表在《Nature Genetics》上[35],本研究團隊成員把序列下載到本地,進行本地化建庫獲得庫文件,T-DNA側翼序列Blastn比對,發現T-DNA插入位點可能位于該基因組的Chr1B非編碼區92 525 762位點、Chr3B非編碼區8 557 277位點、Chr3D非編碼區7 168 624位點、Chr5C非編碼區38 753 724位點。從上述結果看,外源基因插入位點可能是甘蔗基因組的重復序列區,此重復序列存在于甘蔗割手密基因組多條染色體上;從Southern雜交的結果看,外源基因在甘蔗基因組中只有一個整合位點,而就目前缺少甘蔗基因組序列數據庫的情況下,根據甘蔗某段核酸序列而進行準確的染色體定位是極其困難的,最好的解決方法是加快甘蔗基因組的測序工作。據了解,國內已經有甘蔗研究團隊準備甘蔗ROC22的基因組測序工作,該項工作的完成,勢必將甘蔗基因組功能研究等基礎研究推上一個新的臺階。
通過擴增T-DNA插入片段和受體基因組的連接區域而建立起來的事件特異性檢測法,是鑒定含有相同T-DNA插入片段的不同轉化事件的有效方法,是目前用于轉基因作物及其產品中轉基因成分檢測的最常用方法。本研究建立的轉基因甘蔗BtG-2的事件特異性PCR檢測方法特異性好、靈敏度高,轉基因成分含量0.1%及以上時,利用常規定性PCR方法即可檢測,此檢測靈敏度遠高于歐盟對轉基因植物衍生的食品及飼料標識的最低限量(0.9%)[36]。
4? 結論
本研究利用Southern雜交確定了轉基因甘蔗BtG-2的外源基因的插入拷貝為單拷貝;利用染色體步移方法,擴增得到轉基因甘蔗BtG-2外源基因插入位點的左右側翼序列;以此序列為基礎,建立了轉基因甘蔗BtG-2的事件特異性檢測方法。該方法特異性好、靈敏度高,為轉基因甘蔗BtG-2的利用及產品檢測提供關鍵技術基礎。
參考文獻
[1]? 中國糖業協會. 2019/20年制糖期截至3月底食糖產銷情況簡報[EB/OL]. (2020-04-08)[2020-08-28]. http://www. chinasugar.org.cn/i,35,3430,0.html.
[2]? 謝江江, 楊俊賢, 羅青文, 等. 廣西湘桂蔗區甘蔗蟲害調研及防控對策[J]. 甘蔗糖業, 2019(5): 24-28.
[3]? Zhang J, Zhang X, Tang H, et al. Allele-defined genome of the autopolyploid sugarcane Saccharum pontaneum L.[J]. Nature Genetics, 2018, 50(S1): 1565-1573.
[4]? 崔? 帥, 王作平, 于江輝, 等. 轉基因水稻BPL9K-2事件特異性檢測方法的建立[J]. 中國生物工程雜志, 2018, 38(11): 32-41.
[5]? 金永梅, 馬? 瑞, 于志晶, 等. 轉基因水稻吉生粳2號的外源基因旁側序列分離及事件特異性PCR檢測方法[J]. 東北農業科學, 2016, 41(1): 14-19.
[6]? Dong Y, Jin X, Tang Q, et al. Development and event-spe-cific detection of transgenic glyphosate-resistant rice expressing the G2-EPSPS gene[J]. Frontiers Plant Science, 2017, 8: 885-895.
[7]? 郭? 超, 何行健, 鄧力華, 等. 轉基因水稻 BarKasalath-01事件特異性檢測[J]. 分子植物育種, 2017, 15(11): 4466-4475.
[8]? 金永梅, 馬? 瑞, 于志晶, 等. 轉cry1C基因抗蟲水稻吉生粳3號外源基因整合分析與品系特異性檢測[J]. 生物技術通報, 2019, 35(3): 6-12.
[9]? 包婉瑩, 仲曉芳, 杜? 茜, 等. 抗病轉基因大豆事件B4J8049外源T-DNA整合位點分析及特異性檢測[J]. 東北農業科學, 2018, 43(5): 21-26.
[10]????? 蔡勤安, 尚麗霞, 姜志磊, 等. 轉HAL1基因大豆側翼序列分析及特異性檢測方法研究[J]. 大豆科學, 2018, 37(1): 32-38.
[11]????? Federici S, Paraskevopoulos K. Risk assessment of new sequencing information for genetically modified soybean A2704-12[J]. EFSA Journal, 2018, 16(11): 5496-5503.
[12]???? 陳天子, 凌溪鐵, 楊郁文, 等. 轉GbVe1基因在棉花基因組中的整合與定位分析[J]. 棉花學報, 2019, 31(1): 1-11.
[13]????? 王? 葉, 謝家建, 黃春蒙, 等. 轉cry1Aa基因抗蟲棉整合結構解析及轉化體特異性檢測方法的建立[J]. 棉花學報, 2017, 29(4): 307-315.
[14]????? 郭? 翠, 張? 維, 余桂容, 等. 轉G2-EPSPS基因玉米D-3側翼序列分析與轉化體特異性檢測方法[J]. 作物雜志, 2016(1): 69-75.
[15]????? Rao J, Yang L T, Guo J C, et al. Development of event- specific qualitative and quantitative PCR detection methods for the transgenic maize BVLA430101[J]. European Food Research and Technology, 2016, 242(8): 1277-1284.
[16]????? 王翠云, 劉? 艷, 劉允軍. 外源基因在轉基因玉米中的整合位點分析[J]. 生物技術通報, 2019, 35(3): 1-5.
[17]????? Siddique K, Wei J, Li R, et al. Identification of T-DNA insertion site and flanking sequence of a genetically modified maize event IE09S034 using next-generation sequencing technology[J]. Molecular Biotechnology, 2019, 61(9): 694-702.
[18]????? Zhang M H, Yu Y B, Gao X J, et al. Event-specific quantitative detection of genetically modified wheat B72-8-11 based on the 3 flanking sequence[J]. European Food Research and Technology, 2015, 240(4): 775-782.
[19]????? Zhang P Q, Xu J Y, Zheng Q Y, et al. Flanking sequence determination and event specific detection of transgenic wheat B72-8-11b strain[J]. Applied Biochemistry and Biotechnology, 2013, 169(5): 1523-1530.
[20]????? Xu J Y, Cao J J, Cao D M, et al. Flanking sequence determination and event-specific detection of genetically modified wheat B73-6-1[J]. Acta Biochimicaet Biophysica Sinica, 2013, 45(5): 416-421.
[21]????? 閆建俊, 白云鳳, 左靜靜, 等. 轉基因馬鈴薯外源基因插入位點分析及檢測方法的建立[J]. 分子植物育種, 2020, 18(16): 5361-5366.
[22]????? Gao H, Yu X, Deng T, et al. Event-specific detection of transgenic potato AV43-6-G7 using real-time and digital PCR methods[J]. Bmc Biotechnology, 2016, 16(1): 74-83.
[23]????? Chen S, Shen A J, Zhou X Y, et al. Analysis of the flanking sequence and event-specific detection of transgenic line W-4 of Brassica napus[J]. Journal of Agricultural Science and Technology, 2014, 15(7): 1089-1094.
[24]????? 陳紅運, 黃? 峰, 陳? 青, 等. 應用hiTAIL-PCR擴增轉基因木瓜的側翼序列[J]. 植物檢疫, 2018, 32(5): 24-27.
[25]????? Ley T J, Mardis E R, Ding L, et al. DNA sequencing of a cytogenetically normal acute myeloid leukaemia genome[J]. Nature, 2008, 456(7218): 66-72.
[26]????? Guo B F, Guo Y, Hong H L, et al. Identification of genomic insertion and flanking sequence of G2-EPSPS and GAT transgenes in soybean using whole genome sequencing method[J]. Frontiers in Plant Science, 2016, 7: 1009.
[27]????? 仲曉芳, 楊? 靜, 賀紅利, 等. 基于基因組重測序分析的高含量油酸轉基因大豆事件外源T-DNA旁側序列分析及事件特異性PCR檢測[J]. 農業生物技術學報, 2018, 26(12): 2017-2026.
[28]????? 馬? 闊, 仲曉芳, 牛? 陸, 等. 耐鹽轉基因大豆事件FA8015旁側序列分離及定性PCR檢測[J]. 大豆科學, 2018, 37(6): 854-859.
[29]???? 劉東波, 邢國杰, 趙倩倩, 等. 抗病毒轉基因大豆事件外源T-DNA序列分析及定性檢測[J]. 大豆科學, 2020, 39(1): 23-29.
[30]????? 徐紀明, 胡? 晗, 毛文軒, 等. 利用重測序技術獲取轉基因植物T-DNA插入位點[J]. 遺傳, 2018, 40(8): 676-682.
[31]????? 馮翠蓮, 張樹珍. 抗蟲轉基因甘蔗的培育及其抗性喪失的防控策略[J]. 生物技術通報, 2020, 36(7): 209-219.
[32]????? 王關林, 方宏筠. 植物基因工程[M]. 2版. 北京: 科學出版社, 2002: 744.
[33]????? 崔學強, 張樹珍, 沈林波, 等. 轉基因甘蔗植株Southern雜交體系的優化[J]. 生物技術通報, 2015, 31(12): 105- 109.
[34]????? 2019年全球轉基因觀察[EB/OL]. (2020-01-17) [2020- 08-28]. http://cn.agropages.com/News/NewsDetail 20390.htm.
[35]????? Zhang J, Zhang X, Tang H, et al. Allele-defined genome of the autopolyploid sugarcane Saccharum spontaneum L.[J]. Nature Genetics, 2018, 50(S1): 1565-1573.
[36]????? Marmiroli N, Maestri E, Gulli M, et al. Methods for detection of GMOs in food and feed[J]. Analytical and Bioanalytical Chemistry, 2008, 392(3): 369-384.
責任編輯:黃東杰
