熱帶珊瑚礁區紅腹海參及綠刺參食物來源分析
——脂肪酸標志法
2021-10-19 09:05:30吳沛霖黃端杰馬文剛王愛民
海洋科學 2021年9期
吳沛霖, 黃端杰, 馬文剛, 高 菲, 王愛民, 許 強
(海南大學 海洋學院 南海海洋資源利用國家重點實驗室, 海南 海口 570228)
近幾十年來, 隨著國際市場對海參產品需求量的持續提高, 海參捕撈在熱帶海域得到了迅速發展[1-2]。在市場需求和海參自身存在易于捕撈, 且資源恢復速度較慢等特點的影響下, 大多數具有較高經濟價值的海參種類資源面臨著被過度開發的問題[3-4]。世界各地熱帶海參資源也已經顯現出枯竭的跡象[5-6]。因此, 迫切需要采取適當的種群保護和恢復措施,以確保這些物種的可持續利用。紅腹海參(Holothuria edulis)及綠刺參(Stichopus chloronotus)分別屬于楯手目(Aspidochirotida)的海參科(Holothuriidae)及刺參科(Stichopododae), 是印度洋-太平洋區域大部分地區分布非常廣泛的海參種類[7-8]。它們多棲息于珊瑚礁淺水區域的海草床和砂質海底, 以及具有藻類和海草分布的巖礁區[9]。紅腹海參和綠刺參皆為沉積食性海參, 它們以沉積物中的動植物有機質碎屑、小型底棲生物、微生物等為食, 在珊瑚礁生態系統的物質循環中發揮著重要的作用。但是目前對兩種海參詳細的食物組成及其季節差異知之甚少。與此同時, 我國熱帶海域海參資源衰退趨勢日益嚴峻[10], 特別是海南島周邊海域的紅腹海參和綠刺參資源日趨稀少, 對其攝食生態及食物來源的了解也有助于開展資源恢復工作。
生活在珊瑚礁中的熱帶海參, 它們通過攝食和埋棲等行為對其生活環境產生影響, 進而影響到動物的營養環境等[11], 對海洋生態系統產生十分重要的影響[12]。海洋動物所需的大多數脂肪酸都是從食物中獲得的, 然后被相對保守地同化至消費者的組織中[13], 食物中的脂肪酸特征得以保留。而海洋食物網中的脂肪酸主要來源于海洋自養藻類的合成。每種海洋藻類因其不同的脂肪酸合成途徑而具有自己的特征性脂肪酸[14-17]。Zhang等[18]人利用脂肪酸標志法調查研究了不同海域內刺參(Apostichopus japonicus)的脂肪酸組成和食物來源, 發現不同海域內的同種海參在食物來源上存在較大差異, Xu等[19]人的研究結果也證實了環境會對海參脂肪酸組成造成影響。因此, 海參的脂肪酸組成不同, 不僅受其自身生理生態特性、季節、緯度和個體生長發育階段的影響[18-19], 還會受食物來源影響。
本研究以海南三亞蜈支洲島珊瑚礁海域的紅腹海參及綠刺參為研究對象, 分析了這2種海參夏季和冬季的脂肪酸組成情況, 并利用脂肪酸標志法探討2種海參的食物來源異同, 揭示其種間及季節差異, 研究結果可為揭示紅腹海參及綠刺參生態功能提供數據支持。
1 材料與方法
1.1 研究區域概況
蜈支洲島位于海南島南部(18°18′30′′N, 109°45′40′E)(圖1), 面積1.48 km2[9], 是典型的近岸熱帶島礁。該地區屬于熱帶季風氣候, 11月至4月為旱季, 5月至10月為雨季。在夏季和冬季, 該海域受季風的影響比較明顯[20]。在夏季, 海南島東南部沿海區域會出現上升流(QDU)[21]。實驗區域位于蜈支洲島北部(圖1)。該海區覆蓋著細沙、石塊、珊瑚碎枝以及人工增殖礁。
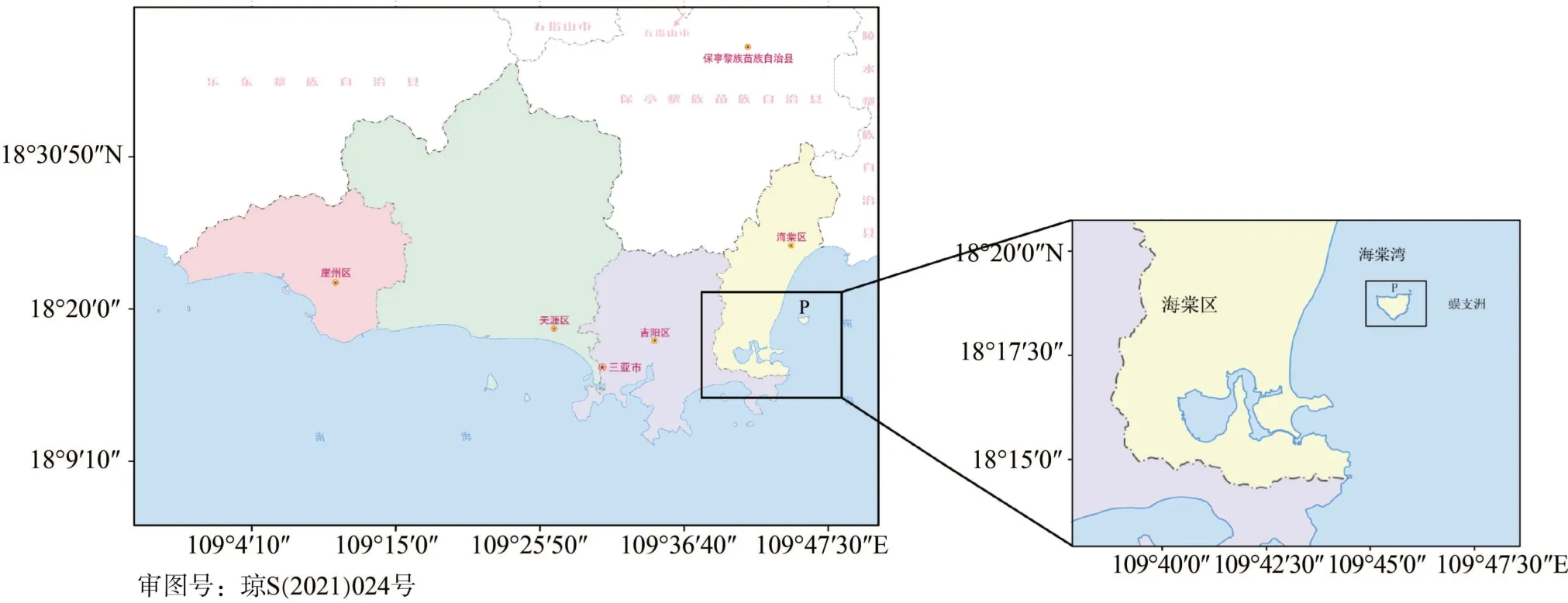
圖1 取樣區域Fig. 1 Sampling area
1.2 樣品采集及溫度、鹽度的測定
在2019年, 利用多參數水質分析儀(YSI-Model 6600v2, Xylem, 美國)監測了蜈支洲島海域水深1 m處的水溫和鹽度。
紅腹海參及綠刺參樣品采集于2019年7月25日與12月21日, 采集地點位于蜈支洲島北部海域(圖1)。由潛水員在實驗區域內采集紅腹海參和綠刺參, 每種海參每次采集3~5頭, 采集后半小時內運到實驗室進行取樣。每次采集的個體大小如表1所示。

表1 不同月份采集海參的個體大小(mean±SD)Tab. 1 Individual size of sea cucumbers collected in different months (mean±SD)
由于消化道組織對食物脂肪酸組成的變化響應最快[22], 因此本研究選擇海參腸道組織作為脂肪酸組成及標志物的分析對象。將采集到的紅腹海參和綠刺參放置在暫養箱(40 cm×30 cm)中, 待海參狀態穩定后, 立即注射3~5 mL的KCl溶液(0.35 mol/L)刺激紅腹海參和綠刺參排出內臟[23]。將海參腸道從排出的內臟團中分離, 小心去除腸道含物, 利用蒸餾水清洗干凈后, 放置于-20 ℃冰柜中冷凍保存備用。
1.3 樣品預處理與脂肪酸測定
海參腸道樣品經冷凍干燥48 h后, 研磨成粉末狀。取約0.5 g左右的樣品進行總脂提取, 過程參考Floch等[24]的方法。經二氯甲烷∶甲醇(2∶1)提取總脂后, 可保存至二氯甲烷中。在溫和的氮氣流下將提取的總脂蒸發至接近干燥, 然后加入2%的濃硫酸甲醇溶液并在氮氣的保護下于80 ℃水浴中甲酯化2 h, 利用酯化反應制備脂肪酸甲酯(FAME)[25]。冷卻后用正己烷萃取脂肪酸甲酯, 樣品定容至1 mL后上機測定。脂肪酸組成利用氣相色譜儀(Agilent Technologies 6890N)進行分析。以魚肝油脂肪酸甲酯(Cod liver oil fatty acid methyl esters(C2294-5G), Sigma-Aldrich, 德國)作為標準, 通過比對保留時間對脂肪酸進行定性分析。脂肪酸的相對含量用峰面積歸一法進行計算。
色譜條件如下:
毛細管色譜柱: DB-FFAP(30 m × 0.25 mm × 0.25 μm);
載氣: 氦氣(純度為99.999%);
進樣口溫度: 250 ℃;
檢測器溫度: 250 ℃;
柱溫: 初始溫度為100 ℃, 并在100 ℃下保持5 min, 以4 ℃/min的速度升至240 ℃, 然后在240 ℃下保持15 min。
1.4 脂肪酸標志物的選擇
綜合以往的研究結果與海區內實際情況選用以下脂肪酸標志: 硅藻、鞭毛藻及原生動物、褐藻、綠藻、紅藻、異養細菌以及陸源有機質等幾類(詳見第3節)。
1.5 統計分析
采用數據分析軟件SPSS (PASW Statistics 23, 美國)進行數據分析, 利用獨立樣本t檢驗(Independent-Samples T Test)分析不同月份、不同物種是否有顯著差異性。數據在進行分析前利用Levene檢驗進行均勻性檢驗。在 RStudio 1.3.1093軟件中利用相關矩陣對不同月份、不同物種的脂肪酸標志物數據進行標準化, 然后進行主成分分析(PCA)。
2 結果
2.1 水溫與鹽度
2019年, 實驗區域鹽度為32.58至34.44, 溫度為23.76 ℃至29.31 ℃。7月份時, 受海南島東部上升流的影響, 海水溫度下降了3 ℃(圖2)。
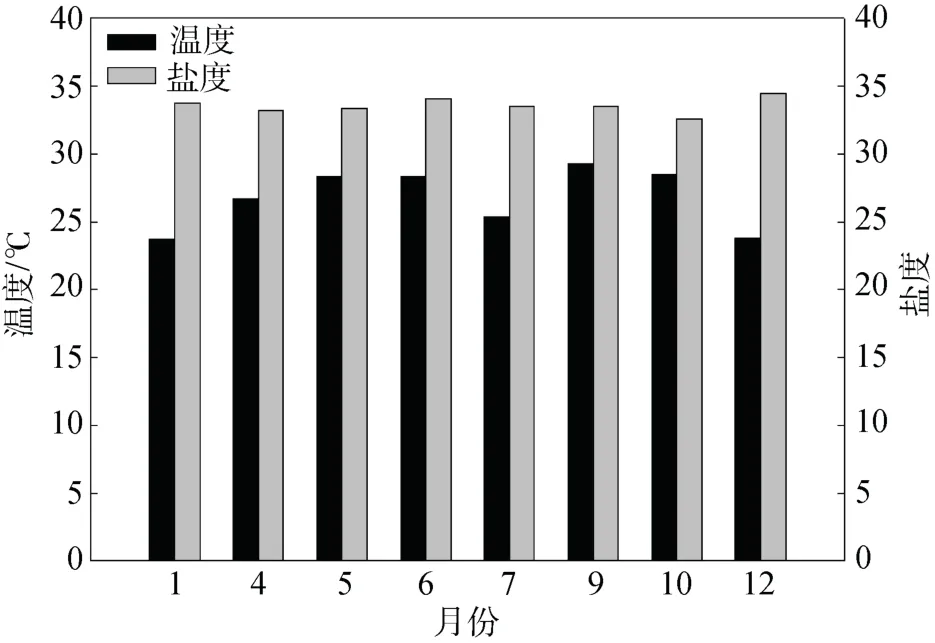
圖2 取樣地點水溫與鹽度隨時間的變化Fig. 2 Temporal variations in the water temperature and salinity of the study site
2.2 綠刺參及紅腹海參的脂肪酸組成
本研究對綠刺參及紅腹海參中14—22碳的脂肪酸進行了測定, 并分離出33種脂肪酸(表2), 發現2種海參在不同月份的脂肪酸相對含量均以多不飽和脂肪酸(PUFA)為主(圖3)。2種海參TFA的含量在12月份較高, 但飽和脂肪酸(SFA)、單不飽和脂肪酸(MUFA)以及PUFA含量均表現出相反的季節差異。綠刺參的PUFA在12月份較高(P>0.05), SFA和MUFA含量較低(P>0.05), 紅腹海參則相反。
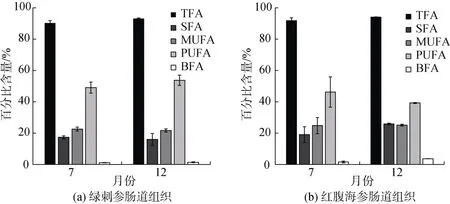
圖3 綠刺參腸道組織(a)與紅腹海參腸道組織(b)脂肪酸組成相對含量月份變化Fig. 3 Fatty acid profiles of the gut of S. chloronotus (a) and H. edulis (b) in different months
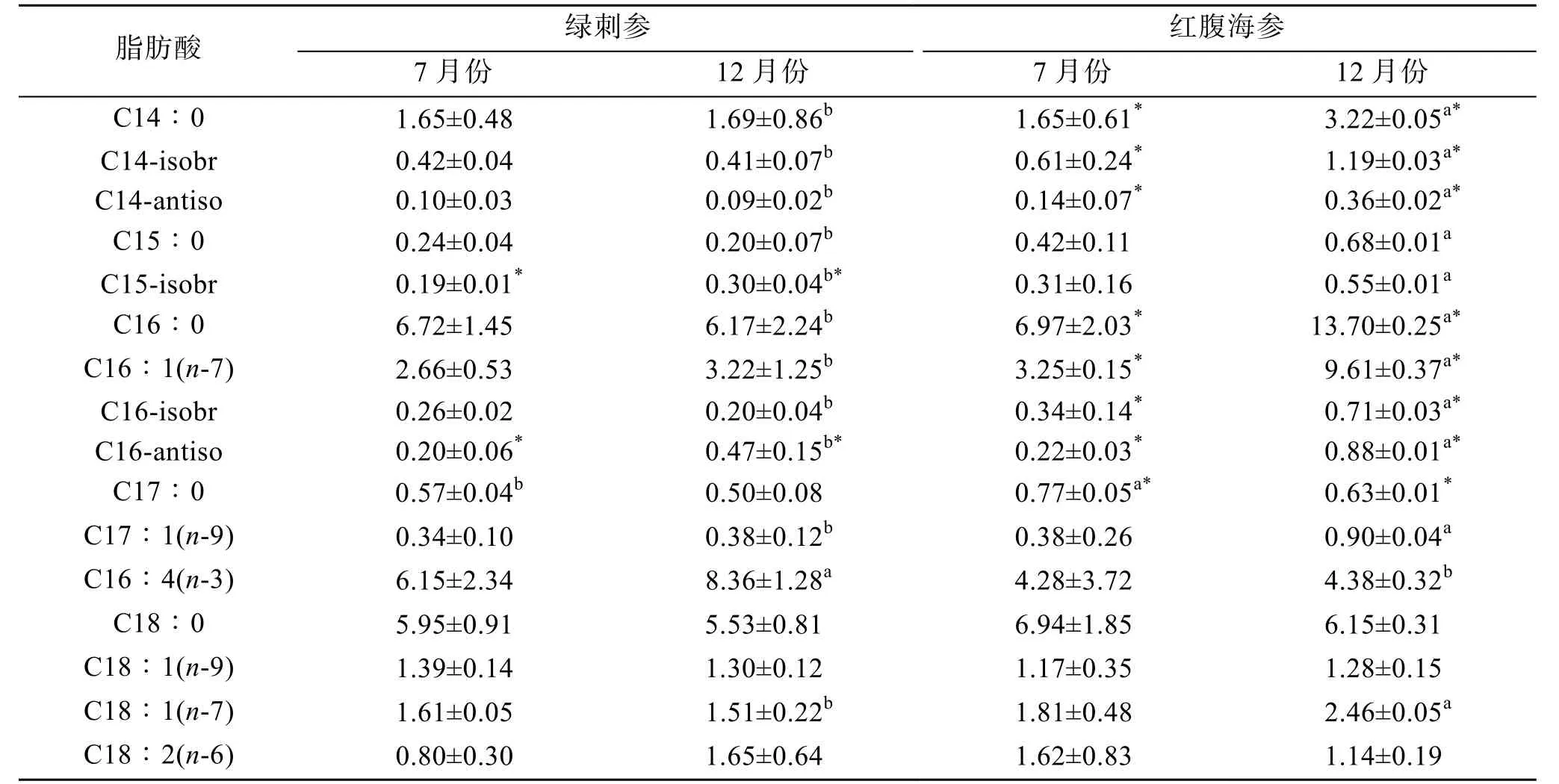
表2 綠刺參、紅腹海參腸道組織脂肪酸相對含量(%, mean±SD)Tab. 2 Relative content of fatty acids in the gut of H. edulis and S. chloronotus (%, mean ± SD)
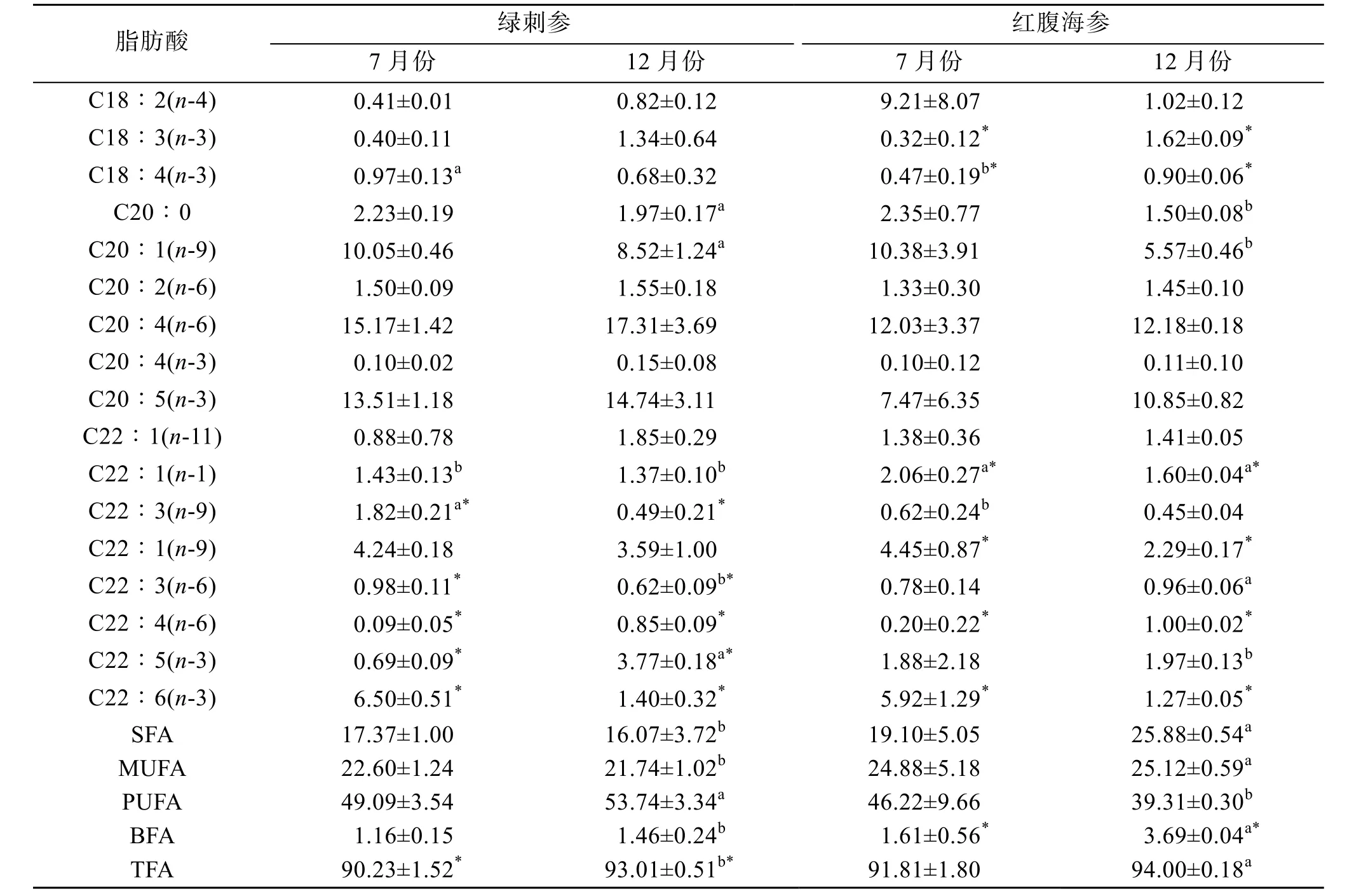
續表
2種海參的SFA主要由16∶0和18∶0組成, 均占SFA含量的50%以上。2種海參的18∶0含量在12月份時較低(P>0.05)。綠刺參16∶0的含量是7月份高于12月份, 與紅腹海參相反, 紅腹海參明顯7月份時較低(P>0.05)。2種海參MUFA的主要成分20∶1(n-9)和22∶1(n-9)含量均是7月份較高。
綠刺參與紅腹海參的PUFA含量較高, 占TFA的40%以上。在2種海參組織中含量最高的脂肪酸為C20∶4(n-6), 在12月份時較高(P>0.05)。在7月份時, DHA[C22∶6(n-3)]含量在綠刺參中最高(6.5%),紅腹海參DHA的含量為5.92%; 在12月份時, 綠刺參與紅腹海參的DHA含量均明顯低于7月份(P>0.05)。紅腹海參與綠刺參組織中的EPA[C20∶5(n-3)]含量在12月份最高, 7月份較低。
2.3 綠刺參及紅腹海參的脂肪酸標志物季節差異
紅腹海參與綠刺參的硅藻脂肪酸標志物C16∶1/C16∶0比值均小于1(圖4a), 這表明硅藻不是兩種海參的食物來源組成部分。根據表3, EPA還可以作為紅藻的特征脂肪酸。因此, 12月份時兩種海參體內有較高的EPA含量可能攝入環境中更多的紅藻有機質碎屑。7月份時紅腹海參的EPA含量較低, 且存在較大的個體差異(圖4b)。
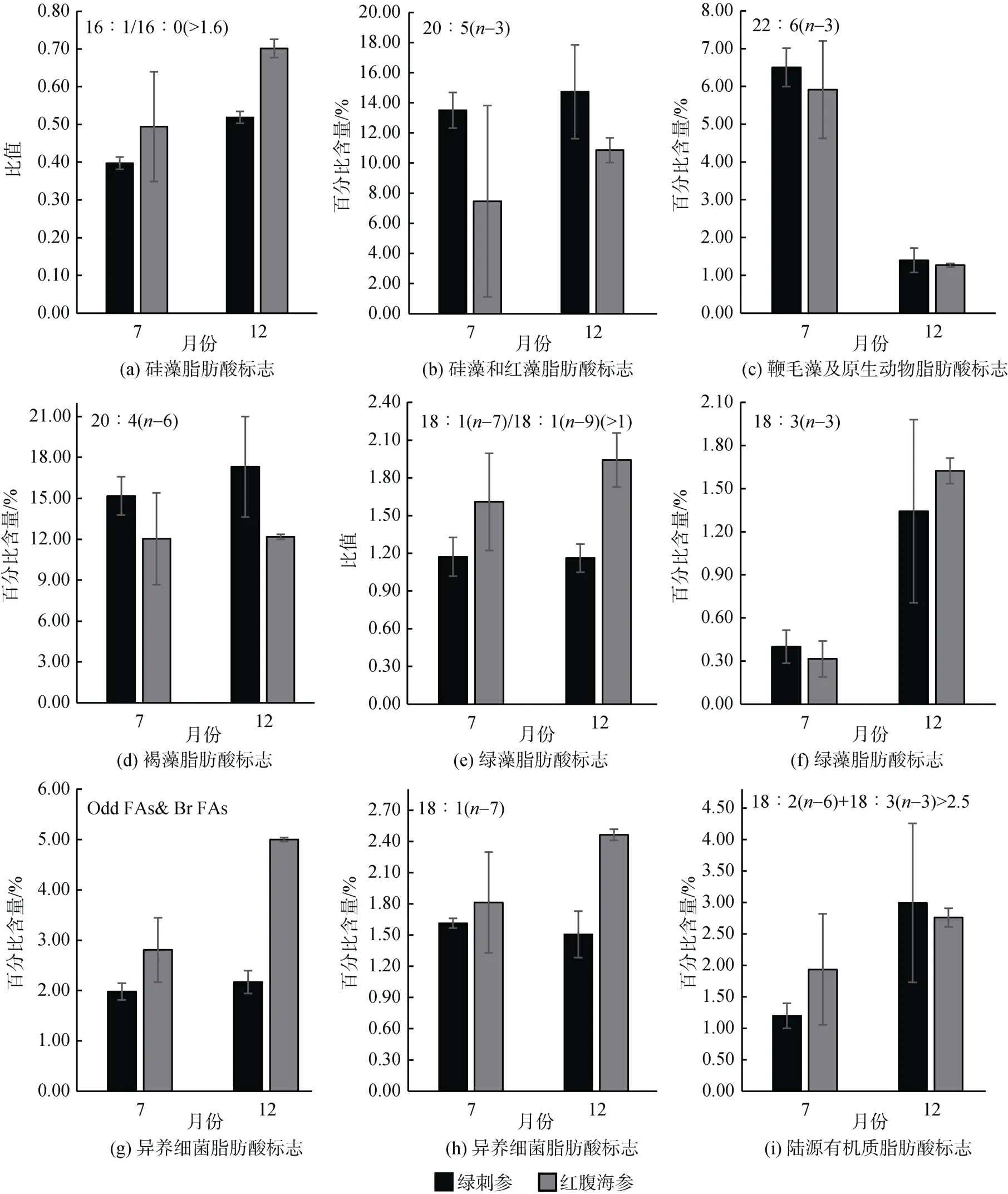
圖4 綠刺參與紅腹海參脂肪酸標志的月份變化Fig. 4 Fatty acid biomarkers of S. chloronotus and H. edulis in different months
紅腹海參與綠刺參的鞭毛藻及原生動物脂肪酸標志物DHA含量分別為5.92%和6.5%。同時綠刺參和紅腹海參中DHA的含量存在明顯的季節差異, 且2種海參都表現出7月份較高, 12月份較低(圖4c)。
紅腹海參和綠刺參組織中的褐藻脂肪酸標志物C20∶4(n-6)含量較其他生物標志物均具有較高水平,分別為12.03%~12.18%和15.17%~17.31%, 且無明顯的季節差異。這表明褐藻類食物來源在不同月份均是2種海參的重要食物來源(圖4d)。
綠刺參與紅腹海參體內的綠藻脂肪酸標志物C18∶1(n-7)/C18∶1(n-9)比值在不同季節均大于1(圖4e), 且無明顯的季節差異(P>0.05), 這表明綠藻也是2種海參的食物來源之一。另一種綠藻脂肪酸標志18∶3(n-3)在2種海參體內表現出相似的季節差異, 7月份的含量較低。12月份時綠刺參的18∶3(n-3)含量較高, 也表現出較大的個體差異。
Odd FAs & Br FAs與C18∶1(n-7)是異養細菌的脂肪酸標志物。綠刺參體內的Odd FAs & Br FAs與C18∶1(n-7)的含量在不同季節均無明顯的差異(P>0.05)(圖4g, 4h)。紅腹海參體內的Odd FAs & Br FAs與C18∶1(n-7)的含量均為12月份較高, 且Odd FAs& Br FAs含量有明顯的季節差異(P<0.05)。
C18∶2(n-6)+C18∶3(n-3)是陸源有機質的脂肪酸標志物。與7月份相比, 12月份2種海參體內的含量均較高, 且大于2.5(圖4i), 這表明陸源有機質在12月份時是2種海參的食物來源。7月份紅腹海參和12月份綠刺參的陸源有機質脂肪酸標志含量均存在個體差異。
在2種海參食物來源組成中, 褐藻、綠藻、紅藻均是重要的食物來源, 同時還有異養細菌、陸源有機質、鞭毛藻以及原生動物等。2種海參表現出攝食大型藻類有機質碎屑的情況, 同時也有對其他食物來源的需求。
2.4 脂肪酸標志物種間及季節差異性分析
以2種海參腸道中8種脂肪酸標志物[C20∶4(n-6)、C20∶5(n-3)、C22∶6(n-3)、C18∶1(n-7)、C18∶3(n-3)、C18∶1(n-7)/C18∶1(n-9)>1, Odd FAs&Br FAs, C18∶2(n-6)+ C18∶3(n-3)]的含量或比值為變量, 對不同季節采集的海參樣品數據進行主成分分析, 以期得出2種海參食物組成的種間與季節差異(圖5)。研究結果發現, 不同季節的2種海參形成顯著的聚類區系。7月份時, 綠刺參以鞭毛藻及原生動物為主要食物來源; 12月份綠刺參的食物來源較為廣泛, 涵蓋鞭毛藻及原生動物、異養細菌褐藻和紅藻等。紅腹海參7月份的食物來源顯現出典型的褐藻和紅藻類特征, 而12月份則主要為異養細菌和綠藻特征。
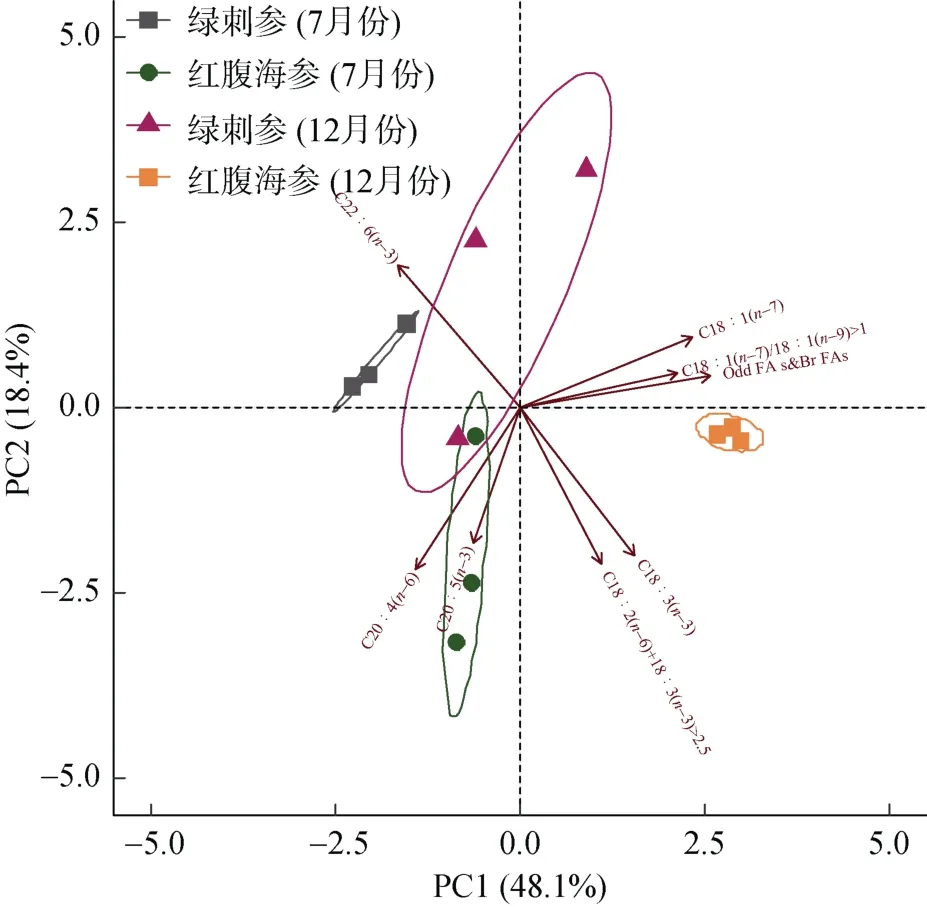
圖5 綠刺參及紅腹海參不同月份脂肪酸標志物主成分分析因子載荷圖Fig. 5 PCA of food resources component in H. edulis and S.chloronotus in different months
3 討論
海參的食物組成會受到海區天然餌料環境的影響, 例如, 大型藻類的有機碎屑, 微型藻類, 以及細菌等等通常具有不同的物理結構和生物化學組成, 這都會影響海參的食物偏好[26-28]。以往的研究表明, 紅腹海參可棲息于沙底, 活珊瑚以及礁石上, 它們對于生境的選擇沒有明顯的偏向性[9]。綠刺參偏向選擇活珊瑚及礁石等硬質底的生境,其腸道內含物中也多以泥砂為主, 混有大型藻類有機質碎屑、細菌、原生動物等[29]。本研究根據海區實際情況, 以及以往的研究結果選擇了幾類脂肪酸標志物對海南三亞海域的紅腹海參及綠刺參食物來源分析發現(表3), 它們的食物來源可能包括鞭毛藻及原生動物、褐藻、綠藻、紅藻以及異養細菌等。
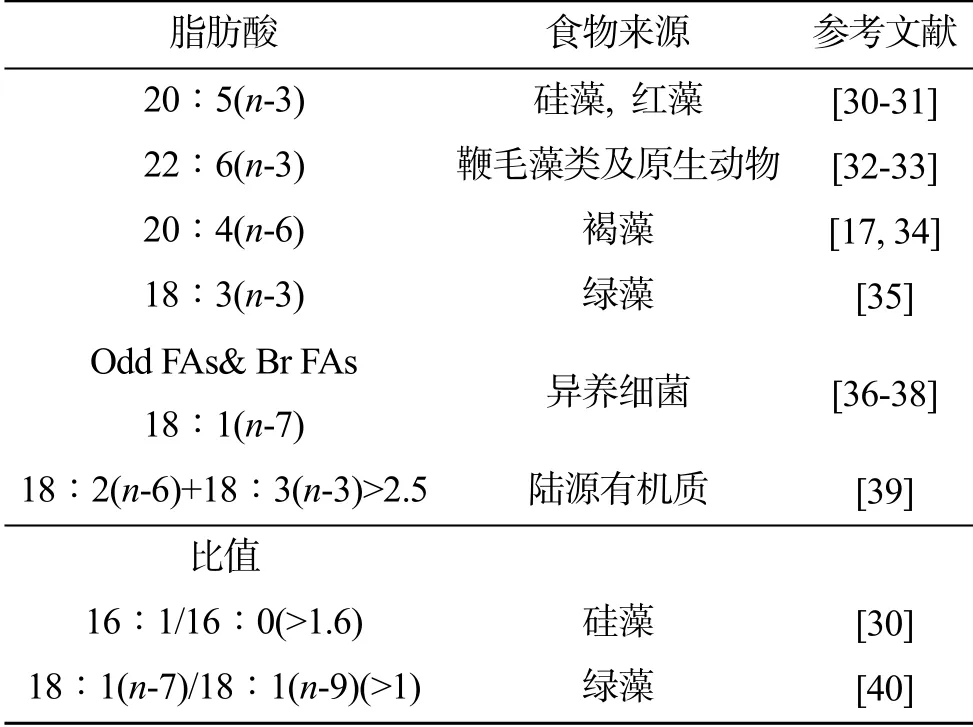
表3 作為食物來源標志的脂肪酸和脂肪酸比值Tab. 3 Fatty acids and fatty acid ratios that were used as markers for a food source
3.1 綠刺參食物來源組成
根據特征脂肪酸分析發現, 綠刺參脂肪酸組成中表現出明顯的鞭毛藻及原生動物、綠藻、褐藻、紅藻、異養細菌以及陸源有機質的特征。在7月份時, 鞭毛藻及原生動物在綠刺參的食物來源中占據較大比例。12月份時, 綠刺參食物組成復雜, 包括鞭毛藻及原生動物、異養細菌、褐藻和紅藻等。紅藻、褐藻、陸源有機質等在綠刺參的食物來源中占據的比例與7月份相比較高(P>0.05), 鞭毛藻及原生動物脂肪酸標志則明顯低于7月份(P<0.05)。DHA是鞭毛藻及原生動物類的脂肪酸標志物[32], 同時,也可以指示原生動物[33]等。以前的研究表明[29], 沉積物有機質主要由底棲微藻、原生動物及有機質碎屑組成。綠刺參為沉積食性, 同時其食物來源在不同季節均包括鞭毛藻及原生動物。因此, 綠刺參體內的DHA可能更多的來自沉積物中的原生動物。在12月份時, 根據現場調查[41]的結果, 各類大型藻類會由于水溫的下降變得繁盛。大型藻類產生的大量生物碎屑最終可能被沉積食性的底棲動物(包括海參)攝取[42-43]。因此, 其他食物來源變得更加豐富可能是原生動物對海參的貢獻明顯低于7月份的原因。沿海地區是海洋和陸地環境之間的動態過渡區域。該區域大型底棲無脊椎動物受到陸地來源的影響非常明顯[44]。另一方面, 選擇不同外源性食物的海洋動物會取決于食物來源的可獲得性和自體食物的偏好[45]。陸地有機物的輸入是復雜的, 它不構成海參穩定的食物來源。這可能是綠刺參體內陸源有機質的脂肪酸標志物具有季節差異的原因。
3.2 紅腹海參食物來源組成
在7月份時, 紅腹海參的食物來源顯現出典型的褐藻和紅藻類來源有機物特征。12月份時, 紅腹海參的食物來源組成中異養細菌與綠藻占據較大比例。紅腹海參體內的褐藻脂肪酸標志物20∶4(n-6)的相對含量(12.03%~12.18%)在實驗期間內比其他脂肪酸標志含量較高。綠藻脂肪酸標志[18∶1(n-7)]/[18∶1(n-9)]比值在不同月份均大于1, 表現出典型的綠藻脂肪酸特征。綠藻的生物量在不同季節沒有明顯的變化[41], 可作為海參穩定的食物來源。紅腹海參以多種來源的有機物為食, 包括有機顆粒物和生物碎片, 其中含有大量異養細菌, 它們也是紅腹海參的重要食物來源。
3.3 兩種海參食物來源的異同
紅腹海參的食物來源組成與綠刺參相似, 涵蓋了鞭毛藻及原生動物、異養細菌、褐藻、綠藻、紅藻、陸源有機質等。7月份時, 紅腹海參與綠刺參體內各類食物來源的脂肪酸標志的含量或比值均無明顯的種間差異(P>0.05), 但根據主成分分析發現, 綠刺參是以沉積物中的鞭毛藻及原生動物為主要食物來源,紅腹海參則是以褐藻及紅藻有機質碎屑為重要的食物來源。紅腹海參與綠刺參體內的紅藻、褐藻、陸源有機質等脂肪酸標志的含量或比值均是12月份高于7月份, 鞭毛藻及原生動物則顯著低于7月份。12月份時, 綠刺參的食譜較為豐富, 以鞭毛藻及原生動物、異養細菌、褐藻和紅藻等有機質來源為主要食物來源, 紅腹海參則主要為異養細菌和綠藻。根據Li等人[41]對蜈支洲島海域藻類調查, 結果顯示: 在實驗區有大量的綠藻、褐藻以及紅藻鑲嵌分布。同時, 紅藻類是實驗海域內生物量及種類最為豐富的藻類,主要包括紫杉狀海門冬(Asparagopsis Taxiformi)、巢沙菜(Hypnea pannosa)、矮型石葉藻(Lithophyllum pygmaeum)等。褐藻類為低溫類群, 在冬春季水溫較低時繁盛, 其中主要藻類包括加勒比海褐藻(Lobophora variegata), 半葉馬尾藻(Sargassum hemiphyllum)等。由于沉積食性生物可以通過攝食大型藻類脫落碎屑獲得有機質來源, 因此, 實驗海域內大型藻類產生的有機質碎屑很有可能是綠刺參的重要食物來源組分, 紅腹海參很可能也是通過這種方式獲得了較多的藻類食物來源。異養細菌也是海參的重要食物來源[18,46]。Moriarty等[47]認為綠刺參和黑海參(Holothuria atra)對細菌的同化效率(32%~44%)要高于沉積物中的有機質, 同時, 其結果也表明海參腸道中較高的胞壁酸值是由于海參消化了大量細菌, 而不是由于攝入較多的底棲微藻。
根據實驗結果, 在不同月份中2種海參個體間存在的脂肪酸標志含量差異, 可能與熱帶海參在自然生境中活動范圍較小, 常呈現不均勻或斑塊分布[48-49],不同個體生存區域提供的餌料會有一定的差異有關。因此, 所采海參不同個體間食物來源的可獲得性會有所不同, 導致海參的體內脂肪酸標志含量存在個體差異。
盡管脂肪酸標志法已廣泛運用到海洋營養關系的研究中[22], 但該方法仍是無法對海洋生物的食物來源進行精確地定量定性, 只能對可能食物來源進行示蹤。為改進或彌補這些缺點, 一方面, 可以根據實驗區域內野外調查的結果, 合理地選擇脂肪酸標志物, 并根據特定生物特定時期的代謝特點選擇合適的組織進行脂肪酸組成分析[50]。另一方面, 可以聯合運用脂肪酸標志法和DNA條形碼技術[51]或穩定同位素技術[52]提升對食物來源分析的精度。
4 結論
在自然條件下, 鞭毛藻及原生動物、褐藻、紅藻、大型綠藻、異養細菌, 以及陸源有機質等均是紅腹海參與綠刺參的潛在食物來源。但不同季節, 2種海參的主要食物來源存在差異。7月份時, 鞭毛藻及原生動物在綠刺參的食物來源組成中占據較大比例;12月份, 綠刺參食物組成復雜, 主要以鞭毛藻及原生動物、異養細菌、褐藻和紅藻等為食物來源。大型藻類是紅腹海參在不同季節的主要食物來源。7月份, 紅腹海參的主要食物來源組分為紅藻和褐藻,異養細菌和綠藻是12月份時紅腹海參食物組成中主要組分。本研究結果可為揭示海參攝食活動產生的生態功能提供數據支持, 以及為熱帶海參經濟種的大規模底播增養殖提供參考。
