珠江口及其鄰近海域赤潮物種的生物多樣性研究進展
2021-10-19 09:05:46姚艷欣陳楠生
海洋科學 2021年9期
關鍵詞:物種
姚艷欣 , 陳楠生
(1. 中國科學院海洋研究所 海洋生態與環境科學重點實驗室, 山東 青島 266071; 2. 青島海洋科學與技術試點國家實驗室 海洋生態與環境科學功能實驗室, 山東 青島 266071; 3. 中國科學院海洋大科學研究中心, 山東青島 266071; 4. 中國科學院大學, 北京 100049)
珠江又名粵江, 是長江以南最大的河流, 也是中國的第三大河流。珠江口水域系指21°30′~22°49′N,113°30′~114°30′E的海域[1], 包括珠江的河口灣, 毗鄰粵港澳三地, 面積約為210 km2[1]。珠江口海域浮游植物按照地理位置可以劃分為河口類、近岸類和外海類三大生態類群[2]。珠江口海域由于受到珠江水系徑流匯入的影響, 加上廣東沿岸流、南海暖流和黑潮的影響, 具有豐富的營養鹽。該海域浮游植物物種多樣性較高, 物種組成跨度大, 其中豐水期以淡水種和廣溫廣鹽種為主, 而枯水期以廣溫廣鹽種為主[2]。
赤潮(red tide)是一種由海洋藻類、原生動物或細菌在水體中過度繁殖或聚集導致海水變色的全球性海洋生態異常現象, 目前國際上更多稱其為有害藻華(harmful algal blooms, HABs)[3]。近年來珠江流域經濟迅速發展, 廣東省城市人口急劇增加, 沿岸地區大量的工業和農業廢水直接排放進珠江口, 廢水中含有大量的有機物、重金屬、無機鹽和污染物, 導致珠江口海域出現嚴重的污染。另外, 珠江口海域大規模的水產養殖業也是其重要的污染物來源。這些污染源導致珠江口海域的營養鹽濃度不斷升高, 尤其是大量的含氮營養鹽, 使得水體富營養化趨勢日益嚴重, 成為赤潮暴發的主要原因[2]。另外, 壓艙水導致的外來有毒有害藻類的引入、核電站的溫排水影響及底層休眠孢囊的存在可能也促成了珠江口海域赤潮暴發[4-5]。
赤潮研究的方法有很多, 主要包括利用形態和分子方法對赤潮物種進行分類、追蹤和生理生化分析, 研究赤潮的時空分布特征以及與環境相互作用的關系[6-8], 并對其進行防治研究[9-12]。近年來, 各種組學技術不斷應用于赤潮研究中, 對物種鑒定、發現新種以及對赤潮生物的組成、分布、動態變化等方面都有很大的提升。除此之外, 遙感技術、地理信息系統(geography information system, GIS)以及現場調查的方式進行調查, 建立赤潮監測數據庫, 為赤潮防治奠定基礎[13-18]。本文綜述了珠江口及其鄰近海域浮游植物, 重點分析了珠江口及其鄰近海域的赤潮物種的生物多樣性研究進展, 為研究珠江口浮游植物物種和赤潮物種組成和動態變化提供了幫助,對解讀宏組學以及宏條形碼分析結果、分析珠江口浮游植物和赤潮物種的動態變化提供了基礎。
1 珠江口及其鄰近海域的赤潮
1980年以來, 隨著廣州經濟的高速發展, 污水的排放量增加, 珠江口及其近海海域污染嚴重, 海域內營養鹽含量迅速升高, 成為有害藻華的高發地帶, 暴發頻次逐漸升高。1982年11月上旬南海內伶仃洋淺灘首次報道了甲藻赤潮[5]。李永振在1990—1991年在廣東沿岸河口海域采集浮游植物樣品, 并結合近幾年廣東沿岸河口海域采樣分析資料, 報道了47種硅藻赤潮物種[19]。據統計, 1987—2017年珠江口及其近海海域共發生了454次赤潮[4]。珠江口及其近海海域一年四季均有赤潮發生, 其中春季赤潮暴發頻率最高, 其次是夏秋冬季節。4月和8月為甲藻赤潮暴發的高峰期, 其中4月主要赤潮物種為夜光藻(Noctiluca scintillans)、米氏凱倫藻(Karenia mikimotoi)、裸甲藻(Gymnodiniumspp.)、雙胞旋溝藻(Cochlodinium geminatum)和多環旋溝藻(Cochlodinium polykrikoides), 8月多暴發錐狀斯氏藻(Scrippsiella trochoidea)赤潮, 5月為硅藻赤潮暴發的高峰期, 其中中肋骨條藻(Skeletonema costatum)被認為是赤潮暴發的主要原因種。秋冬季節多暴發定鞭藻赤潮, 其中棕囊藻(Phaeocystisspp.)占主導[4]。王漢奎等人分析了2002年6月珠江口海域無紋環溝藻(Gyrodinium instriatum)赤潮的生長消亡過程, 認為海域富營養化是誘發此次赤潮的主要原因[20]。雙胞旋溝藻赤潮多次出現在珠江口海域。2006年10月至11月和2009年10月, 珠江口海域暴發了雙胞旋溝藻赤潮, 研究人員發現富營養化水體和適宜其生長的溫度促使了該藻赤潮的暴發[21]。在雙胞旋溝藻赤潮發生后期, 海水中出現雙胞旋溝藻休眠期的群體細胞[22]。暴發赤潮的原因很復雜, 其中營養鹽濃度的升高是此次赤潮暴發的重要原因之一[16]。雙胞旋溝藻赤潮暴發時會產生致畸物視黃醛, 嚴重危害浮游動物[23]。雙胞旋溝藻赤潮對鹵蟲幼體也有一定的毒害作用[24]。
2 珠江口及其鄰近海域的航次調查
通過對公開發表的航次調查數據分析, 我們總結了1985—2014年間22個針對珠江口及其鄰近海域的浮游植物和赤潮物種調查(附表1)。從空間上來看, 這些航次調查站位集中于珠江口及其鄰近海域,即18°~24°N, 109°~119°E范圍內, 幾乎涵蓋了珠江口及沿岸海域所有的赤潮暴發區, 共描述了幾百種浮游植物物種以及幾十種重要的赤潮物種, 體現了珠江口及鄰近海域浮游植物和赤潮物種的生物多樣性。從時間上來看, 航次調查時間跨度越長, 盡管大部分調查集中在夏季, 我們仍然可以通過它們了解該海域浮游植物物種的組成和變化, 并了解赤潮物種時空變化, 對珠江口以及沿岸海域浮游植物物種和赤潮物種的研究有重要的參考價值。
3 珠江口及其鄰近海域的浮游植物
3.1 珠江口及其鄰近海域浮游植物全長18S rDNA序列統計分析及優勢浮游物種分析
針對珠江口的22個航次調查總共報道了428種浮游植物, 包括硅藻門(Bacillariophyta)270種, 甲藻門(Dinophyta)130種, 綠藻門(Chlorophyta)9種, 藍藻門(Cyanophyta)8種, 定鞭藻門(Coccolithophore)5種,金藻門(Chrysophyta)4種, 著色鞭毛藻門(Cryptophyceae)1種和裸藻門(Euglenophyta)1種(附表2)。
在這428種浮游植物物種中, 有120種的全長18S rDNA序列(>1 600 bp)可以在公共數據庫NCBI中查到, 不到總數的三分之一(28.0%), 表明珠江口及其鄰近海域浮游植物物種中絕大多數物種還沒有得到分子分析。
利用數據庫中查找到的全長18S rDNA序列構建系統進化樹發現珠江口及其鄰近海域的浮游植物聚類到6個門, 其中以硅藻門物種最多(72個), 其次是甲藻門(36個)。在珠江口及其鄰近海域歷年的航次調查中出現頻率最高的浮游植物包括中肋骨條藻、菱形海線藻(Thalassionema nitzschioides)、洛氏角毛藻(Chaetoceros lorenzianus)、柔弱偽菱形藻(Pseudonitzschia delicatissima)、鐵氏束毛藻(Trichodesmium thiebautii)、紅海束毛藻(Trichodesmium erythraeum)和叉狀角藻(Ceratium furca)(圖1b、1c)。
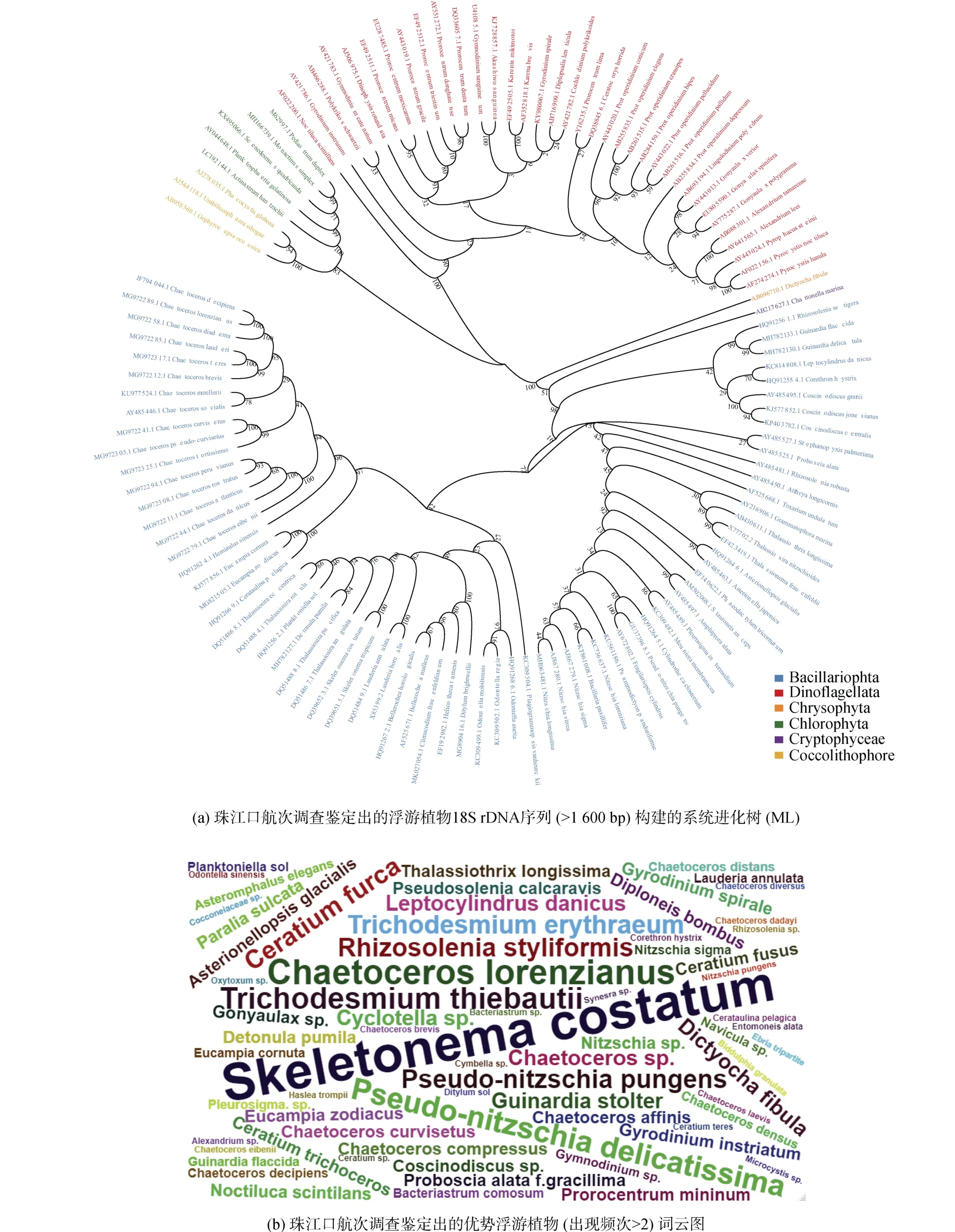
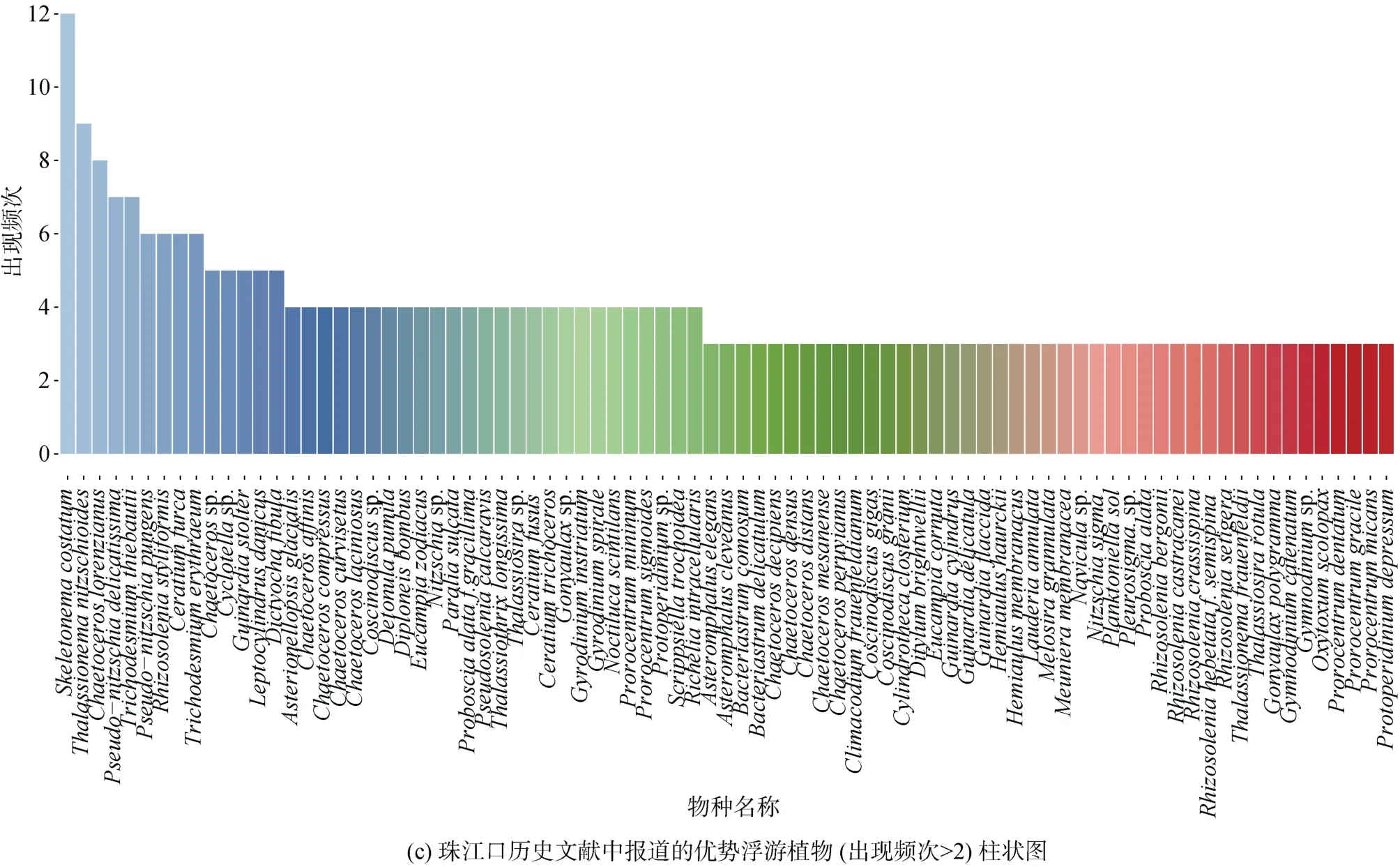
圖1 珠江口浮游植物物種組成Fig. 1 Composition of phytoplankton species in the Pearl River Estuary
3.2 珠江口及其鄰近海域航次調查結果的比較分析
通過對航次調查結果的比較分析(圖2a)發現,不同航次鑒定出的浮游植物物種組成差異較大, 較多浮游植物物種只在單一航次中都被調查到, 并且沒有任何一種浮游植物物種出現在所有的航次調查結果中。造成這種調查結果差異的原因有多種, 比如航次調查季節存在差異, 航次調查時采樣范圍不盡不同, 航次調查針對的浮游植物對象具有不同的聚焦, 對浮游植物的分析方法不同等。
硅藻和甲藻是浮游植物物種中的主要組成部分,赤潮物種中多數種類也以硅藻和甲藻為主。有研究表明珠江口海域優勢物種趨于簡單, 小型硅藻的數量逐漸增加, 優勢度顯著提升[25]。本文利用22個航次中甲藻和硅藻的數目以及各自相對所占的比例來跟蹤分析歷年來珠江口及其鄰近海域甲藻和硅藻物種數目的相對變化。結果表明, 歷年來珠江口及其鄰近海域中甲藻和硅藻物種相對數目并沒有顯示出明顯的變化規律(圖2b)。不過, 上面的分析可能不足以充分揭示甲藻和硅藻物種數目的相對變化規律, 因為這些航次各自獨立, 不同航次采樣目的也不相同,航次采樣季節的不同, 物種分類及分析標準存在差異, 并不能準確跟蹤珠江口浮游植物相對豐度的變化。基于形態的分析方法與基于分子生物學(比如比較宏條形碼分析[26])的分析方法的聯合使用, 將促進珠江口及其鄰近海域浮游植物組成的更加準確的判斷、鑒定和分析。
萊茵哈德·根策爾成長于德國黑森林里的小城市弗萊堡。年輕的時候,他是德國最好的標槍運動員之一,甚至能與國家隊一起備戰1972年慕尼黑奧運會。現在,他的“標槍”擲向了更遠的地方。

圖2 珠江口浮游植物組成的歷史變遷Fig. 2 Historical changes in the composition of phytoplankton species in the Pearl River Estuary
4 珠江口及其鄰近海域常見赤潮物種
珠江口及其附近海域的這些航次調查共鑒定了5個門54種常見的赤潮物種, 包括硅藻門25種、甲藻門24種、金藻門2種、藍藻門2種以及著色鞭毛藻門1種(表1)。這些物種為珠江口及其鄰近海域常見的赤潮物種, 且在珠江口及其鄰近海域暴發的次數較多。不同航次報道的赤潮物種的種類和數目有顯著差異, 在圖3a中可以發現, 不同航次調查結果差異較大, 并且沒有一個赤潮物種出現在所有航次調查中(圖3a)。其中最常見的赤潮物種有中肋骨條藻、洛氏角毛藻、柔弱偽菱形藻、鐵氏束毛藻、梭角藻(Ceratium fusus)、尖刺偽菱形藻(Pseudonitzschia pungens)、紅海束毛藻、叉狀角藻、丹麥細柱藻(Leptocylindrus danicus)和微小原甲藻(Prorocentrum mininum)(圖3b、3c)。此外, 米氏凱倫藻、夜光藻(Noctiluca scintilans)、雙胞旋溝藻和球形棕囊藻(Phaeocystis globosa)也是珠江口海域經常暴發的赤潮物種。
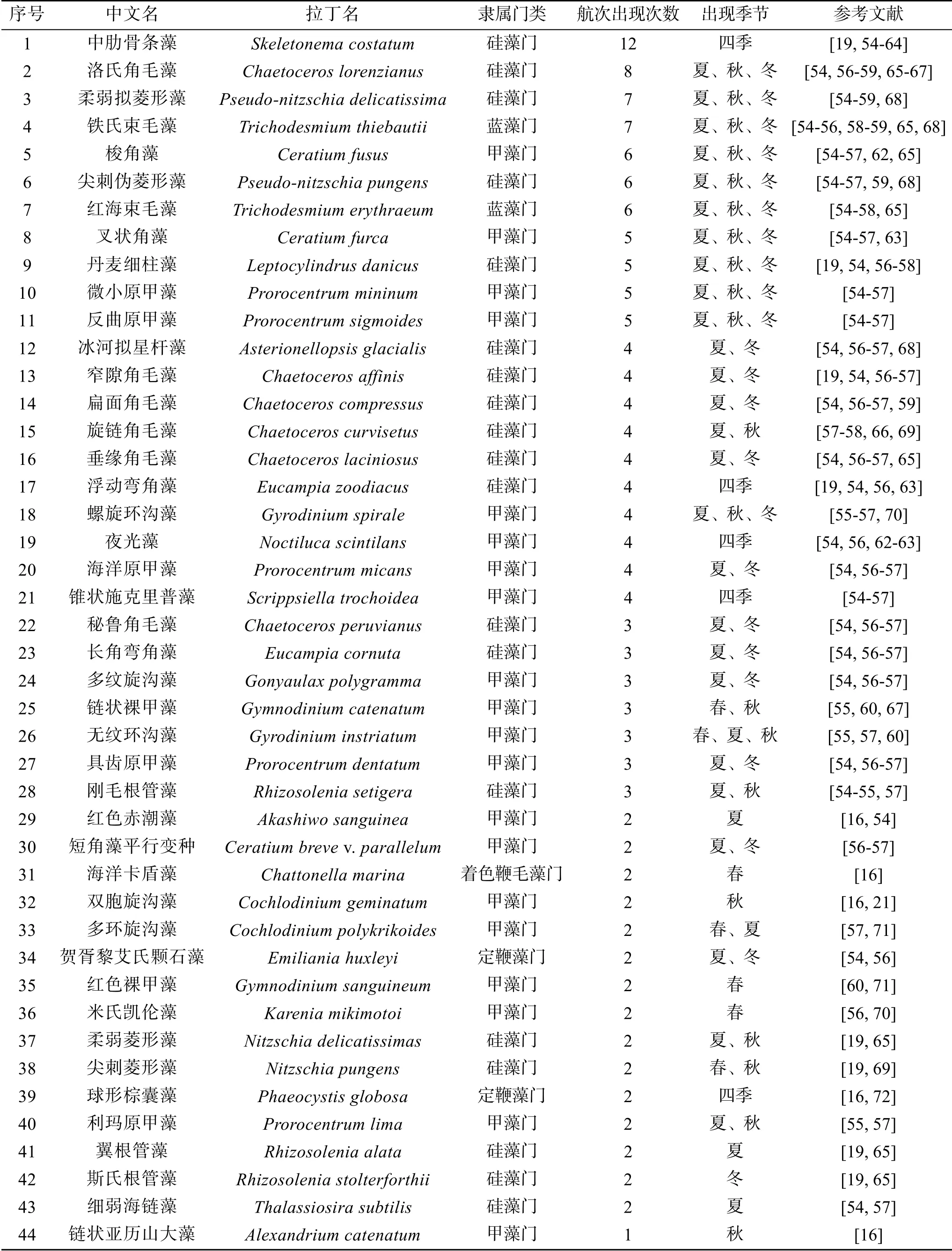
表1 珠江口歷史航次赤潮物種名錄Tab. 1 HAB species reported in the Pearl River Estuary in historical expeditions
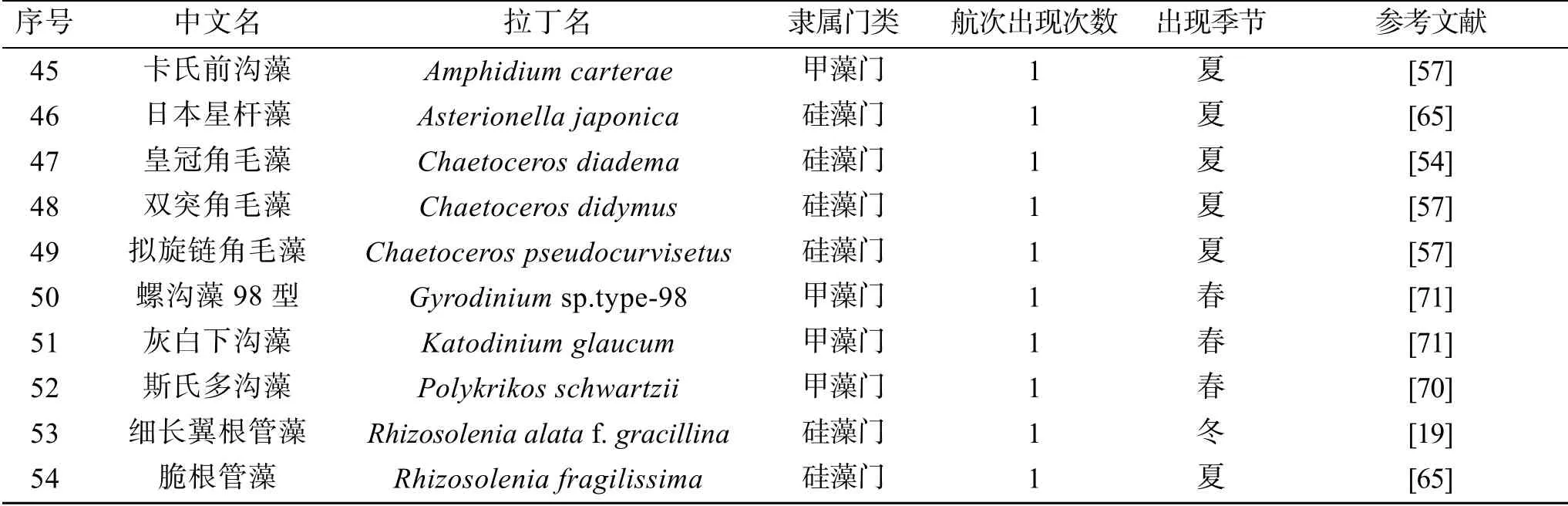
續表

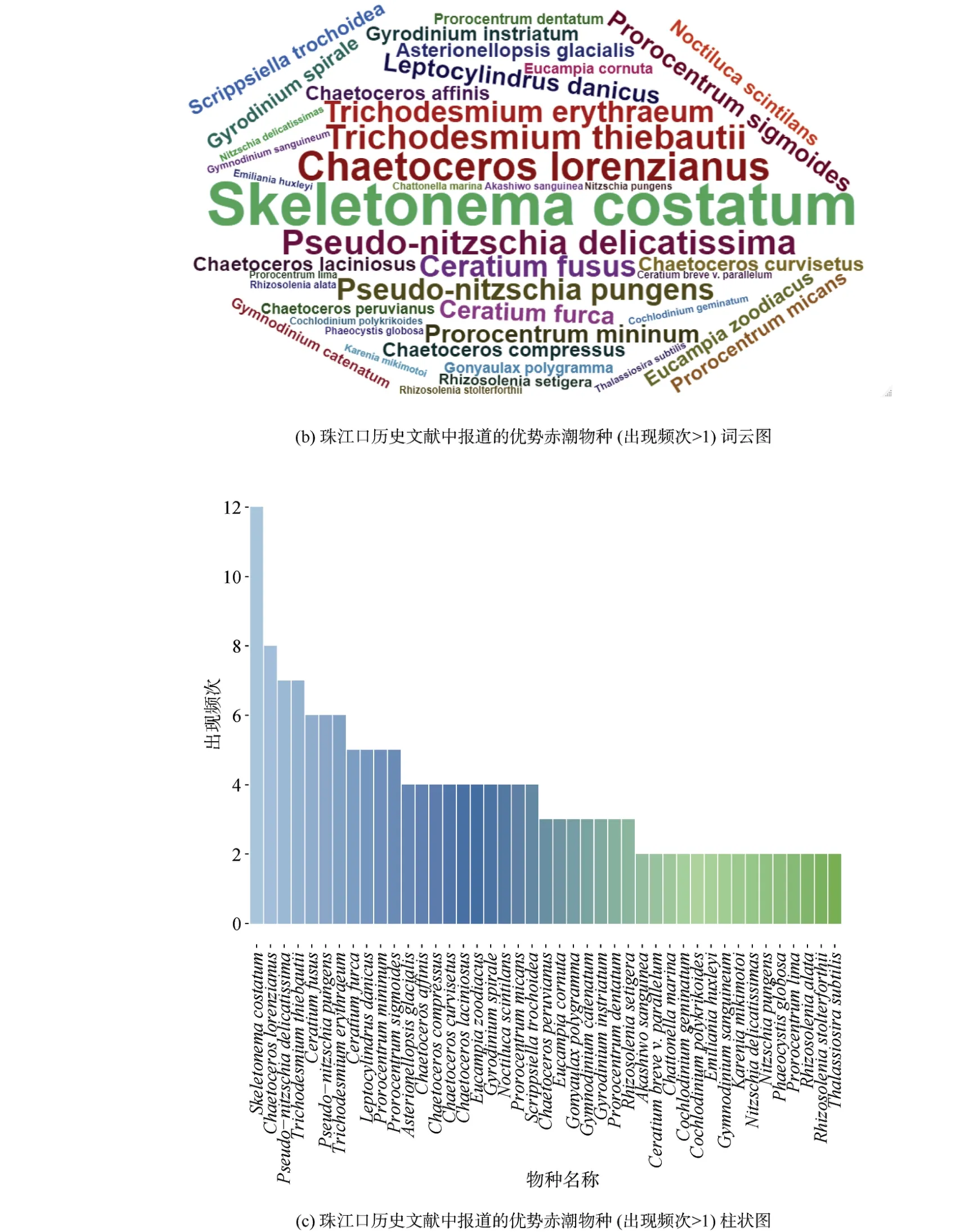
圖3 珠江口赤潮物種組成(出現頻次>1)Fig. 3 Composition of phytoplankton species in the Pearl River Estuary
4.1 中肋骨條藻
中肋骨條藻隸屬于硅藻門(Bacillariophyta), 中型硅藻綱(Mediophyceae), 海鏈藻目(Thalassiosirales), 骨條藻科(Skeletonemataceae), 骨條藻屬(Skeletonema), 細胞為圓柱形或透鏡形, 直徑大小6~7 μm,殼面鼓起, 相鄰細胞間通過刺連成長鏈, 無毒, 屬于浮游廣布種, 是廣溫廣鹽的典型代表[19]。分布于朝鮮、阿拉斯加灣、白令海、黑海、歐洲沿岸等海區, 中國包括珠江口海域的各個海區均有出現[19]。該種是珠江口—大亞灣海域秋季優勢種, 細胞總量可以達到238萬個/m3[19]。中肋骨條藻也是川山群島海域秋季優勢種, 細胞總量可以達到12萬個/m3[19]。自1990年以來, 中肋骨條藻在大鵬灣水域多次引發赤潮[19]。2002年6月在珠江口發生了無紋環溝藻和中肋骨條藻的雙相赤潮[27]。據統計, 自1980—2017年間, 硅藻中的主要赤潮種類為中肋骨條藻, 有記錄的中肋骨條藻赤潮為43起, 其中時間分布中春季發生次數最多, 珠江口附近暴發次數最多[4]。中肋骨條藻在珠江口分布格局呈現為由內河口向外河口遞增的趨勢。此研究還表明中肋骨條藻的豐度和硅酸鹽呈現顯著的正相關關系, 和磷酸鹽呈現顯著的負相關關系, 兩者都是影響珠江口海域中肋骨條藻豐度的重要因素[28]。有研究者利用中肋骨條藻作為赤潮代表種, 滸苔作為綠潮代表種, 表明在未來全球氣候變暖和海洋酸化的環境下, 赤潮比綠潮的影響更大[29]。研究者通過對日本海域中分離得到的中肋骨條藻全基因組序列分析發現, 該種有與氧化應激反應和細胞分裂素相關的多拷貝基因, 并在暴發赤潮時得到表達[30]。
4.2 雙胞旋溝藻
雙胞旋溝藻隸屬于甲藻門(Dinophyta), 橫裂甲藻綱(Dinophceae), 裸甲藻目(Gymnodiniales), 裸甲藻科(Gymnodiniaceae), 和旋溝藻屬(Cochlodinium)[75],有游泳細胞和成對細胞兩種形態, 細胞近橢圓形,背腹略扁, 細胞長寬為30~40 μm, 色素體為網狀黃褐色[21]。該藻有休眠期細胞, 細胞呈卵形或深褐色,通過黏液相連成群, 在赤潮暴發期間不產生毒素[21]。2006年4月在廣東珠海桂山島附近海域首次暴發了雙胞旋溝藻赤潮, 同年10至11月期間又暴發了兩次該種引發的赤潮[21]。雙胞旋溝藻易與同屬的多環旋溝藻比較容易混淆, 關于該藻赤潮暴發的種源問題還有待進一步的研究[21]。2009年10月至11月再次暴發大規模的該藻赤潮, 富營養化水平高是此次赤潮暴發的基礎。2018年11月期間在珠江口暴發雙胞旋溝藻赤潮, 在赤潮期間, 浮游植物群落結構發生了改變, 優勢浮游物種的數量明顯下降。同時赤潮期間視黃醛的含量升高, 視黃醛可導致水生生物發生畸變, 對水生生物具有潛在的威脅[31]。
4.3 夜光藻
夜光藻隸屬于甲藻門(Dinophyta), 橫裂甲藻綱(Dinophceae), 環溝藻目(Gymnodiniales), 夜光藻科(Noctiluceae)和夜光藻屬(Noctiluca), 是一種分布于世界沿岸的廣布型低鹽發光種, 是完全異養型浮游生物, 同時具有植物的屬性和動物的功能[32]。夜光藻利用分泌黏液的觸手將食物送入泡口。水體中營養鹽含量的增加促進其他浮游生物的生長來間接促進夜光藻的繁殖[32]。夜光藻的大量繁殖并不直接依賴于水體的高營養環境, 不能單一地將水體富營養化程度作為該藻赤潮暴發的標準[33]。長江口及其鄰近海域、南海海域都是夜光藻暴發的高發海域, 自20世紀30年代首次暴發夜光藻赤潮以來, 赤潮范圍不斷擴大, 直至覆蓋了中國近海所有的海域[34]。夜光藻在暴發赤潮時, 其空間分布格局受到生物因素和物理因素的綜合影響, 但是主要受到風、潮汐和海流等物理過程的影響[35]。一方面, 夜光藻會產生氨, 在堿性條件下危害有鰓的水生動物[36]; 另一方面, 夜光藻還會分泌大量黏液, 導致魚類以及其他水生生物死亡[37]。此外, 在赤潮衰亡之后, 細胞內會有大量的內噬細菌釋放到水體中, 這些內噬細菌會產生有毒物質, 威脅水生生物的生存[32]。田媛等收集了1980—2017年廣東沿岸赤潮發生數據, 結果發現珠江口大鵬灣海域多發生夜光藻赤潮, 其中在223起甲藻赤潮中就有77起是夜光藻赤潮[4]。
4.4 球形棕囊藻
球形棕囊藻隸屬于定鞭藻門(Haptophyta), 顆石藻綱(Coccolithophyceae), 棕囊藻目(Phaeocystales),棕囊藻科(Phaeocystaceae)和棕囊藻屬(Phaeocystis),是廣溫廣鹽性藻類[38], 能產生溶血性毒素, 是全球海洋廣泛分布的赤潮物種[39]。該種有復雜的異型生活史, 存在有單細胞和膠質囊體兩種形態[40]。在富營養條件下能夠暴發性增殖形成赤潮, 大量膠質囊體的形成不僅有效抵御細菌、病毒侵入及浮游動物的攝食[41], 在赤潮發生過程中產生溶血性毒素、二甲基硫化物(DMSP/DMS)及硫丙酸等化合物毒害海洋生物[42]。球形棕囊藻溫度適應范圍明顯高于其他藻類,能夠在短時間內暴發赤潮, 嚴重影響了海洋生態系統的結構和功能, 同時對海洋漁業以及養殖業造成了巨大損失。1997年10月中國東南沿海首次暴發球形棕囊藻赤潮, 覆蓋范圍廣, 從福建泉州、廈門以南直至廣東汕尾海域, 持續時間長達6個月, 經濟損失嚴重。據不完全統計, 從1997年至2017年20年間,共暴發球形棕囊藻赤潮50起, 面積超過10 000 km3,嚴重影響了海洋生態系統正常的結構和功能。球形棕囊藻赤潮多發生在汕頭汕尾海域, 據統計, 在1980—2017年間, 廣東省沿岸共發生該藻赤潮39起[4]。此外, 球形棕囊藻能夠產生溶血性毒素, 嚴重危害海洋中魚類以及其他海洋生物的生存[39]。我國對球形棕囊藻赤潮防治的方法有很多, 主要有生物方法、化學方法、光催化法以及黏土礦物絮凝法等, 此外有研究者利用海洋放線細菌對有毒球形棕囊藻囊體進行有效地移除[43]。
4.5 米氏凱倫藻
米氏凱倫藻隸屬于甲藻門(Dinophyta), 橫裂甲藻綱(Dinophceae), 裸甲藻目(Gymnodiniales), 凱倫藻科(Kareniaceae)和凱倫藻屬(Karenia), 藻體呈單細胞,細胞背腹扁平, 呈寬卵圓形。細胞長20~30 μm, 寬16~30 μm, 側面寬度10~15 μm, 長略大于寬[44]。細胞上殼部為半球形或寬圓錐形。下殼部底部中央有凹陷,為兩淺裂片狀。下殼部大于上殼部[44]。該藻為游泳生活, 主要分布在溫帶和熱帶海域, 是一種常見的有毒有害赤潮物種, 在廣東省沿岸春季為多發季節[4]。1980年, 該藻首次在香港海域暴發赤潮, 1998年3月中旬至4月中旬, 在珠江口海域暴發了一次嚴重的米氏凱倫藻赤潮, 持續時間近30 d, 造成了大規模的魚類和貝類死亡, 并且值得注意的是1998年在廣東省沿岸海域就發生了8起米氏凱倫藻赤潮[4]。2002年,該藻首次在東海暴發赤潮[45]。2001年至2006年間, 我國共發生米氏凱倫藻赤潮59起[46]。米氏凱倫藻赤潮是我國近海魚毒性赤潮的典型代表種, 該藻能產生細胞毒素和溶血性毒素等一系列毒素, 這些毒素會造成大量魚類死亡。此外, 還會影響浮游動物的生長發育、攝食、產卵等生命活動。比如, 米氏凱倫藻能影響部分敏感貝類幼體的早期發育, 可致其幼體和成體死亡[44]。有相關研究表明, 雖然米氏凱倫藻的代謝產物有毒, 但是卻為抗腫瘤藥物以及新藥的設計提供了潛在的可能[46]。
4.6 洛氏角毛藻
4.7 鐵氏束毛藻
鐵氏束毛藻隸屬于藍藻門(Cyanobacteria), 藍藻綱(Cyanophyceae), 顫藻目(Oscillatoriales), 微鞘藻科(Microcoleaceae)和束毛藻屬(Trichodesmium), 是全球性廣布型藍藻, 普遍存在于熱帶、亞熱帶和溫帶海域中。藻絲體放射狀排列成球形或中部扭曲成繩狀, 束中藻絲體的排列較亂, 但頂端游離。藻絲直徑6~16 μm, 長與寬相等或2~8倍于寬。在太平洋和大西洋海域經常會形成優勢物種, 被認為是東海和黃海黑潮流系的指示種[50]。鐵氏束毛藻是一種非常重要的固氮藍藻, 無異形胞。在貧營養海域中, 該種生物量占主導, 成為海洋初級生產力和全球氮循環過程的重要組成部分。該藻在暴發赤潮時會產生毒素, 導致海洋中魚類、蝦類和雙殼類等海洋生物的死亡[51]。中國海域固氮藍藻中鐵氏束毛藻成為東海低氧區和鄰近海域的優勢物種, 并且細胞豐度在8月份最高。透明度會影響鐵氏束毛藻的生長, 溫度會促進其生長, 而鹽度對其生長的影響不顯著[52]。在本文統計的22個航次的歷史文獻中, 該藻共發生過7次赤潮。
5 主要結論與展望
本文通過對珠江口及其鄰近海域歷史航次資料整理, 主要有以下幾點總結:
1) 本文綜合了1985—2014年間22個航次調查結果, 共整理了428種浮游植物物種, 其中硅藻270種,甲藻130種, 綠藻9種, 藍藻8種, 定鞭藻5種, 金藻4種, 著色鞭毛藻1種和裸藻1種。這表明珠江口海域浮游植物物種組成豐富, 多樣性較高。
2) 我們共統計了54種赤潮物種, 其中硅藻25種,甲藻24種, 定鞭藻2種, 藍藻2種, 著色鞭毛藻1種。其中暴發頻率最高的赤潮物種為中肋骨條藻, 其次是洛氏角毛藻和柔弱偽菱形藻。珠江口及其鄰近海域甲藻和硅藻赤潮物種數不相上下, 有些甲藻赤潮物種,比如雙胞旋溝藻和米氏凱倫藻, 一旦暴發赤潮, 其危害程度遠遠大于硅藻赤潮物種, 應該引起充分的重視。分析發現珠江口及其鄰近海域硅藻和甲藻的相對組成在1985—2014年間沒有顯著變化。不過, 由于航次調查結果分析時的方法有限以及各個航次采樣目的可能不同等原因, 此結論會存在一定的誤差, 需要結合將來的數據進一步核實。
3) 通過分析比較歷年的航次結果數據可以發現,航次之間報道的物種豐度以及物種多樣性方面還是存在一定的差距, 可能是因為航次調查時間、海域,以及對樣品的處理方式存在不同, 但是最主要的原因是物種鑒定方法存在一定的局限性, 最初航次調查時利用傳統的形態學鑒定方法會遺漏許多隱存物種, 相對降低了物種多樣性。
4) 對珠江口歷年航次中統計到的浮游植物物種構建進化樹時發現, 數據庫中并不是所有的物種都有18S rDNA的分子標記, 并且單個物種的代表性分子標記(如18S rDNA, 28S rDNA, ITS,cox1,rbcL)種類不全面, 限制了基因組宏條形碼分析結果的解讀和分析。
因此, 基于以上幾點結論, 我們建議:
1) 對于珠江口及其鄰近海域赤潮高度暴發海區,增加對航次采樣的頻率, 結合遙感技術、GIS以及以往的赤潮數據庫對赤潮暴發時間和海域進行及時預測, 避免造成嚴重的生態災害和經濟損失。
2) 對浮游植物物種進行系統分析, 補充相應的18S rDNA的分子標記序列, 并且豐富分子標記的種類, 特別是對于高度頻發的赤潮物種, 豐富數據庫,支撐宏條形碼分析方法對赤潮物種進行解讀和分析。近年來, 分子分析方法迅速發展, 基于DNA分子標記擴增和測序的宏條形碼分析方法(metabarcoding analysis)逐漸應用到野外樣本的研究中, 并已取得很多有益的成果。比如, 有研究者將形態學觀察和基于18S rDNA V2-V3區的高通量測序分析相結合, 發現黃海和東海海域的寄生性甲藻在高通量測序方法中占優勢, 卻在形態學研究中被忽略[53], 可見分子分析方法能夠彌補傳統形態學研究的不足。通過采用具有高分辨率和高特異性的分子生物學方法, 同時結合傳統鏡檢鑒定方法, 對數據進行深入準確的分析, 全面揭示自然樣本中物種的組成及相對豐度, 從而系統了解珠江口及其鄰近海域浮游植物尤其是赤潮物種的詳細信息, 豐富珠江口及其鄰近海域赤潮物種數據庫數據, 進而對赤潮暴發時間進行準確預測和預報, 對赤潮物種進行準確的鑒定和分析, 推進赤潮研究領域更進一步。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
