MAOA在誘導前列腺癌神經內分泌分化中的作用*
2021-11-04 10:52:56謝清華張彩勤譚鄧旭師長宏趙菊梅
實驗動物科學 2021年4期
稅 雪 許 榮 謝清華 張彩勤 譚鄧旭 師長宏 趙菊梅
(1. 延安大學醫學院,延安 716000)(2. 空軍軍醫大學實驗動物中心,西安 710032)
前列腺癌(prostate cancer,PCa)是一種雄激素驅動的疾病,它依賴于配體介導的雄激素受體(androgen receptor,AR)激活從而促進疾病進展[1]。盡管雄激素剝奪可以有效控制腫瘤生長,但大多數患者最終會發展成為去勢抵抗性前列腺癌(castration-resistant prostate cancer,CRPC)[2]。Watson等[3]、Parker等[4]以及Hussain等[5]研究表明,大多數CRPC仍依賴于AR信號通路,一些新型的AR通路抑制劑(androgen receptor inhibitor,ARPI),如:恩雜魯胺(enzalutamide,ENZ)和阿比特龍作為治療藥物應用于晚期前列腺癌患者以延長生存期。Nadal等[6]和Bishop等[7]報道,ARPI會誘導神經內分泌性前列腺癌(neuroendocrine prostate cancer,NEPC),出現特征性多器官轉移,導致難以治愈。目前,ARPI引起PCa神經內分泌分化(neuroendocrine transdifferentiation,NED)的確切分子機制尚未完全闡明,主要原因是缺乏模擬臨床特征的異質性轉化模型。因此,構建NEPC轉化模型已成為PCa防治研究亟待解決的重大問題。
單胺氧化酶A(monoamine oxidase A,MAOA)是一種位于線粒體上的氧化還原酶,通過氧化脫氨催化膳食胺和單胺神經遞質降解,產生副產物過氧化氫,即活性氧(reactive oxygen species,ROS)的主要來源[8-10]。Trachootham等[11]報道,ROS可使細胞發生氧化損傷,進而導致腫瘤的發生和發展。Wu等[12]、Liao等[13]及Li等[14]研究證實MAOA表達水平的升高會促進PCa細胞發生上皮—間質轉化,誘導癌細胞干性等。目前,MAOA對ENZ誘導NED的影響還未見報道。因此,本研究擬通過ENZ誘導建立NEPC細胞模型和動物模型,探索MAOA在該誘導過程中發揮的作用及潛在機制,旨在為揭示 NEPC的發生機制提供良好的模型,為PCa的治療提供新靶點。
1 材料和方法
1.1 材料
1.1.1實驗動物:本研究選用15只6~7周齡SPF級雄性BALB/c裸鼠,體質量22~25 g,購自常州卡文斯實驗動物有限公司,實驗動物生產許可證號:SCXK(蘇)2016-0010,在空軍軍醫大學實驗動物中心SPF環境中飼養和繁殖,實驗動物使用許可證號:SYXK(陜)2018-001。動物實驗通過空軍軍醫大學實驗動物福利及倫理委員會批準,動物倫理證書編號:20190207。
1.1.2細胞:人前列腺癌細胞C4-2和22RV1購自國家實驗細胞資源共享平臺。細胞培養條件為RPMI-1640培養基(10%胎牛血清+1%青鏈霉素),37 ℃、5% CO2孵箱中培養。
1.1.3試劑:RPMI-1640培養基和0.05%胰酶購自HyClone;FBS購自浙江天杭生物科技股份有限公司;免疫組化染色試劑盒購自北京中杉金橋生物技術有限公司;Western blot制膠試劑盒購自上海碧云天生物技術有限公司;MAO-Glo試劑盒購自Promega;熒光定量PCR等分子生物相關試劑購自寶生物工程(大連)有限公司。抗體:神經內分泌標志物抗體(CGA和SYP)和前列腺癌特異性抗體(PSA和AR)均購自Abcam;神經內分泌標志物抗體ENO2抗體購自ProMab;Actin抗體和二抗購自康為世紀生物科技股份有限公司。藥品:ENZ購自Selleck;MAOA特異性抑制劑氯吉靈(clorgyline,CLG)購自MCE;異氟烷購自深圳市瑞沃德生命科技有限公司。
1.1.4儀器:酶標儀購自Biotek Take;基因擴增儀購自Eppendorf;Real-time PCR購自StepOne Plus、Thermo Fisher等。
1.2 方法
1.2.1RNA提取和Real-time PCR:使用TRIzol破碎和溶解細胞, 收集總RNA。使用逆轉錄酶逆轉錄獲得cDNA。使用 Step One Plus 實時 PCR 檢測系統(Thermo Fisher ABI),PrimeScriptTMRT 試劑盒和 TB Green? Fast qPCR Mix(Takara)進行Real-time PCR實驗,95 ℃聚合酶激活3 min,然后進行40個循環的兩步法PCR(95 ℃、30 s和60 ℃、40 s),最終PCR反應結束后直接進行熔解曲線分析。使用2-ΔΔCT方法計算mRNA的相對表達水平,β-actin作為內參。
1.2.2免疫印跡分析:細胞或破碎后瘤體組織,使用RIPA裂解液,置于冰上裂解30 min,4 ℃、12 000 r/min離心10 min收集上清液,使用BCA法進行蛋白定量。蛋白在10% SDS-PAGE凝膠上分離,然后轉移到PVDF膜上并在室溫下用5%脫脂奶粉封閉2 h。特異性一抗4 ℃孵育過夜,酶標二抗室溫2 h后,通過ECL試劑盒觀察細胞膜上的信號。β-actin作為內參。
1.2.3MAOA活性分析:熒光素酶裂解緩沖液裂解1×104個細胞,MAO-Glo試劑盒分析細胞裂解液的MAOA活性,MAO-Glo檢測系統記錄熒光信號值[15]。
1.2.4MTT法檢測各細胞株耐藥指數:將2×103/孔接種于96孔板中,待細胞貼壁后,使用100 μL含有不同濃度的ENZ培養基培養細胞96 h后,向每孔加入培養基/ CCK8混合物(9∶1),繼續孵育2.5 h,用酶標儀檢測,于450 nm處讀取吸光度值。
1.2.5體內實驗:所有小鼠均接受手術去勢,于一周后右側皮下植入1 × 106個22RV1細胞,建立異種移植模型。腫瘤體積=1/2 ×(長軸 × 寬軸)2。當腫瘤體積達到100 mm3,將小鼠隨機分為三個治療組,A組:對照組(22RV1)、B組:10 mg/kg的ENZ每日灌胃(22RV1+ENZ)及C組:10 mg/kg ENZ+10 mg/kg 的CLG,每日腹腔注射(22RV1+ENZ+CLG),n=15。治療兩周后視為實驗終點,對所有荷瘤小鼠CO2窒息處死,并取組織用于免疫組化分析。
1.2.6免疫組化:小鼠腫瘤組織經4%多聚甲醛固定,石蠟包埋,切成4 μm切片,通過間接免疫過氧化物酶技術進行免疫組織化學染色,光學顯微鏡下觀察出現背景清晰的棕色或棕黃色顆粒判為陽性染色。
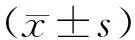
2 結果
2.1 ENZ誘導的NEPC模型建立與表征
為了選擇適合NED研究的PCa細胞系,對PCa細胞系C4-2和22RV1對AR抑制劑的敏感性進行了評估。MTT檢測結果顯示,C4-2細胞對ENZ敏感,22RV1細胞對ENZ不敏感(圖1A),因此選擇C4-2細胞進行 10~20 μmol/L ENZ連續誘導培養4個月,以期獲得神經內分泌樣的細胞。結果如圖1B所示,與親本相比,ENZ處理的C4-2細胞能在更高濃度的藥物中增殖,耐受濃度從34.64 μmol/L升高至82.17 μmol/L。同時,細胞形態發生明顯改變,呈現突觸增多、胞體聚集等神經樣細胞表型(圖1C)。因此,將ENZ耐藥細胞株C4-2命名為C4-2ENZR。Western blot結果表明,與親本細胞相比,C4-2ENZR細胞中NE標志物嗜鉻粒蛋白A(CGA)、突觸素(SYP)和神經元特異性烯醇化酶(ENO2)的蛋白表達增加,腺癌標志物雄激素受體(AR)表達降低(圖1D)。這表明該細胞模型為NEPC模型[7]。
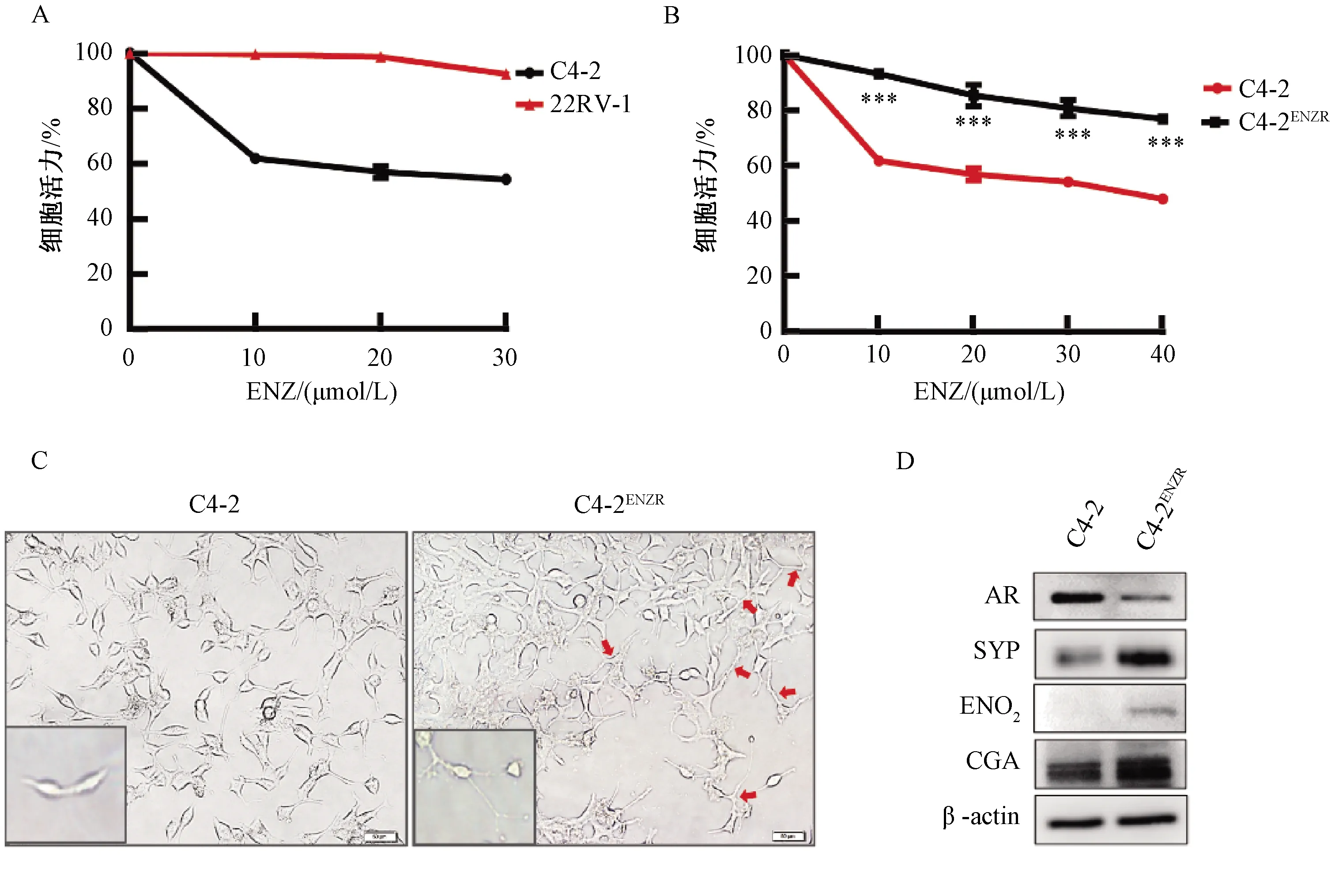
圖1 NEPC模型的建立注:***P<0.001Fig.1 Establishment of NEPC modelNote:***P<0.001
2.2 NEPC細胞模型中MAOA表達升高
根據Wu等[12]報道,MAOA的表達與PCa的惡性程度相關,因此使用Oncomine數據庫分析MAOA在兩個不同臨床隊列的表達。結果顯示,與正常前列腺組織相比,人類PCa組織中MAOA的表達更高,且與Gleason評分呈正相關(圖2A)。進一步通過RT-PCR和Western blot檢測發現,與親本C4-2細胞相比,C4-2ENZR細胞中MAOA的mRNA和蛋白的表達顯著升高(圖2B、圖2C),從而驗證了上述生物信息學結果,即:MAOA可能在ENZ誘導的NED中起關鍵作用。
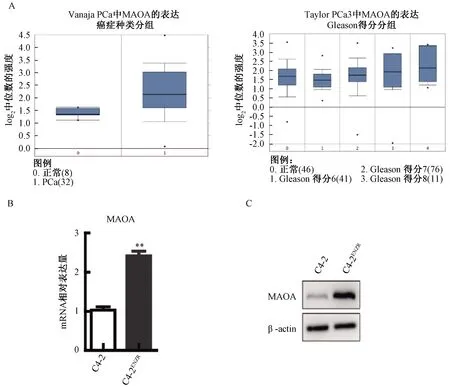
圖2 MAOA的表達與ENZ誘導的神經內分泌分化有關注:*P<0.05,**P<0.01Fig.2 The expression of MAOA is associated with the development of NED induced by ENZNote:*P<0.05, **P<0.01
2.3 MAOA抑制劑CLG可以延緩ENZ引起的NED發生
CLG作為一種選擇性抑制劑,可以顯著降低MAOA酶的活性(圖3A)。對C4-2細胞進行ENZ給藥時,同時添加1 μmol/L的CLG聯合處理細胞2個月后檢測發現,ENZ與CLG聯合使用可以減輕ENZ所引起的神經內分泌樣細胞形態學改變,并抑制神經內分泌標志物NCAM1和ENO2表達(圖3B、圖3C)。這表明MAOA是促進NED和維持神經內分泌細胞特性的關鍵因素,以MAOA為靶點的抑制劑CLG可以延緩ENZ引起的NED的發生。為了進一步證實體外細胞系研究的結果,建立了22RV1裸鼠異種移植模型,結果顯示ENZ確實可以誘導NEPC體內模型形成,該模型表現出超高水平的神經內分泌標志物陽性表達。更重要的是,與ENZ單藥治療的腫瘤相比,ENZ+CLG聯合使用能夠降低22RV1小鼠腫瘤中的神經內分泌標志物表達(圖3D)。
圖3 MAOA抑制劑CLG可以顯著延緩體內和體外的NED注:***P<0.001Fig.3 Clorgyline, inhibitors of MAOA, can significantly delay NED in vivo and in vitroNote:***P<0.001
2.4 靶向MAOA抑制缺氧信號以克服NED
缺氧是多種實體瘤的常見情況,Lu等[16]研究發現,缺氧可以促進ROS的產生,從而促進疾病進展。Shih等[10]證實,MAOA介導的酶促反應產生大量作為ROS主要來源的過氧化氫。因此,流式分析C4-2、C4-2+ENZ 和C4-2+ENZ+CLG細胞的ROS水平。如圖4所示,MAOA刺激ROS的產生,由26.5%增至48.8%;CLG延緩ROS的升高,使其降為40.3%。以上結果提示,MAOA可以通過改變C4-2細胞中的缺氧信號來克服ENZ引起的NED。
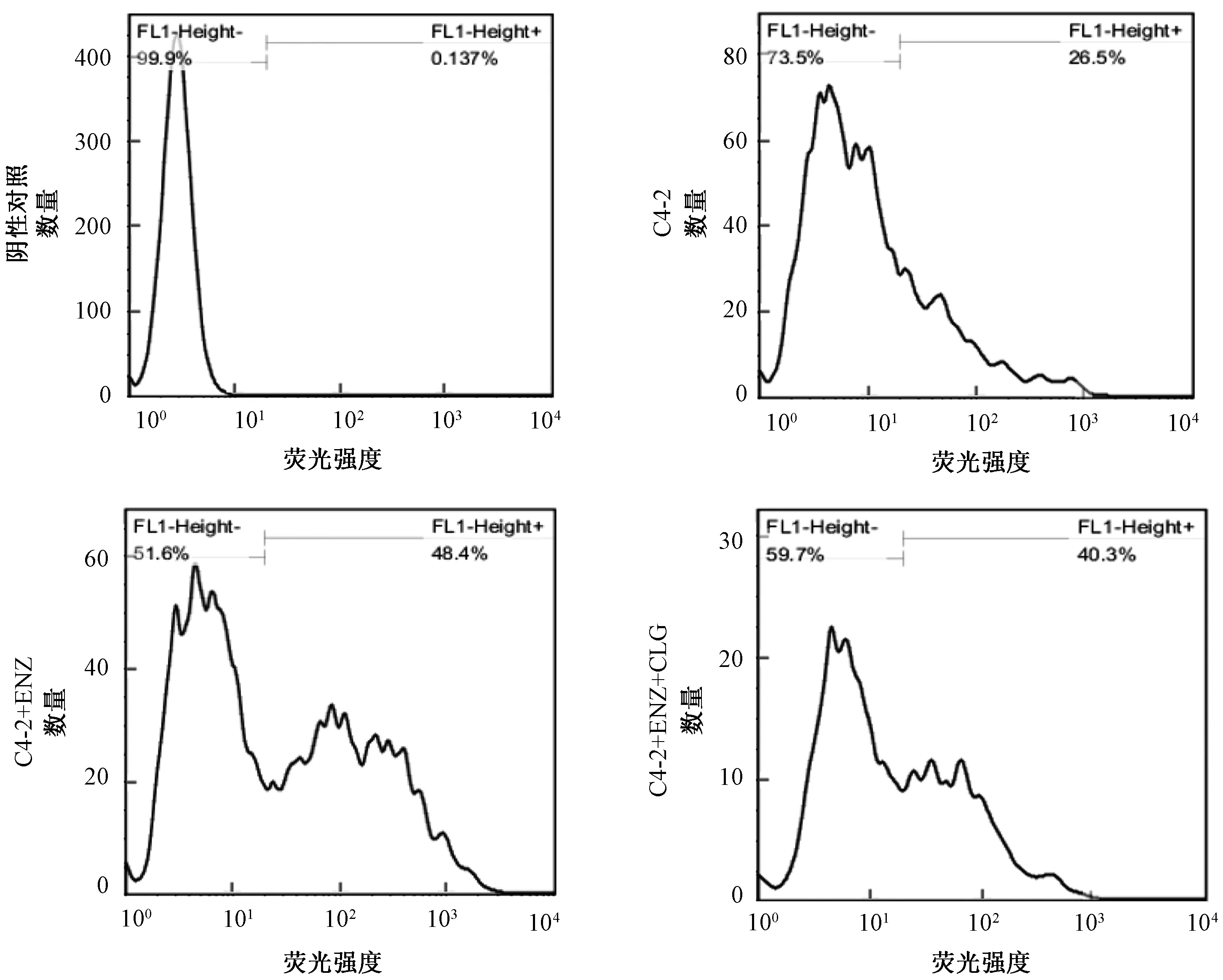
圖4 MAOA 激活缺氧信號以促進 NEDFig.4 MAOA triggers hypoxia signaling to promote the NED
3 討論
PCa是老年男性患者死亡的主要原因之一,NEPC屬于PCa的終末階段,侵襲性強且預后極差[17]。目前NEPC的研究進展緩慢,主要是缺乏模擬臨床特征的異質性轉化模型[18]。因此, NEPC模型的構建是本研究的關鍵。通過ENZ構建具有神經內分泌特征的NEPC體外模型C4-2ENZR,該細胞模型高表達SYP,CGA和ENO2,低表達AR,為典型的NEPC細胞。體內模型選取22RV1細胞系,相比于C4-2細胞,22RV1細胞在裸鼠體內具有較好的成瘤率,并且對ENZ表現出耐藥性,更接近于NEPC特征[7,19]。本研究將22RV1移植入去勢小鼠體內建立異種移植模型,進一步用ENZ誘導并經組化染色證實,神經內分泌標志物SYP表達強陽性,提示該模型具有NEPC特征。以上模型的成功建立為后續分子機制研究提供了堅實基礎。
目前,ENZ引起NED的具體機制仍不完全清楚。Luo等[20]發現LncRNA-p21調節EZH2/STAT3信號通路改變ENZ誘導的PCa NED。Bishop等[7]也發現主神經轉錄因子(BRN2)是AR抑制后NEPC形成的主要驅動基因,但MAOA與NEPC的關系尚未見報道。此外,目前除化療外,尚無針對NEPC患者的有效治療方法。一些靶向抑制劑,如EZH2抑制劑GSK126[21],AR降解增強劑ASC-J934[21-23]等被證明能起到一定的治療效果,但多處于臨床前研究或臨床試驗階段,距離新藥上市時間較長。因此,重新利用現有藥物是解決這些問題的有效方法。
現有的抗抑郁藥物CLG可有效抑制MAOA的活性,Wu等[15]發現CLG可以擾亂導致癌細胞侵襲和增殖的信號通路,進而抑制PCa進展。同樣,本研究也證實,在晚期PCa中,靶向MAOA可能是抑制PCa進展的潛在治療策略,結果表明(1)ENZ處理后,MAOA的活性和表達量增加,提示MAOA可能在ENZ誘導的NED中起關鍵作用;(2)MAOA升高可以促進神經神經內分泌標志物的表達,并且可能通過改變下游缺氧信號通路實現的。最重要的是,證明CLG可以有效延緩ENZ引起的NED。
綜上所述,本研究證實,MAOA是促進NED和維持神經內分泌細胞特性的關鍵因素,現有的MAOA抑制劑CLG可以延緩NEPC發生。將CLG與ENZ聯合使用,有可能成為PCa患者的新型治療方案。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
家庭醫學(下半月)(2020年3期)2020-05-30 12:42:02
家庭醫學(下半月)(2020年3期)2020-05-30 12:42:00
家庭醫學(下半月)(2020年3期)2020-05-30 12:42:00
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年7期)2019-01-06 09:27:34
光學精密工程(2016年6期)2016-11-07 09:07:19
