Egr-1在單眼形覺剝奪性弱視幼貓視皮質(zhì)中的表達(dá)及其意義
2021-10-22 10:20:46范浩博唐秀平楊麗源宋唯琦王英陳思宇鄒云春
川北醫(yī)學(xué)院學(xué)報(bào) 2021年9期
關(guān)鍵詞:檢測(cè)
范浩博,唐秀平,楊麗源,宋唯琦,王英,陳思宇,鄒云春
(川北醫(yī)學(xué)院眼視光學(xué)系,四川 南充 637000)
弱視是造成當(dāng)今世界上兒童視力下降的重要疾病之一[1],亞洲地區(qū)的發(fā)病率約1.09%[2]。近年來,隨著分子生物學(xué)、神經(jīng)生物學(xué)等多學(xué)科在弱視致病機(jī)制方面的研究深入,目前研究[3-4]證實(shí),弱視治療的基本依據(jù)在于視覺發(fā)育敏感期內(nèi)視覺可塑性的存在,而視覺發(fā)育可塑性機(jī)制與多種神經(jīng)遞質(zhì)相關(guān),但具體發(fā)病機(jī)制尚未完全闡明,還不能在分子水平上詳細(xì)解釋弱視關(guān)鍵期及可塑性發(fā)生的變化。突觸目前被認(rèn)為是弱視的最關(guān)鍵環(huán)節(jié),其可塑性按時(shí)間上可分為長(zhǎng)時(shí)程增強(qiáng)(long-termpotentiation,LTP)和長(zhǎng)時(shí)程抑制(long-termdepression,LTD)。而早期生長(zhǎng)反應(yīng)因子-1(early growth responsive gene-1,Egr-1)作為即刻早期基因(Immediate-early genes,IEGs)中Egr家族內(nèi)的一員,具有編碼鋅指結(jié)構(gòu)的轉(zhuǎn)錄因子的作用。在突觸可塑性和記憶的鞏固以及LTP的誘導(dǎo)和學(xué)習(xí)過程中都伴隨著Egr-1表達(dá)的增加[5];同時(shí),Egr-1也具有將短時(shí)程信號(hào)與長(zhǎng)時(shí)程改變進(jìn)行偶聯(lián)的效應(yīng),但目前尚未有研究表明Egr-1的表達(dá)與弱視的關(guān)聯(lián)。本研究通過觀察形覺剝奪性弱視幼貓視皮質(zhì)Egr-1的表達(dá),探討Egr-1對(duì)視覺發(fā)育的作用及機(jī)制。
1 材料與方法
1.1 材料
1.1.1 實(shí)驗(yàn)動(dòng)物 選取3周齡幼貓30只,性別、毛色不限,體重240~350 g,眼底鏡檢查排除屈光介質(zhì)混濁及眼底異常;檢影驗(yàn)光顯示幼貓屈光度為+1.25~+3.25 D。由于幼貓?jiān)谀挲g較小階段尚無(wú)法自主攝取固態(tài)食物,故采取人工定時(shí)喂養(yǎng)貓奶粉與飲水的方式進(jìn)行飼養(yǎng),每日喂食4~5次,保證其健康成長(zhǎng)。待其能夠自主攝食后,每日早中晚共計(jì)3次,檢查貓糧與飲水的供應(yīng),以保證食物的充沛。飼養(yǎng)房間內(nèi)溫度設(shè)定為(24±1)℃,相對(duì)濕度保持在50%,房間內(nèi)各處均能夠接受自然光線照射。本研究動(dòng)物均由川北醫(yī)學(xué)院實(shí)驗(yàn)動(dòng)物中心提供,通過川北醫(yī)學(xué)院實(shí)驗(yàn)動(dòng)物倫理委員會(huì)批準(zhǔn)并全程接受監(jiān)督。
1.1.2 實(shí)驗(yàn)試劑與儀器 主要試劑:PBS(PH 7.2~7.6)緩沖液(武漢Boster公司,貨號(hào):AR0030)、枸櫞酸鹽(PH 6.0)緩沖液(武漢Boster公司,貨號(hào):AR0024)、Egr-1抗體(Bioss公司,貨號(hào)BS-1076R)、SABC免疫組化染色試劑盒(武漢Boster公司,貨號(hào):SA1022)、DAB顯色試劑盒(武漢Boster公司,貨號(hào):AR1022)、Mayer`蘇木素染液(Solarbio公司,貨號(hào):G1080)、伊紅染色液(Solarbio公司,貨號(hào):G1100)。主要儀器:動(dòng)物電極針(法國(guó)羅蘭,型號(hào):RL-1223000030-RC-D)、視覺電生理儀(法國(guó)羅蘭,型號(hào):RETI port/scan 21)、帶狀檢影鏡(蘇州六六,型號(hào):YZ24B)、病理切片機(jī)(Leica,型號(hào):RM2235)、顯微成像系統(tǒng)(尼康,型號(hào):DS-Fi2)
1.2 方法
1.2.1 動(dòng)物模型的建立 隨機(jī)數(shù)字表法將幼貓分為形覺剝奪組與對(duì)照組,每組15只。采用1%戊巴比妥鈉(35 mg/kg)腹腔注射麻醉兩組幼貓。對(duì)照組僅做麻醉處理;剝奪組在麻醉后采用“套環(huán)遮蓋法”[6]進(jìn)行弱視造模:將3-0線在其右眼眼眶四周對(duì)稱做四個(gè)小套環(huán),用縫線穿過套環(huán)連接黑色不透明遮蓋布,并于中央制作了一個(gè)高約0.7 cm,直徑為3 cm的環(huán)形遮蓋圈,防止光線從側(cè)面透過(圖1A)。剝奪組遮蓋前與遮蓋后的第1、3、5周,對(duì)剝奪組幼貓右眼、左眼與對(duì)照組右眼行圖形視覺誘發(fā)電位(pattern visual evoked potential,PVEP)檢測(cè)。PVEP檢測(cè)需矯正幼貓屈光不正,檢測(cè)前將幼貓遮蓋布套環(huán)上的連接線剪斷,取下遮蓋布,采用帶狀檢影鏡進(jìn)行檢影驗(yàn)光以確定幼貓雙眼屈光度(圖1B)。PVEP檢測(cè)時(shí),將三根動(dòng)物電極針分別依次插入:參考電極(藍(lán)色)置于前額正中皮下,作用電極(紅色)置于雙耳連線枕部皮膚正中皮下,接地電極(黑色)置于耳尖皮下。調(diào)整幼貓頭位使其被檢測(cè)眼視軸與屏幕中央處于同一水平線上(圖1C)。PVEP采用棋盤翻轉(zhuǎn)模式進(jìn)行刺激,距離為50 cm,刺激模式設(shè)定為0.3 cpd,視角16.7°,時(shí)間頻率為1 Hz,對(duì)比度97%,采樣時(shí)間300 ms,疊加64次。并根據(jù)此前檢影驗(yàn)光結(jié)果,選取相應(yīng)鏡片對(duì)其屈光不正進(jìn)行足矯,于暗室條件下進(jìn)行檢測(cè),每只眼重復(fù)三次取平均值。檢測(cè)過程中隨時(shí)觀察并調(diào)整幼貓頭位使其檢測(cè)眼視軸與屏幕中央處于同一水平線上。檢測(cè)后記錄剝奪組幼貓右眼、左眼與對(duì)照組幼貓右眼的P100波振幅與潛伏期。動(dòng)物弱視模型建立的成功標(biāo)準(zhǔn)以剝奪組右眼與剝奪組左眼、剝奪組右眼與對(duì)照組右眼之間P100波振幅與潛伏期存在統(tǒng)計(jì)學(xué)時(shí)即認(rèn)為弱視造模成功[7-8]。

1.2.2 取材與切片制作 遮蓋后第5周PVEP檢查完成后,根據(jù)《貓解剖彩色圖譜》將所有幼貓進(jìn)行視皮質(zhì)與外側(cè)膝狀體取材:使用解剖刀從枕外隆突處向前額骨處劃開皮膚,剝離顱骨外軟組織,使顱骨面暴露(圖2A)。使用直剪從枕骨處剪開顱骨,并向前端繼續(xù)分離兩側(cè)顱骨,直至腦組織完全暴露。用組織剪剪斷腦部組織與脊髓處的連接,用鑷子輕輕提起整個(gè)腦組織,從腦組織下方可看見視交叉,剪斷視交叉后,即可使整個(gè)腦部組織脫離。由背視視角即可見端腦枕回(圖2B);由腹視視角可見端腦梨狀葉,分離梨狀葉,暴露外側(cè)膝狀體(圖2C)。隨后快速切下端腦枕回與間腦外側(cè)膝狀體部,并立即將其放入4 %多聚甲醛固定液(含DEPC)中固定,隨后脫水包埋。采用石蠟切片機(jī)進(jìn)行切片,切片厚度設(shè)定為4 μm。

1.2.3 HE染色及免疫組化 HE染色:石蠟切片常規(guī)脫蠟至水,滴加蘇木素染液,1%鹽酸酒精分化后,滴加伊紅染液,脫水封片。顯微鏡下觀察細(xì)胞形態(tài)。免疫組化:石蠟切片常規(guī)脫蠟至水,3 % H2O2消除內(nèi)源性過氧化物酶活性,微波熱修復(fù)抗原,5 % BSA封閉液封閉,滴加一抗,滴加生物素標(biāo)記山羊抗兔IgG,滴加SABC,DAB顯色,蘇木素復(fù)染細(xì)胞核,脫水封片,顯微成像系統(tǒng)下進(jìn)行圖像采集分析陽(yáng)性細(xì)胞數(shù)與陽(yáng)性細(xì)胞平均光密度。
1.3 統(tǒng)計(jì)學(xué)分析
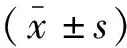
2 結(jié)果
2.1 PVEP檢測(cè)
PVEP檢測(cè)顯示,幼貓PVEP主波圖像表現(xiàn)為N75-P100-N135復(fù)合波,該波具有兩個(gè)正向波與一個(gè)負(fù)向波共同構(gòu)成,呈“M”狀(圖2.1)。遮蓋前,剝奪組右眼、左眼與對(duì)照組右眼在P100波潛伏期差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05);遮蓋3周后,三組P100波潛伏期與振幅差異有統(tǒng)計(jì)學(xué)意義(P<0.05);剝奪組右眼P100波潛伏期高于剝奪組左眼與對(duì)照組右眼(P<0.05);剝奪組右眼振幅低于剝奪組左眼與對(duì)照組右眼(P<0.05);遮蓋5周后,剝奪組右眼P100波潛伏期高于剝奪組左眼與對(duì)照組右眼(P<0.05);剝奪組右眼振幅低于剝奪組左眼與對(duì)照組右眼(P<0.05)。提示遮蓋3周后,剝奪組幼貓右眼已形成單眼形覺剝奪性弱視。見圖3、圖4、表1及表2。



表1 各組遮蓋前后P100波振幅變化

表2 各組遮蓋前后P100波潛伏期變化
2.2 HE染色
HE染色顯示,遮蓋5周后,對(duì)照組幼貓視皮層神經(jīng)元細(xì)胞形態(tài)以多角形、不規(guī)則型居多,細(xì)胞間結(jié)構(gòu)緊密;形覺剝奪組視皮層神經(jīng)元細(xì)胞形態(tài)多為圓形,少數(shù)呈不規(guī)則狀;局部膠質(zhì)細(xì)胞增生,海綿狀變性,部分神經(jīng)元水腫、變性。見圖5。

2.3 免疫組織化學(xué)檢測(cè)
免疫組化檢測(cè)結(jié)果表明,遮蓋5周后兩組幼貓大腦視皮質(zhì)中均有Egr-1的陽(yáng)性信號(hào),即存在Egr-1表達(dá),表達(dá)位于細(xì)胞胞漿之中,呈棕黃色,細(xì)胞核呈藍(lán)色。剝奪組中大部分呈現(xiàn)陽(yáng)性或弱陽(yáng)性,陽(yáng)性細(xì)胞數(shù)較少;對(duì)照組表達(dá)強(qiáng)陽(yáng)性信號(hào)較強(qiáng);剝奪組陽(yáng)性細(xì)胞數(shù)與平均光密度均低于對(duì)照組(P<0.05)。見表3及圖6。
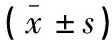
表3 免疫組織化學(xué)檢測(cè)

3 討論
形覺剝奪性弱視是由于嬰幼兒時(shí)期,因先天性白內(nèi)障、上瞼下垂、角膜白斑等原因,使光線不能或不正常進(jìn)入眼內(nèi),剝奪了黃斑接受正常光線刺激的機(jī)會(huì)。其生理性刺激的減少,使處于發(fā)育期內(nèi)的黃斑產(chǎn)生發(fā)育不良或停滯[1]。對(duì)于幼貓而言,形覺剝奪對(duì)其視力的影響一般在出生后至4周齡時(shí)達(dá)到頂峰,在12周齡后,遮蓋產(chǎn)生的形覺剝奪幾乎不能夠?qū)ζ湟暳Πl(fā)育進(jìn)行干擾[9]。因此,選用3周齡的幼貓通過單眼形覺剝奪的方式建立弱視模型。目前,形覺剝奪性弱視造模的經(jīng)典方法為上瞼縫合法,本小組既往曾采用“套環(huán)遮蓋法”[10],即利用黑色遮蓋布替代眼瞼縫合進(jìn)行弱視造模。在避免眼瞼縫合對(duì)眼前節(jié)產(chǎn)生損傷的同時(shí),利用剪斷連接線實(shí)現(xiàn)了對(duì)實(shí)驗(yàn)動(dòng)物PVEP變化的連續(xù)觀察。該方法可以有效阻擋動(dòng)物遮蓋眼各方向的光線進(jìn)入眼內(nèi),并順利建立弱視動(dòng)物模型。但造模期間內(nèi)仍出現(xiàn)個(gè)別幼貓頭部運(yùn)動(dòng)導(dǎo)致遮蓋布側(cè)方有光線透過。為改進(jìn)這一情況,本實(shí)驗(yàn)在原遮蓋方法上加大了遮蓋布的直徑,并在遮蓋布中央制成具有一定高度的黑色不透明遮蓋環(huán),以進(jìn)一步提升遮蓋效能。
本實(shí)驗(yàn)PVEP檢測(cè)采用棋盤翻轉(zhuǎn)刺激,記錄到幼貓PVEP主波圖像由兩個(gè)正向波與一個(gè)負(fù)向波構(gòu)成,呈“M型”。通過比對(duì)同時(shí)間不同眼與同眼不同時(shí)間P100波振幅與潛伏期,提示改良后的遮蓋方法是可行有效的。通過“套環(huán)遮蓋法”能夠成功建立單眼形覺剝奪性弱視動(dòng)物模型,并能實(shí)現(xiàn)對(duì)實(shí)驗(yàn)動(dòng)物的眼部生理參數(shù)動(dòng)態(tài)檢測(cè)。
隨著近年來弱視研究深入開展,多種參與視覺發(fā)育可塑性機(jī)制的神經(jīng)遞質(zhì),如AMPA受體及其GluR2亞基、NEP1-40、突觸素、膽堿能神經(jīng)元、生長(zhǎng)相關(guān)蛋白-43、GABA、cPKC-r、NMDAR、NGR-1等[11]都被認(rèn)為與弱視相關(guān),但仍然不能完全從分子水平上解釋弱視關(guān)鍵期與其可塑性變化的關(guān)系[3-4]。即刻早期基因作為一類可編碼轉(zhuǎn)錄因子的原癌基因[12],具有偶聯(lián)短時(shí)程信號(hào)與長(zhǎng)時(shí)程改變的效應(yīng),主要包括C-fos、Egr家族以及Arc等,其中有不少轉(zhuǎn)錄因子都受到視覺活動(dòng)的調(diào)節(jié)。其中,已有充分證據(jù)顯示C-fos與弱視疾病的關(guān)聯(lián)性。即刻早期基因中Egr家族內(nèi)的Egr-1[13],其轉(zhuǎn)錄因子在視皮層可塑性變化中是必需的。Egr-1是一種編碼鋅指結(jié)構(gòu)的轉(zhuǎn)錄因子,在突觸可塑性和記憶鞏固及LTP的誘導(dǎo)和學(xué)習(xí)過程中,都伴隨著Egr-1表達(dá)增加[5]。而細(xì)胞骨架相關(guān)基因Arc作為Egr-1的靶基因之一,是一個(gè)突觸活性誘導(dǎo)的效應(yīng)分子,在晚期LTP中具有重要作用。有研究[13]表明,一定條件下,Egr-1可以調(diào)控海馬CA1區(qū)晚期活性依賴性Arc基因轉(zhuǎn)錄,即刻早期基因Arc因其具有多能、精細(xì)調(diào)諧系統(tǒng),能夠連接神經(jīng)活動(dòng)的變化模式和突觸可塑性,從而優(yōu)化神經(jīng)系統(tǒng)信息存儲(chǔ)。本實(shí)驗(yàn)利用HE染色與免疫組化分別對(duì)遮蓋5周后的弱視幼貓視皮質(zhì)與對(duì)照組視皮質(zhì)的組織形態(tài)學(xué)差異和Egr-1蛋白表達(dá)進(jìn)行了對(duì)比與分析。提示在弱視幼貓模型中,視皮層神經(jīng)元細(xì)胞組織形態(tài)發(fā)生變化,并且伴隨局部膠質(zhì)細(xì)胞增生。既往弱視動(dòng)物模型的研究[14-15]發(fā)現(xiàn)弱視動(dòng)物的神經(jīng)節(jié)細(xì)胞、外側(cè)膝狀體、視皮質(zhì)都存在結(jié)構(gòu)方面的變化,并伴隨著突觸密度下降,這些結(jié)構(gòu)上的變化,又會(huì)導(dǎo)致其功能方面進(jìn)一步改變。本實(shí)驗(yàn)研究結(jié)果提示在視覺發(fā)育關(guān)鍵期內(nèi),由于雙眼視覺不對(duì)等輸入,可能會(huì)導(dǎo)致視皮質(zhì)細(xì)胞數(shù)目及形態(tài)發(fā)生變化,從而Egr-1蛋白表達(dá)下降,而Egr-1蛋白表達(dá)下降又進(jìn)一步對(duì)視皮質(zhì)的正常生理功能產(chǎn)生異常影響,影響視覺發(fā)育,從而促進(jìn)了弱視的發(fā)生發(fā)展。
視皮層軸突水平的興奮與抑制的平衡是維持視皮層正常發(fā)育和功能的條件,也是影響視覺系統(tǒng)可塑性的重要因素[16]。視神經(jīng)沖動(dòng)在腦的第一個(gè)突觸接替發(fā)生在外側(cè)膝狀體(LGN),每側(cè)的LGN接收來自同側(cè)視網(wǎng)膜顳側(cè)半視纖維和來自對(duì)側(cè)視網(wǎng)膜鼻側(cè)半視纖維的投射,這種投射具有嚴(yán)格的局部區(qū)域?qū)?yīng)關(guān)系。神經(jīng)的可塑性主要表現(xiàn)于突觸結(jié)構(gòu)和功能的可塑性。弱視發(fā)生與突觸可塑性改變有關(guān),弱視產(chǎn)生過程中會(huì)產(chǎn)生長(zhǎng)時(shí)程改變,Egr-1和Arc基因與突觸長(zhǎng)時(shí)程改變密切相關(guān)。目前多數(shù)研究也表明知覺學(xué)習(xí)可以提高弱視患者的視覺功能[17],知覺學(xué)習(xí)是通過反復(fù)的視覺刺激,以及具有監(jiān)督和反饋?zhàn)饔玫囊曈X體驗(yàn),使視功能得到有效的恢復(fù)[18]。研究者發(fā)現(xiàn)其機(jī)制可能與可塑性相關(guān)的興奮和抑制平衡有關(guān)聯(lián),而Egr-1和Arc又在人與動(dòng)物的學(xué)習(xí)記憶中發(fā)揮重要作用[19]。本實(shí)驗(yàn)研究進(jìn)一步證實(shí)了在弱視發(fā)病過程中,視皮質(zhì)的Egr-1蛋白表達(dá)會(huì)產(chǎn)生下降,進(jìn)一步驗(yàn)證了知覺學(xué)習(xí)在弱視治療中的作用。
綜上所述,采用套環(huán)遮蓋法可以實(shí)現(xiàn)弱視動(dòng)物模型的建立和眼球參數(shù)的動(dòng)態(tài)測(cè)量。基于這一方法所制作的弱視動(dòng)物模型,發(fā)現(xiàn)Egr-1在視皮質(zhì)的表達(dá)明顯下降。本實(shí)驗(yàn)推測(cè)Egr-1在視覺發(fā)育過程中起到了重要作用。本研究為進(jìn)一步探索單眼形覺剝奪是如何調(diào)控視皮層神經(jīng)元,為治療弱視與深入了解其發(fā)病機(jī)制提供了新的思路與方向。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
 川北醫(yī)學(xué)院學(xué)報(bào)2021年9期
川北醫(yī)學(xué)院學(xué)報(bào)2021年9期
- 川北醫(yī)學(xué)院學(xué)報(bào)的其它文章
- 護(hù)理人員職業(yè)緊張對(duì)職業(yè)倦怠的影響研究
- miR-182-5p和自噬在外周血單個(gè)核細(xì)胞和THP-1細(xì)胞痛風(fēng)性關(guān)節(jié)炎模型炎癥反應(yīng)中的研究
- 極化液對(duì)內(nèi)毒素休克兔腦氧化損傷的影響
- TLR4 mRNA和TCF21 mRNA在喉癌組織中的表達(dá)及其與臨床病理的相關(guān)性
- 帕金森病伴疼痛患者接觸性熱痛刺激誘發(fā)的腦功能磁共振成像研究
- 分化型甲狀腺癌頸部轉(zhuǎn)移性淋巴結(jié)FNA-Tg測(cè)定結(jié)合超聲風(fēng)險(xiǎn)特征參數(shù)診斷效能的研究