GABA抑制腹主動脈瘤小鼠未成熟樹突狀細胞的遷移機制
2021-10-22 10:20:48楊穎羅暉馮杰曾韡
川北醫學院學報 2021年9期
關鍵詞:小鼠
楊穎,羅暉,馮杰,曾韡
(1.川北醫學院附屬醫院心內科·川北醫學院附屬醫院心血管疾病研究室;2.南充市中心醫院心胸外科,四川 南充 637000)
腹主動脈瘤(abdominal aortic aneurysm,AAA)及其并發的動脈破裂是一類致命的血管性疾病,動脈粥樣硬化(atherosclerosis,AS)是其常見的病因[1]。AAA的主要病理改變為炎癥細胞浸潤,血管平滑肌細胞凋亡,彈力纖維和膠原纖維斷裂,細胞外基質(extracellular matrix,ECM)降解,從而導致動脈管腔擴張或膨出甚至發生破裂,造成嚴重出血,危及生命[2]。因此,如何抑制炎癥細胞的活化,阻止ECM降解是預防AAA的主要措施,也是目前尚未解決的臨床難題。γ-氨基丁酸(γ-aminobutyric acid,GABA)A 型受體是配體門控氯離子通道超家族的成員,通常是由 α,β,γ,δ,ε,π,θ,ρ 8 類亞基組成的異源五聚體。這些亞基又組成眾多功能結構特異的GABA-A受體。GABA激動GABA-A受體后對腦內抑制性神經傳遞進行調控[3]。近年來,研究[4]發現,GABA-A受體同樣表達于外周樹突狀細胞、巨噬細胞、T淋巴細胞等細胞膜上,抑制炎癥免疫細胞的活性,延緩某些自身免疫性疾病的進展。本小組既往研究[5]發現,在AS模型中,激活外周血單核-巨噬細胞膜表面的GABA-A受體可改善巨噬細胞內失衡的膽固醇轉運,抑制巨噬細胞分泌TNF-α,上調調節性T淋巴細胞的比例,減輕載脂蛋白E缺乏(apoE-/-)小鼠主動脈粥樣硬化斑塊的形成。該結果提示,AS 與AAA同為血管炎癥性疾病,發病機制類似,炎癥免疫反應貫穿始終[6]。本研究旨在探討GABA是否能夠通過其受體抑制腹主動脈瘤小鼠未成熟樹突狀細胞(immature dendritic cells iDCs)的遷移機制。
1 材料與方法
1.1 實驗材料
C57BL/6J雄性小鼠(購于川北醫學院實驗動物中心),鼠齡6周,體重18~20 g。日夜節律變化條件為12 h光照和12 h黑暗,食物和飲水按需供給。實驗動物的使用都嚴格按照川北醫學院實驗動物中心的動物倫理學條例執行。GABA購于美國Sigma 公司。
1.2 方法
1.2.1 動物處理 彈性蛋白酶(PPE)構建小鼠腹主動脈瘤動脈模型[7-8]:將小鼠麻醉后,打開腹腔游離出腎動脈以下髂動脈分叉以上腹主動脈段,依次結扎腹主動脈分支,先行阻斷腹主動脈近端,記錄阻斷時間,接著阻斷腹主動脈遠端,在灌注段腹主動脈穿孔,接入PPE管,結扎固定,檢驗灌注段腹主動脈密閉性,PPE灌注濃度采用4.5 U/mL,維持灌注壓100 mmHg,灌注時間為5 min,灌注完畢后拔出PPE管,修復穿刺口,檢查若無出血,逐層關腹。手術時間40~60 min,建模周期14 d。腹主動脈擴張率(%)=(術后4周腹主動脈直徑-灌注前腹主動脈直徑)/灌注前腹主動脈直徑×100%,以術后14 d腹主動脈擴張率50%以上作為診斷腹主動脈瘤的標準。病理組織學檢測提示,細胞外基質降解,平滑肌細胞減少或消失,炎癥細胞浸潤[9]。
1.2.2 細胞培養 獲取C57BL/6J腹主動脈瘤小鼠骨髓來源的未成熟樹突狀細胞。將小鼠脫頸處死,在無菌條件下分離小鼠股骨。用PBS將股骨中骨髓細胞沖洗出來直至股骨泛白。收集沖洗液使用細胞篩過濾。將過濾后的沖洗液移入15 mL離心管中,1 000 rpm離心5 min。倒去上清液,加入適量的紅細胞裂解液輕輕吹打混勻,4℃裂解 1~2 min。加入10~20 mL PBS混勻,離心棄上清。將沉淀加入無血清培養基6 mL吹打混勻后移至6孔板中,每孔1 mL,放入37℃培養箱孵育3 h后小心倒掉上清液,每孔分別加入2 mL完全培養基及CSF和IL-4各10 μL,放入37℃培養箱中繼續培養。隔天換液并加入新的CSF和IL-4,第6天收集細胞液。從而獲得未成熟DCs細胞[10]。
1.2.3 細胞轉染 將收集的未成熟DCs細胞接種至6孔板中,每孔加入細胞液1 mL;用移液槍吸取0.35 μL siRNA加入200 μL 1640培養基;加入15 μL轉染試劑,震蕩混勻后室溫下靜置15 min;混合液加入細胞液中后放入37℃培養箱孵育6 h,每孔加入新鮮完全培養基2 mL。Gabara5 SiRNA GGUGCGAACAGACAUCUAUTT;AUAGAUGUCUGUUCGCACCTT(購于上海生工)。
1.2.4 Transwell遷移實驗觀察細胞遷移能力 轉染48 h后收集細胞離心,將細胞分為對照組(生理鹽水組)、GABA(200 nmol/L)組、Gabara5siRNA+GABA(200 nmol/L)組;上室中各加入細胞懸液100 μL,下室中加入500 μL完全培養基,放入37℃培養箱孵育2 h;PBS淋洗,4%多聚甲醛固定15 min,DAPI染色10 min,PBS洗滌3次;待干燥后熒光顯微鏡下觀察拍照,采用Image J軟件分析,計算細胞數。
1.2.5 熒光技術觀察細胞骨架 收集未成熟的DCs細胞,調整細胞濃度為106/mL,用移液槍抽吸細胞液1 mL接種于多聚賴氨酸處理過的載玻片上,放入37℃培養箱孵育48 h;經PBS清洗,4%多聚甲醛固定,0.25%Tritonx-100破膜,滴加稀釋好的鬼筆環肽至載玻片上,室溫避光孵育,PBS洗滌,待晾干后滴加抗熒光淬滅封片劑,熒光顯微鏡下觀察拍照顯示F-肌動蛋白染色(F-actin)。
1.2.6 蛋白免疫印跡法(Western Blot)檢測 GABAA受體α5亞基的表達 收集細胞轉染后iDCs,提取細胞蛋白,免疫組化BCA法測蛋白濃度,依次通過制膠、電泳、轉膜、染色、封閉、抗體孵育,最后化學發光、成像、分析。
1.3 統計學分析
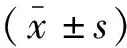
2 結果
2.1 未成熟樹突狀細胞(iDCs)的培養及純度
培養的第0天光學顯微鏡觀察顯示培養液中充滿大量圓形單核細胞;第2天觀察發現細胞貼壁生長且有集落生成,部分細胞開始伸出突起;第4天觀察發現集落數量增加,伸出突起的細胞數量增多;第6天觀察發現有大量細胞懸浮在培養液中,細胞周圍有長短不一的突起。因單核細胞來源的iDC表達CD11c,CD11c-FITC抗體檢測iDC純度。在第6天收集細胞液后加入CD11c-FITC抗體標記,利用流式細胞儀檢測細胞純度在80%左右。見圖1。

2.2 GABAA受體α5亞基SiRNA轉染情況
GABAA受體α5亞基SiRNA轉染率>90%。將iDCs分為空白組(生理鹽水組)、GABA組、轉染試劑組、NC-FAM組(NC組)、Gabara5siRNA組(SiRNA組),轉染48 h后收集細胞利用Western blot檢測GABA-A受體α5亞基的表達發現,GABA可以促進GABA-A受體α5亞基表達,差異無統計學意義(P>0.05),而SiRNA干擾了GABAA受體α5亞基的表達,差異有統計學意義(P<0.05)。見圖2。

2.3 GABA通過GABA-A受體抑制iDCs的遷移
將iDCs分為空白組(生理鹽水組)、GABA組、Gabara5siRNA組(SiRNA組),其中GABA組和SiRNA組分別加入200 nmol/L的GABA共孵育,利用transwell小室檢測細胞遷移能力,結果顯示,與對照組比較,GABA組iDCs的個數明顯減少,差異有統計學意義(P<0.05)。SiRNA組iDCs的個數較對照組無明顯變化,這些結果表明GABA對iDCs的遷移能力起負調節作用,并且可能是通過GABA-A受體α5亞基進行的。見圖3。
2.4 GABA/GABA-A受體通路抑制iDCs偽足的形成
將GABA加入iDCs中共孵育1 h,激光共聚焦顯微鏡發現,加入GABA后iDCs的偽足生成明顯減少。然后利用SiRNA敲低iDCs表面的GABA-A受體α5亞基后與GABA共孵育1 h,F-actin熒光染色結果顯示iDCs周圍有較多的偽足形成,并不受GABA的影響。綜合實驗結果,說明GABA/GABA-A受體通路通過抑制iDCs偽足的形成來抑制細胞的遷移能力。見圖4。


3 討論
本研究發現,GABA/GABA-A受體通路可以抑制未成熟樹突狀細胞的遷移能力,當利用SiRNA敲低GABA-A受體α5亞基表達后,GABA/GABA-A受體通路則不再抑制未成熟樹突狀細胞的遷移。進一步研究發現,在GABA作用下,未成熟樹突狀細胞偽足生成明顯減少,提示了GABA/GABA-A受體通路可能通過抑制未成熟樹突狀細胞偽足的生成來影響細胞的遷移。
研究[11]證實,敲除漿細胞樣DCs 基因的AAA模型小鼠,動脈瘤的形成明顯減輕。在正常主動脈,少量DCs存在于內膜下并與平滑肌細胞和內皮細胞直接接觸;但在承受血流渦流壓力較大的部位,如動脈擴張處,大量成熟DCs呈簇狀聚集與T淋巴細胞接觸,進行抗原呈遞,激活T淋巴細胞的致炎活性[12]。同時,富集于病變血管處的未成熟DCs開始向血管外膜遷移,DCs胞內的運動裝置骨架蛋白發生構象改變,p38MAPK信號通路被激活,細胞骨架相關蛋白絲切蛋白去磷酸化,肌動蛋白單體在肌絲末端的解聚速度增加,DCs偽足伸出,遷移開始[13-14]。隨著DCs到達ECM,除借助偽足穿行外,尚需分泌相關蛋白酶降解ECM。未成熟DCs主要分泌基質金屬蛋白酶(matrix metalloproteinase,MMP)-1、MMP-2、MMP-9和基質金屬蛋白酶抑制劑(tissue inhibitor of metalloproteinase,TIMP)-1等,分泌過程與MAPK信號途徑中p38/ERK1/2的磷酸化狀態有關[15]。因此,抑制成熟DCs的抗原呈遞功能,削弱未成熟DCs的遷移及分泌MMPs的能力可有效阻止動脈瘤破裂。哺乳動物骨髓來源的DCs胞內含有GABA合成酶、GABA轉運蛋白和GABA-A受體亞基(α3,α5,β1,β3,ρ1)[4]。當弓形蟲感染DCs后,DCs內合成的GABA通過自體分泌的方式作用于GABA-A受體,調控DCs的運動;此過程中,細胞表面GABA-A受體的亞基表達也發生變化,α3,β3 和 ρ1亞基的轉錄明顯增加[16]。另有研究[17]發現,神經元突觸表面的GABA-A受體通過胞吞和受體再循環,能夠在細胞膜和細胞質之間發生遷移,如亞基α5發生膜受體內化后在埃茲蛋白-根蛋白-膜突蛋白(ezrin-radixin-moesin,ERM)家族中根蛋白的幫助下錨定肌動蛋白,從而引發抑制性突觸可塑性的改變。
綜上,GABA/GABA-A受體通路可以抑制未成熟樹突狀細胞的遷移能力。后續本研究小組將在體外實驗中繼續研究GABA/GABA-A受體通路對腹主動脈瘤小鼠樹突狀細胞抗原提呈功能的影響,從而為腹主動脈瘤的臨床治療尋找新的靶點。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
