自建Tg.C57-ras V1.0小鼠模型與日本Tg.rasH2小鼠遺傳學(xué)及其雜交1代生物學(xué)特性比較*
2021-11-04 10:48:22劉甦蘇楊艷偉王辰飛王三龍范昌發(fā)
實(shí)驗(yàn)動(dòng)物科學(xué) 2021年4期
劉甦蘇 曹 愿 楊艷偉 王辰飛 王三龍 范昌發(fā)
(1.中國(guó)食品藥品檢定研究院實(shí)驗(yàn)動(dòng)物資源研究所,北京 102629)(2.中國(guó)食品藥品檢定研究院國(guó)家藥物安全評(píng)價(jià)監(jiān)測(cè)中心,北京 100176)
致癌實(shí)驗(yàn)是創(chuàng)新藥物安全性評(píng)價(jià)和上市風(fēng)險(xiǎn)控制的重要組成部分。傳統(tǒng)的評(píng)價(jià)方法是采用兩年期大、小鼠致癌性實(shí)驗(yàn)。1997年,人用藥品注冊(cè)技術(shù)規(guī)定國(guó)際協(xié)調(diào)會(huì)(ICH)會(huì)議提出了致癌實(shí)驗(yàn)新框架(ICH S1B),要求在一項(xiàng)長(zhǎng)期嚙齒類動(dòng)物實(shí)驗(yàn)的基礎(chǔ)上,可選擇轉(zhuǎn)基因致癌模型進(jìn)行致癌實(shí)驗(yàn)。目前這種新的評(píng)價(jià)方法已獲得美國(guó)、歐盟、日本等管理部門的認(rèn)可[1-4],我國(guó)也在2010年提出了建議使用替代方法的致癌性實(shí)驗(yàn)指導(dǎo)原則[5],常用的小鼠模型為日本實(shí)驗(yàn)動(dòng)物中心研究所建立的Tg.rasH2[6-10]。通過代理機(jī)構(gòu)從日本購(gòu)買該模型,存在價(jià)格高、供應(yīng)周期長(zhǎng)等弊端,間接增加藥物研發(fā)成本,延緩國(guó)內(nèi)創(chuàng)新藥物發(fā)展。本單位從2004年開始籌備建立該致癌模型,2010年基本構(gòu)建成功[11]后陸續(xù)開展背景數(shù)據(jù)分析[12-17]、保種技術(shù)研究[18]等多項(xiàng)工作,并于2013年獲得國(guó)家發(fā)明專利(專利號(hào):ZL 2008 1 0101666.0.)[19]。構(gòu)建該模型不僅用于基礎(chǔ)研究,而且擬在中國(guó)建立具有自主知識(shí)產(chǎn)權(quán)的,符合ICH的藥物致癌實(shí)驗(yàn)替代方法,推動(dòng)我國(guó)該領(lǐng)域替代方法的建立。
本單位Tg.C57-rasV1.0和日本Tg.rasH2小鼠模型均采用傳統(tǒng)的轉(zhuǎn)基因方法獲得,因而模型的遺傳學(xué)特性是比較關(guān)注的問題[20]。其次,鑒于實(shí)驗(yàn)動(dòng)物的生物學(xué)特性是藥物臨床前長(zhǎng)期毒性研究中必須觀測(cè)的項(xiàng)目,且基因改造及繁育飼養(yǎng)條件的不一致,有必要將兩種模型的相關(guān)生物學(xué)特征進(jìn)行測(cè)定分析[21-23]。由于自發(fā)腫瘤是該轉(zhuǎn)基因動(dòng)物模型應(yīng)有的特性,但是自發(fā)腫瘤過高,必然會(huì)影響其應(yīng)用。在實(shí)際安全性評(píng)價(jià)中,使用的是ras轉(zhuǎn)基因動(dòng)物與BALB/c雜交F1代,以降低自發(fā)腫瘤的背景值,生物學(xué)特性比較使用的是自主建立的Tg.C57-ras轉(zhuǎn)基因小鼠與BALB/c小鼠的雜交 1 代小鼠(簡(jiǎn)稱CB6F1-NIFDC)與日本Tg.rasH2轉(zhuǎn)基因小鼠模型雜交 1 代小鼠(簡(jiǎn)稱CB6F1-CIEA),為國(guó)內(nèi)藥物安全評(píng)價(jià)機(jī)構(gòu)提供多元化的選擇,以期推動(dòng)我國(guó)自建致癌性小鼠模型的應(yīng)用。
1 兩種模型遺傳學(xué)特性比較
1.1 模型的構(gòu)建方式
日本科學(xué)家首先通過轉(zhuǎn)基因方法將人源的c-Ha-ras基因?qū)胄∈笈咛ブ校⒘藃as基因的小鼠模型,命名為Tg.rasH2。該轉(zhuǎn)入基因是通過將膀胱癌和黑色素瘤病人激活突變的c-Ha-ras進(jìn)行拼接操作得到,僅最后一個(gè)內(nèi)含子有突變,有自己的啟動(dòng)子和增強(qiáng)子(圖1A)。另外,日本模型是在DBA/2和C57BL/6小鼠品系的雜交胚中進(jìn)行轉(zhuǎn)基因操作,產(chǎn)生并獲得8只首建鼠,其中2只存活。初步比較后,選擇其中2系作為模型的首建鼠,并將首建鼠與C57BL/6J品系進(jìn)行超過10代的回交,使其與C57BL/6J回交純度達(dá)到99.5%后開始大規(guī)模生產(chǎn),目前使用的為C57BL/6J品系的Tg.rasH2小鼠模型[24-26]。
本單位從2004年開始,參照日本模型設(shè)計(jì)思路,構(gòu)建了人源的c-Ha-ras的轉(zhuǎn)基因小鼠模型,命名為Tg.C57-rasV1.0。該模型同樣使用轉(zhuǎn)基因方法構(gòu)建,PCR方法克隆人的原癌基因c-Ha-ras,含有4個(gè)外顯子和該基因本身的啟動(dòng)子、調(diào)控序列和polyA信號(hào)序列[11](圖1B)。轉(zhuǎn)入基因在原核注射后的整合率達(dá)到1.6%,共得到6只首建鼠,其中3只成活,篩選后選擇3系保種繁殖。目前,已獲得10余代仍能穩(wěn)定遺傳和表達(dá)c-Ha-ras基因的致癌性小鼠[12]。

圖1 我國(guó)Tg.C57-ras V1.0與日本Tg.rasH2致癌性轉(zhuǎn)基因小鼠的轉(zhuǎn)基因結(jié)構(gòu)策略注:MM: 來(lái)自于黑素瘤患者的人源c-Ha-ras基因片段(Bam HI)該患者的61號(hào)密碼子發(fā)生了突變。BB: 來(lái)自于膀胱癌患者的的人源c-Ha -ras基因片段(Bam HI)片段,患者的12號(hào)密碼子發(fā)生了突變;MB:MM和BB沒有突變的重組基因,MB基因被用來(lái)構(gòu)建Tg.rasH2小鼠模型;★指在基因最后的內(nèi)含子突變的位置Fig.1 Transgenic structure strategy of Tg.C57-ras V1.0 and Japanese Tg.rasH2 transgenic miceNote: MM: a human c-HRAS gene (Bam HI fragment) derived from a patient with melanoma carrying the mutation at codon 61. BB:a human c-HRAS Bam Hl gene fragment derived from a patient with urinary bladder carcinoma carrying the mutation at codon 12.MB: the recombinant of MM and BB with no mutation at codons 12 and 61. The MB gene was introduced to create the original rasH2 mouse.
1.2 拷貝數(shù)分析
c-Ha-ras基因的拷貝數(shù)對(duì)于模型的遺傳穩(wěn)定性和致癌性評(píng)價(jià)功能非常重要[27]。根據(jù)Suemizu[28]報(bào)道,日本模型的拷貝數(shù)通過PCR方法一共轉(zhuǎn)入3個(gè)拷貝基因,并在15號(hào)染色體E3區(qū)以從頭到尾的方式排列,且有自己的啟動(dòng)子和增強(qiáng)子。
根據(jù)文獻(xiàn)報(bào)道采用Real-time PCR方法,可以更加迅速的鑒定拷貝數(shù)。根據(jù)標(biāo)準(zhǔn)曲線方程,通過各樣本c-Ha-ras和GAPDH的Ct值分別計(jì)算出各自的初始模板數(shù)(NRAS和NCAPDH),經(jīng)計(jì)算,目前保留的轉(zhuǎn)基因3系小鼠的拷貝數(shù)為4[14]。
1.3 mRNA表達(dá)水平及蛋白表達(dá)水平
1.3.1兩種模型不同組織轉(zhuǎn)入基因,mRNA水平結(jié)果:轉(zhuǎn)基因沉默,即不能正常表達(dá)或者表達(dá)量過低,是建立轉(zhuǎn)基因模型過程中常見的現(xiàn)象,也是轉(zhuǎn)基因模型建立失敗的重要原因[29-30]。分別選取8周齡小鼠3只,解剖其主要目的臟器,結(jié)果表明轉(zhuǎn)入基因在兩種小鼠的心、肝、脾、肺、腎以及前胃6種組織器官中具有相似的mRNA表達(dá)譜,均在肺臟表達(dá)最高,而在心臟表達(dá)最低;肺臟中,日本模型表達(dá)量高于自建模型,其他臟器表達(dá)量相當(dāng)(圖2A)[12]。
1.3.2兩種模型肺組織轉(zhuǎn)入基因,蛋白表達(dá)水平結(jié)果:mRNA水平兩模型轉(zhuǎn)入基因在肺組織中表達(dá)最高,因此選取肺組織檢測(cè)蛋白表達(dá)并進(jìn)行灰度掃描,內(nèi)參GAPDH校正,衡量蛋白表達(dá)水平。CB6F1-CIEA小鼠的灰度值(96.4±9.9)、CB6F1-NIFDC小鼠的灰度值(63.4±1.5),野生型小鼠的灰度值(25.7±1.8)。兩種轉(zhuǎn)基因小鼠與野生型的表達(dá)差異均有統(tǒng)計(jì)學(xué)意義(P<0.01)。蛋白結(jié)果及灰度統(tǒng)計(jì)結(jié)果見圖2B。
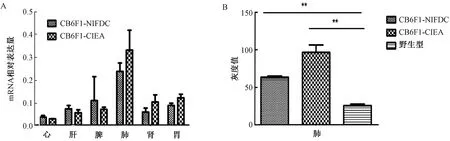
圖2 轉(zhuǎn)入基因在兩種小鼠組織器官中的mRNA及蛋白水平表達(dá)注:**P<0.01:兩種轉(zhuǎn)基因小鼠與野生型小鼠的表達(dá)差異極顯著Fig.2 mRNA and protein expressions of the transferred gene in 2 kinds of mouse tissues and organsNote:**P<0.01, the expression difference between the two transgenic mice and the wild type was very significant
2 兩種模型的生物學(xué)特性比較
2.1 模型生長(zhǎng)曲線
鼠的生長(zhǎng)發(fā)育性能是動(dòng)物飼養(yǎng)管理和健康狀況的重要指標(biāo)。本研究中測(cè)定的是致癌實(shí)驗(yàn)中使用的是c-Ha-ras陽(yáng)性小鼠與BALB/c小鼠的雜交一代。Yamamoto等[31]測(cè)定了日本模型55周的生長(zhǎng)曲線(圖3A),基因陽(yáng)性的雄性和雌性鼠分別是野生型小鼠體質(zhì)量的80%~90%。另外,Nambiar等[32]報(bào)道輝瑞公司的研究數(shù)據(jù)表明,6個(gè)月的致癌實(shí)驗(yàn)中,Tg.rasH2雄性小鼠和雌性小鼠的平均初始體質(zhì)量分別為24.7 g和19.6 g,最終體質(zhì)量分別為28.4~34.5 g和22.9~25.1 g。
關(guān)于自建模型,本實(shí)驗(yàn)室也選取雌雄小鼠各20只,測(cè)定其離乳后的體質(zhì)量共24周,具體生長(zhǎng)曲線見圖3B所示。比較兩種模型的生長(zhǎng)曲線數(shù)據(jù),生長(zhǎng)趨勢(shì)一致。另致癌實(shí)驗(yàn)所用的雄雌鼠的平均初始體質(zhì)量分別為24.2 g和18.3 g,與日本小鼠差別不大。

圖3 兩種模型的生長(zhǎng)曲線注:A,CB6F1-CIEA小鼠及同窩野生鼠的生長(zhǎng)曲線,參考Yamamoto等[31];B,CB6F1-NIFDC小鼠及同窩野生輸?shù)纳L(zhǎng)曲線Fig.3 Growth curves of two modelsNote:A,The growth curve of CB6F1-CIEA mice and wild mice in the same nest, modified from Yamamoto et al.[31];B,Growth curve of CB6F1-NIFDC mice and wild mice in the same nest
2.2 血液生理數(shù)據(jù)
根據(jù)Sehata等[33]數(shù)據(jù)顯示日本模型心、肺、肝、脾、腎、腎上腺、睪丸、卵巢等主要臟器臟體比、血液化學(xué)和血液學(xué)與陰性小鼠沒有顯著差異。對(duì)于自建模型,本單位也多次測(cè)定及比較臟器質(zhì)量及血液生理生化指標(biāo),包括22項(xiàng)血液生理學(xué)指標(biāo)及12 項(xiàng)血液生化指標(biāo)。其中,雌性陽(yáng)性鼠和陰性鼠的嗜中性粒細(xì)胞(NEUT)及嗜中性粒細(xì)胞百分比(NEUT%),差異顯著(P<0.05),血小板壓積(PCT)差異極顯著(P<0.01),其余19 項(xiàng)無(wú)顯著性差異(P>0.05)。雌性陽(yáng)性鼠和陰性鼠的12項(xiàng)生化指標(biāo)均差異不顯著(P>0.05);雄性中,丙氨酸氨基轉(zhuǎn)移酶(ALT)和甘油三酯(TG)差異顯著(P< 0.05),其余10項(xiàng)差異不顯著(P>0.05),具體見前期研究[13-15]。
3 討論與展望
近幾年來(lái),隨著新藥研發(fā)項(xiàng)目增加,國(guó)內(nèi)藥物安全評(píng)價(jià)機(jī)構(gòu)對(duì)致癌性小鼠模型的需求不斷增加[34-35]。我國(guó)擁有自主知識(shí)產(chǎn)權(quán)的致癌動(dòng)物模型非常必要。為推動(dòng)國(guó)內(nèi)基于轉(zhuǎn)基因小鼠模型的致癌實(shí)驗(yàn)平臺(tái)加快發(fā)展,自建Tg.C57-rasV1.0小鼠模型與日本Tg.rasH2小鼠進(jìn)行了比較。遺傳學(xué)特性:自建與日本模型均是通過轉(zhuǎn)基因方法構(gòu)建,拷貝數(shù)分別是4和3,轉(zhuǎn)入基因c-Ha-ras在不同臟器的mRNA水平表達(dá)趨勢(shì)相似,肺組織中蛋白水平表達(dá)無(wú)統(tǒng)計(jì)學(xué)差異。生物學(xué)特性:兩種模型的生長(zhǎng)曲線,特別是致癌實(shí)驗(yàn)所需周齡的小鼠平均初始體質(zhì)量差別不大。另外,兩種模型和同窩轉(zhuǎn)基因陰性動(dòng)物相比較,其血液生理、臟器和臟器系數(shù)相差不大,這有利于模型將來(lái)在臨床前安全評(píng)價(jià)中應(yīng)用。雖然近幾年來(lái),國(guó)內(nèi)市場(chǎng)采用我國(guó)致癌性轉(zhuǎn)基因模型呈上升趨勢(shì),但實(shí)例較日本的少很多,背景數(shù)據(jù)不及日本模型豐富。
通過進(jìn)一步文獻(xiàn)查閱,Tamaoki[36]報(bào)道日本模型構(gòu)建時(shí)采用的是DBA/2和C57BL/6小鼠品系的雜交胚,如果回交不充分,會(huì)導(dǎo)致模型遺傳背景不純,從而降低了實(shí)驗(yàn)數(shù)據(jù)的質(zhì)量。本單位自建模型為C57BL/6的背景,小鼠種群不經(jīng)過回交,用于藥物致癌實(shí)驗(yàn)的動(dòng)物遺傳背景一致,減少了個(gè)體差異對(duì)結(jié)果的影響,是一種有實(shí)際意義的改進(jìn)。
眾所周知,采用轉(zhuǎn)基因方式構(gòu)建的小鼠模型存在一定程度的不足,主要表現(xiàn)在其外源基因在模型中遺傳和表達(dá)的穩(wěn)定性較差,需要通過不間斷遺傳監(jiān)測(cè)來(lái)維持種群遺傳穩(wěn)定性。目前,基于干細(xì)胞打靶技術(shù)在編輯復(fù)雜基因領(lǐng)域已經(jīng)獲得長(zhǎng)足進(jìn)步[37]。本單位已獲得基于干細(xì)胞打靶技術(shù)建立的致癌性模型,相較于日本Tg.rasH2小鼠模型,吸取其優(yōu)勢(shì),例如:與日本模型插入基因序列c-Ha-ras的拷貝數(shù)、基因位點(diǎn)以及小鼠的遺傳背景均一致。按照現(xiàn)代遺傳學(xué)認(rèn)知,可以推測(cè)新構(gòu)建的模型具有與日本Tg.rasH2小鼠相似或者相同的表型。本研究采用定點(diǎn)插入技術(shù),摒棄了轉(zhuǎn)基因動(dòng)物模型遺傳穩(wěn)定性差,個(gè)體差異較大的缺點(diǎn),新建的模型稱為致癌性模型KI.C57-rasV2.0,是原有Tg.C57-rasV1.0致癌性小鼠的升級(jí)版模型。新的致癌模型已經(jīng)由本單位安評(píng)中心及國(guó)內(nèi)多家安評(píng)中心展開聯(lián)合驗(yàn)證,實(shí)現(xiàn)有動(dòng)物模型可用、有技術(shù)標(biāo)準(zhǔn)可查、有關(guān)鍵技術(shù)可用及有背景數(shù)據(jù)可考的新局面[38],并滿足我國(guó)藥物安全性評(píng)價(jià)、藥品檢定及新藥創(chuàng)制中對(duì)此類遺傳修飾模型的需要。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
童話王國(guó)·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
