miR-376b-3p對HepG2細胞Ⅱ相代謝酶UGT2B7的調控
2021-10-16 14:10:06江澤斌鄭付春
汕頭大學醫學院學報 2021年3期
李 敏,江澤斌,鄭付春
(汕頭大學醫學院第一附屬醫院,廣東 汕頭 515041)
微RNA(miRNA)是一類內源性的非編碼小RNA分子,也是最大的基因家族之一[1]。1993年,第一個miRNA lin-4被發現參與線蟲發育的調控[2],隨后的20年間,成千上萬種miRNAs在動植物以及人體內被相繼發現。miRNA的產生主要分為兩個階段:miRNA初級轉錄產物在細胞核中被核糖核酸酶Drosha處理成一個約70個核苷酸長度的前體mRNA;之后在轉運蛋白exportin-5的作用下被轉移到胞質內,進而被Dicer酶切割成長約22個核苷酸的成熟miRNA。成熟miRNA可以與細胞內的Argonaute蛋白等結合形成miRNA誘導的沉默復合體,與靶基因的3′-UTR區域特異性結合,引起靶基因mRNA的降解或翻譯抑制。人體中的miRNAs參與約30%基因的表達調控,與整個生命活動密切相關[3]。多個miRNA能同時調控同一個代謝酶,一個miRNA也能同時調控多種代謝酶。尿苷二磷酸葡萄糖醛酸轉移酶(uridine diphosphate-glucuronyl transferase,UGT)是一類重要的Ⅱ相代謝酶,可以在內源物和外源物分子上加入一個葡萄糖醛酸基團,增加極性,利于其經尿液與膽汁排泄,從而調節體內穩態。UGT2B7是UGT家族中重要的一員。本課題組前期研究發現,miR-376b-3p與癲癇患者丙戊酸鈉血藥濃度有相關性,從丙戊酸鈉的代謝酶中篩選出UGT2B7能被miR-376b-3p調控,且UGT2B7在丙戊酸鈉的主要代謝途徑中所占比例最高。本研究探討miR-376b-3p對HepG2細胞UGT2B7的調控作用。
1 材料與方法
1.1 主要材料與儀器
HepG2細胞(廣州賽庫生物技術有限公司),HEK293T細胞由牛永東老師饋贈,miR-376b-3p模擬物及陰性對照、miR-376b-3p抑制劑及陰性對照(上海誠鑫生物科技有限公司),UGT2B7抗體(武漢三鷹生物技術有限公司),GAPDH抗體(武漢博士德生物工程有限公司),qPCR試劑盒(寶日醫生物技術有限公司),雙熒光素酶報告基因檢測試劑盒(廣州致邦生物科技有限公司),UGT2B7 3′-UTR野生型(WT)以及突變體(Mut)質粒、miR-376b-3p質粒、海腎熒光素酶報告基因質粒(上海吉凱基因科技有限公司),DMEM、胎牛血清、胰酶、青霉素—鏈霉素雙抗溶液(美國Hyclone公司),ABI QS5熒光定量PCR儀(美國ABI公司),報告基因檢測儀(美國PROMEGA公司)。
1.2 方法
1.2.1 雙熒光素酶報告基因分析實驗分組為miR-376b-3p陰性對照+UGT2B7 3′-UTR WT組、miR-376b-3p+UGT2B7 3′-UTR WT 組、miR-376b-3p陰性對照+UGT2B7 3′-UTR Mut組、miR-376b-3p+UGT2B7 3′-UTR Mut組、miR-376b-3p陰性對照+UGT2B7 3′-UTR WT陰性對照組、miR-376b-3p+UGT2B7 3′-UTR WT陰性對照組(WT陰性對照也為Mut陰性對照)。將融合度為70%~80%的HEK293T細胞用胰酶消化后進行細胞計數,按2×104個細胞/孔接種于白色96孔板,培養16~24 h后;每孔轉染25 ng WT/Mut質粒,5 ng海腎熒光素酶報告基因質粒,100 ng miR-376b-3p質粒或miR-376b-3p陰性對照質粒;轉染48 h后,使用雙熒光素酶報告基因檢測試劑盒分別檢測螢火蟲熒光素的熒光強度和海腎熒光素的熒光強度,以螢火蟲與海腎熒光素熒光強度的比值作為結果進行統計。
1.2.2 實時PCR檢測HepG2細胞中UGT2B7的mRNA水平HepG2細胞置于含10%胎牛血清的DMEM培養基中,在37℃、5%CO2的恒溫培養箱中進行傳代培養。使用miR-376b-3p模擬物或miR-376b-3p抑制劑以及陰性對照(30 nmol/L)與5 μL Lipofectamine RNAiMAX轉染試劑形成脂質體復合物,再用無抗生素的培養基稀釋細胞(1×105~3×105個/mL)加入六孔板中,培養36 h后,使用Trizol裂解液進行總RNA提取。PCR反應條件:95℃,30 s;95℃,5 s;60℃,34 s,40個循環,以β-actin作為內參基因檢測UGT2B7 mRNA的表達水平。
1.2.3 蛋白印跡法檢測HepG2細胞中UGT2B7的蛋白水平在六孔板中轉染miR-376b-3p模擬物或miR-376b-3p抑制劑以及陰性對照48 h后,使用PBS緩沖液洗3次,每孔加入50 μL裂解液,冰上裂解30 min,隨后12 000 r/min,4℃離心,收集上清液。采用BCA法蛋白定量,完成后將蛋白在沸水中變性5 min,冷卻后待使用。將蛋白樣本進行SDS-PAGE凝膠電泳,采用12%的分離膠,上樣量10 μg,電泳90 min,隨后采用濕轉法將凝膠上的蛋白轉移至硝酸纖維素膜,轉膜時間為90 min,再將條帶置于5%脫脂牛奶中封閉1 h,封閉結束后在4℃條件下孵育一抗過夜(UGT2B7抗體稀釋比例為1∶2 000,GAPDH抗體稀釋比例為1∶5 000);第2天在室溫下孵育二抗1 h(UGT2B7 1∶40 000稀釋,GAPDH 1∶100 000稀釋),加入發光液,在暗室中曝光,Gel-pro圖像分析軟件分析蛋白條帶的灰度值。
1.3 統計學分析
應用GraphPad Prism 6軟件進行分析,符合正態分布的計量資料以xˉ±s表示,2組間比較采用配對t檢驗,多組間比較采用單因素方差分析,以P<0.05為差異有統計學意義。
2 結果
2.1 miR-376b-3p與UGT2B7 mRNA的結合
結合位點如圖1A所示,miR-376b-3p與UGT2B7 3′-UTR的結合位點具有廣泛的種子配對且第1位核苷酸是A,在第16~20位也有結合。雙熒光素酶質粒突變位點如圖1B所示。在HEK293T細胞中通過雙熒光素酶報告基因實驗驗證miR-376b-3p能否與UGT2B7 3′-UTR直接結合。如圖1C所示,與miR-376b-3p陰性對照組相比,共轉染miR-376b-3p與UGT2B7 3′-UTR WT組的熒光素酶活性降低13%,差異具有統計學意義(P<0.05),共轉染 UGT2B7 3′-UTR Mut組的熒光素酶活性差異無統計學意義(P>0.05)。
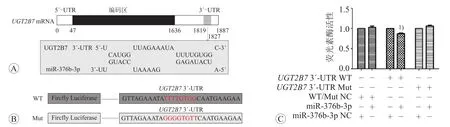
圖1 miR-376b-3p與UGT2B7 mRNA的結合
2.2 miR-376b-3p對UGT2B7 mRNA的調控
如圖2所示,與陰性對照組相比,miR-376b-3p模擬物組UGT2B7 mRNA水平下降44%,miR-376b-3p抑制劑組UGT2B7 mRNA水平升高41%,差異均具有統計學意義(P<0.05)。

圖2 miR-376b-3p模擬物和抑制劑對UGT2B7 mRNA表達的影響
2.3 miR-376b-3p對UGT2B7蛋白表達的調控
如圖3和圖4所示,與陰性對照組相比,miR-376b-3p模擬物組UGT2B7蛋白表達下降34%,miR-376b-3p抑制劑組蛋白表達升高67%,差異均具有統計學意義(P<0.05)。肝藥酶抑制劑雙氯芬酸和肝藥酶誘導劑利福平作為免疫印跡法的陽性藥。

圖3 miR-376b-3p模擬物對UGT2B7蛋白表達的影響
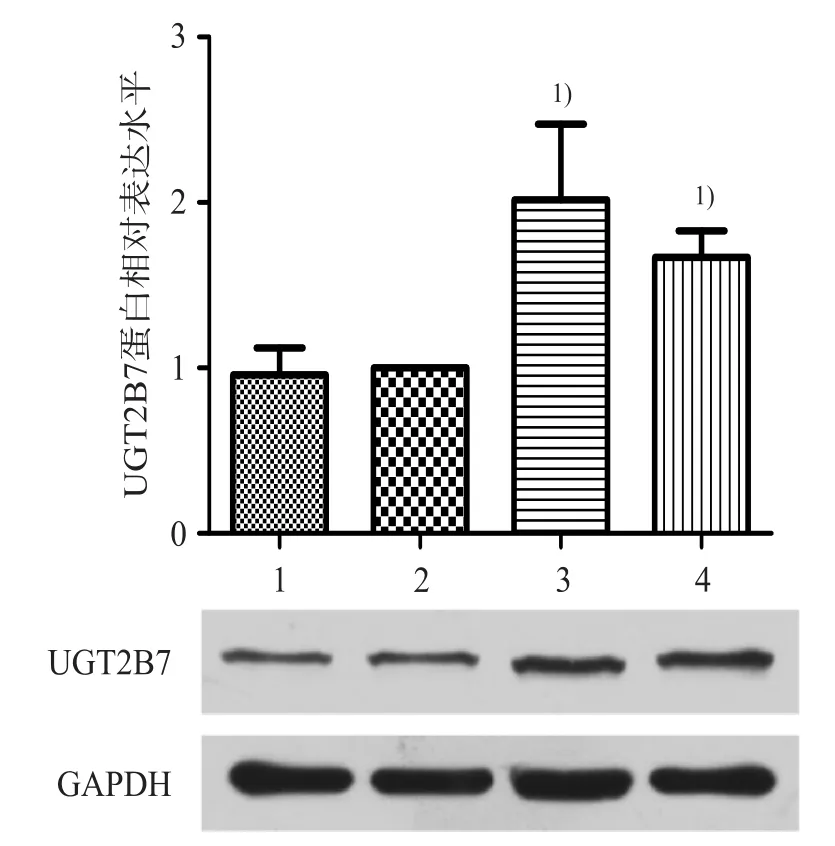
圖4 miR-376b-3p抑制劑對UGT2B7蛋白表達的影響
3 討論
UGT廣泛分布在肝臟、腎臟、心臟、腦、皮膚等器官和組織中,其中在肝臟中活性最高,是生物體內外源性物質進行Ⅱ相生物轉化主要的代謝酶。UGT參與約35%臨床用藥的代謝,35%被UGT代謝的臨床用藥是UGT2B7的底物[4]。UGT2B7在肝臟中高表達,對各種內源性化合物包括膽汁酸、視黃酸、類固醇、鹽皮質激素、糖皮質激素、脂肪酸,以及外源性化合物包括致癌物質,治療藥物如嗎啡、可待因[5]、鹽酸表柔比星[6]、丙戊酸[7],及非甾體類抗炎藥具有高活性。迄今為止,人們已經利用肝癌細胞系、人原代肝細胞和轉基因小鼠等肝臟模型對UGT2B7的轉錄調控進行了深入研究[8]。已發現參與調控UGT2B7表達的轉錄因子包括肝細胞的核因子1α[9]、HNF4α、尾相關同源框因子2[10]、紅細胞來源核轉錄因子Nrf2、核內X受體[11]、p53[12]和激活蛋白1[13]。隨著對miRNA研究的不斷深入,已經證實miRNA不但在生理過程中發揮重要作用,而且在腫瘤發生和藥物代謝過程中也發揮重要的調控作用。miRNA可以直接調控藥物代謝酶UGT2B7的表達,也可以通過調控轉錄因子而間接調控UGT2B7的表達。盡管UGT2B7的轉錄因子調控機制已得到充分研究,但它如何被miRNA調控還鮮有報道。
miRNA通常通過mRNA降解或翻譯抑制來抑制基因表達,具體作用機制取決于miRNA和靶mRNA之間序列互補的程度和性質。本研究miR-376b-3p與UGT2B7 3′-UTR的結合位點具有廣泛的種子配對且第一位核苷酸是A,在第16~20位也有結合,這更有利于向mRNA降解途徑發展。本研究利用雙熒光素酶表明miR-376b-3p通過UGT2B7 3′-UTR的一個結合位點對UGT2B7進行負調控。HepG2細胞癌性較低,有較完整的Ⅰ、Ⅱ相代謝酶的表達。本研究在HepG2細胞中轉染miR-376b-3p模擬物,UGT2B7 mRNA水平降低44%,轉染miR-376b-3p抑制劑,UGT2B7 mRNA水平升高41%,在蛋白水平上,轉染miR-376b-3p模擬物使UGT2B7蛋白水平下降34%,轉染miR-376b-3p抑制劑能使UGT2B7蛋白水平升高67%。這表明miR-376b-3p在UGT2B7 mRNA水平和蛋白表達水平上都有抑制作用。
綜上所述,miR-376b-3p能通過結合UGT2B7 3′-UTR負調控UGT2B7的mRNA和蛋白表達。miR-376b-3p可能通過調控UGT2B7,進而影響丙戊酸鈉的血藥濃度,這或許是丙戊酸鈉個體差異性較大的部分原因。
