合成生物學研究中的微生物啟動子工程策略
2021-09-24 01:19:34于慧敏鄭煜堃杜巖王苗苗梁有向
合成生物學 2021年4期
于慧敏,鄭煜堃,杜巖,王苗苗,梁有向
(清華大學化學工程系,教育部工業生物催化重點實驗室,北京 100084)
近年來,致力于對生物學元器件進行標準化設計構建,使其未來可以在新型人工生物系統中進行常規重復使用的合成生物學研究在全球范圍內方興未艾[1-2]。利用合成生物學理論與工具,對生物體尤其是微生物及其相關蛋白(酶)、基因元件與線路等,進行目標明確的設計、重組改造乃至重新合成,對于解決我國與國計民生相關的生物醫藥、材料、環境、能源等重大問題,實現綠色制造和可持續發展、實現大健康戰略具有重要的理論意義和實踐價值。
在合成生物學研究中,基因的精細表達調控是決定研究進程的關鍵因素;而對于原核微生物,影響基因表達水平的最重要因素則是決定RNA 聚合酶(RNA polymerase,RNAP)轉錄機器識別效率的基因啟動子序列。不同種屬微生物中的不同啟動子序列決定了下游基因的表達強度及表達模式(組成型、誘導型),因此成為驅動基因表達、調控基因線路、賦能微生物功能、獲得高產酶與高性能細胞工廠的合成生物學核心元件[3]。通過啟動子工程策略對微生物啟動子特征及功能、全新的基因誘導表達模式進行發掘與進化改造,使其響應不同的物理、化學信號,實現精細、動態、按需調控目標基因(群)表達,已經成為目前合成生物學的研究熱點。
1 啟動子識別與基因轉錄調控的一般規律
啟動子是一段位于目標基因轉錄區上游、能夠與RNA 聚合酶結合從而實現轉錄起始的DNA 序列。在原核微生物中,負責執行轉錄功能的轉錄機器——RNA 聚合酶全酶(包含α2、β、β'、ω、σ等亞基)依賴于其中的σ因子,特異性地識別具有特定序列特征的啟動子區域并與之結合,從而引發轉錄的開始[4]。為了更好地闡明后文針對啟動子工程研究的各種策略與前沿研究的設計思想,有必要從啟動子識別與轉錄調控的基本規律說起。
以大腸桿菌為例,其σ 因子有7 種,分別為:σ70(σD)、σ54(σN)、σ38(σS)、σ32(σH)、σ28(σF)、σ24(σE)和σfecI[5]。其中,σ70被稱為看家σ 因 子(housekeeping σ factor),它負責起始細胞生長相關的高達1 000多個基因的轉錄;σ54、σ38與σ32分別調控細胞的N 代謝基因、細胞穩定期相關基因以及熱應激響應基因的轉錄。其他σ因子則分別調控極端熱應激響應及外細胞質基因、鞭毛以及檸檬酸鐵代謝相關等基因[6]。不同的σ因子,對應識別不同的目標基因啟動子。
狹義的啟動子通常是指在轉錄起始位點(+1)上游大約10 個及35 個核苷酸(nt)處的兩段DNA短序列(也稱為?10 區與?35 區;或啟動子核心區),它們對于不同σ 因子的識別具有決定性的作用[7]。以大腸桿菌啟動子為例,對所有σ70因子識別基因啟動子區域的統計分析表明,?10 區與?35區的一致序列為TATAAT 與TTGACA。這兩段序列之間通常被17 nt 的間隔序列所隔開。17 nt 的間隔序列長度對于大腸桿菌基因的轉錄水平十分重要。當間隔區長度長于18 nt或短于16 nt時,RNA聚合酶與啟動子的親和性都會顯著降低,從而調低基因轉錄[7]。此外,有些?35 區的上游序列能夠與RNA 聚合酶的α 亞基結合從而影響轉錄強度——這些序列被稱作UP元件(UP element)[6,8]。
以大腸桿菌典型的啟動子結構為例,其Plac和Ptrp啟動子、σ70因子識別保守序列、σ54與σ32因子識別保守序列特征如圖1所示[6-11]。

圖1 大腸桿菌代表性啟動子及幾種σ因子的識別保守序列Fig.1 Representative promoters in E.coli and conserved sequences for RNA polymerase sigma factors
再以另一種模式微生物——枯草芽孢桿菌為例,其看家σ 因子識別的啟動子的?10 區與?35 區保守序列以及優選的間隔區長度都與大腸桿菌相同[8-9]。但與大腸桿菌不同的是,枯草芽孢桿菌的啟動子還存在一些統計學特殊規律,比如,其?48 位大多為保守的T 堿基,?43 位附近富含A 堿基,而?17~?14 位的特征序列則通常為TNTG 等。在其他一些革蘭氏陽性菌中該規律也存在。此外,枯草芽孢桿菌中還存在特定的、負責控制與芽孢形成相關基因表達的轉錄因子σG,其靶基因的保守?10 區與?35 區一致序列變更為CATACTA 與TGAATA,優選的間隔區長度可為17~18 nt[10-11]。
需要進一步強調的是,對于不同的非模式微生物,由于其看家σ 因子(以及其他σ 因子)可識別的啟動子序列經常并不相同,這就決定了在不同微生物種屬之間,乃至同一菌屬但不同種的微生物之間,很多啟動子都無法通用,或者其轉錄啟動效果在不同菌種中呈現顯著差異。表1列出了幾種典型模式微生物與非模式微生物的σ70因子偏愛的?35 與?10 核心區序列。由表可見,盡管大腸桿菌與枯草芽孢桿菌的核心區保守序列具有相同的序列特征,但其他幾種微生物的核心區序列均存在一個或多個堿基的差異。

表1 幾種模式/非模式微生物看家σ因子識別的啟動子保守序列比較Tab.1 Some promoter consensus sequences for housekeep?ing σ factors of model/non-model microorganisms
上述不同σ因子特異識別的保守序列差異與微生物細胞內成百上千不同基因的啟動子序列差異,就導致不同微生物基因在表達時可以形成各自的表達時間模式與強度模式,這些表達模式對于合成生物學研究至關重要。因此,針對不同微生物尤其是重要的非模式工業微生物,其σ 因子種類、數量與特征、受不同σ因子識別調控從而起始轉錄過程的新型啟動子的特征、功能與調控規律研究正在受到越來越多的關注。
此外,按作用強度區分,啟動子可以分為強啟動子(strong promoter)、中等強度啟動子(moderate promoter)和弱啟動子(weak promoter),其實質就是其上述核心區特征序列與RNA 聚合酶σ 因子的特異性結合能力存在差異。通常,強啟動子和中強啟動子常用于目標基因的高表達,而弱啟動子則用于特定基因的弱化表達,以調控細胞內物質和能量的代謝平衡[17]。
按作用方式區分,啟動子又可以分為組成型啟動子(constitutive promoter)和誘導型啟動子(inducible promoter)兩大類。其中,組成型啟動子調控的基因,不需要轉錄調控因子的參與,在細胞生長過程中即可實現基因的正常轉錄與翻譯表達。而誘導型啟動子,則必須要正調控或負調控蛋白與誘導物(外加化學試劑或代謝物分子等化學信號或溫度、光、金屬離子等物理信號)組合的共同作用,才能啟動轉錄過程的順利進行。鑒于合成生物學研究需要對細胞工廠中錯綜復雜的代謝途徑與多目標基因進行差異化精準調控,誘導型啟動子受到更多的關注,現有誘導型啟動子的按需進化改造、新型誘導型啟動子的發現、誘導機制解析以及新誘導方式的普適化拓展應用等已經成為合成生物學領域的研究前沿與熱點。
誘導型啟動子的負調控與正調控機理以及大腸桿菌最典型的β-半乳糖苷酶基因(lacZ)的負調控誘導型啟動子的典型結構特征如圖2所示[6]。
在啟動子的負調控方式中,負調控蛋白(也稱作阻遏蛋白或抑制子等)對目標基因的轉錄起負面的阻遏作用;它通常會結合到啟動子下游、目標基因上游的一段被稱為操縱序列的小DNA 片段上,從而阻止RNA 聚合酶對目標基因的正常轉錄。而當添加外源誘導劑后,誘導劑與負調控蛋白具有更強的結合作用,使其從操縱序列上脫離,繼而基因的正常轉錄被開啟[圖2(a)、(b)]。在啟動子的正調控模式中,情況則恰好相反。調控蛋白對于目標基因的轉錄起正向的激活或促進作用(因此有時也稱作activator)。只有當誘導劑加入時,誘導劑與調控蛋白結合后的復合物才能夠結合到操縱序列上,激活或促進下游目標基因的轉錄。
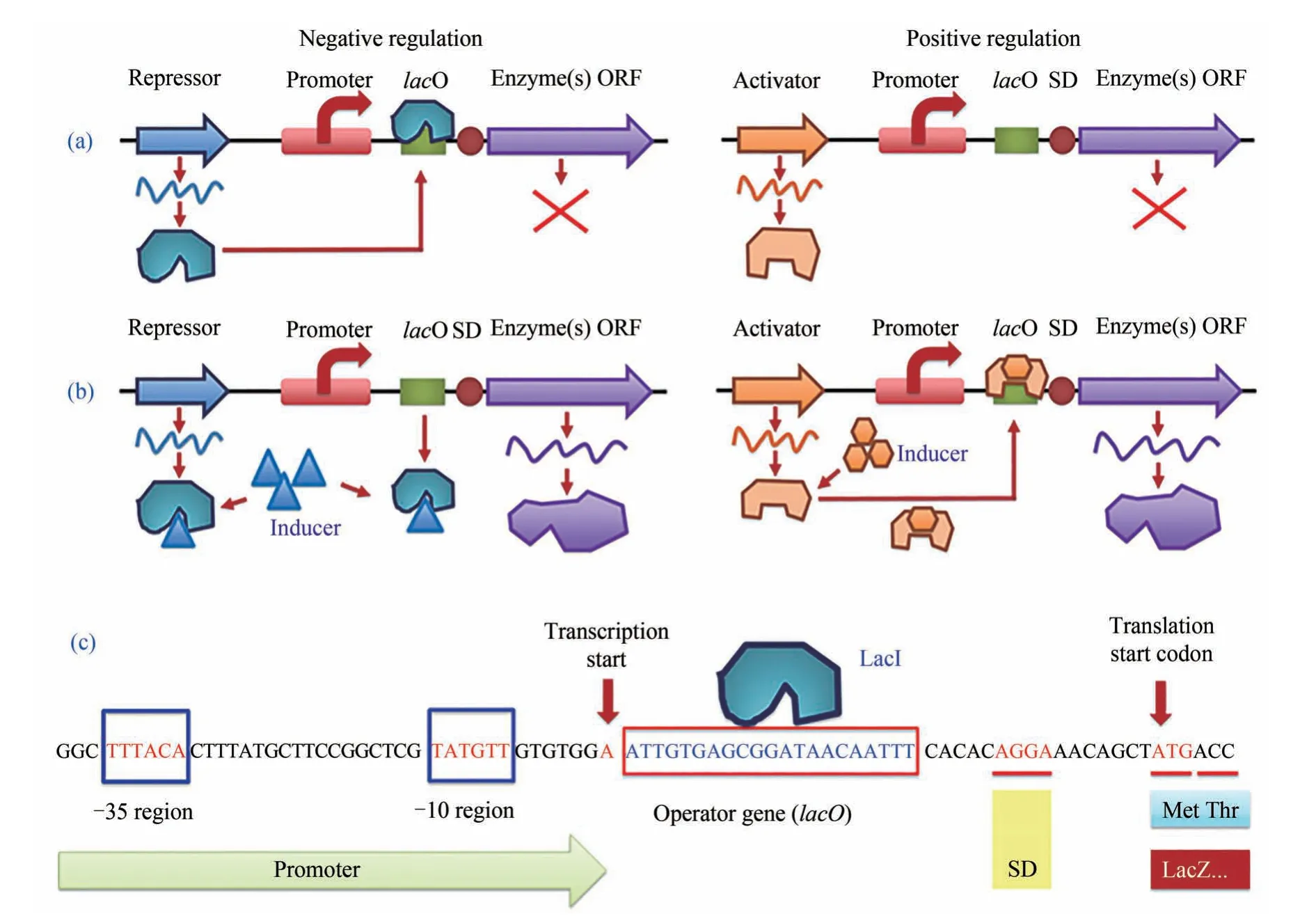
圖2 誘導型啟動子的負調控與正調控誘導機制以及大腸桿菌乳糖操縱子負調控的典型序列(a)無誘導劑條件下的基因調控;(b)誘導劑加入條件下的基因調控;(c) β-半乳糖苷酶(LacZ)啟動子的乳糖操縱子負調控區域DNA序列。LacI—阻遏蛋白;lacO—操縱序列;SD序列—基因的核糖體結合位點(也稱為RBS);Inducer—乳糖操縱子的lacI-lacO體系誘導劑,通常為異丙基-β-D-硫代吡喃半乳糖苷(IPTG)或乳糖(實際為1,6-別乳糖,乳糖的代謝物)Fig.2 Negative and positive regulation mechanism of inducible promoters and the partial sequence of lactose operon(lacO)of E.coli.(a,b)gene regulation mode under inducer-free or inducer-present condition,respectively.(c)partial DNA sequence of inducible promoter of β-galactosidase(LacZ)with negative regulation.LacI—inhibitor(repressor)protein;lacO—operator sequence for repressor binding;SD—Shine-Dalgarno sequence as ribosome biding site(RBS);Inducer—for lacI-lacO system,common inducer is isopropyl-β-Dthiogalactopyranoside(IPTG)or lactose(1,6-allolactose in fact,a metabolite of lactose),respectively
除了上述的正調控機制和負調控機制分別單獨發揮作用的情況外,原核生物中也有許多誘導型啟動子的調控作用需要正調控與負調控因子共同作用完成。其中,有的調控機制只需要2個調控因子合作,而有的則需要多個正負調控因子一起協作完成。
2 啟動子改造的常用策略
為了實現關鍵目標基因的優化表達,經常需要對核心啟動子進行遺傳改造,以實現基因轉錄水平的可控調節乃至表達強度的精細調控。一般采用如下兩種策略:一是對靶基因自身的內源啟動子進行突變改造,或者將啟動子與特定的轉錄因子相結合,從而改變啟動子的強度[18];另一種方式則是將原有的啟動子替換成其他啟動子,從而徹底改變受控基因的表達譜,實現對關鍵基因轉錄水平的人工控制[19]。如圖3所示。
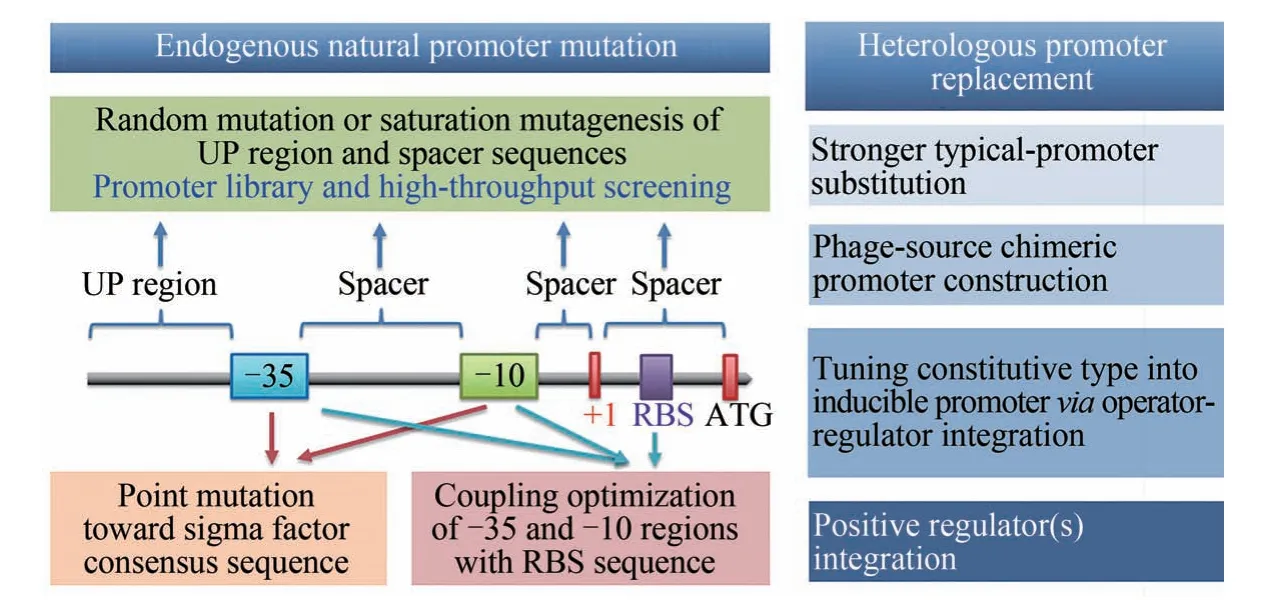
圖3 啟動子改造的常用有效策略?35和?10—啟動子核心區;+1—轉錄起始位點;RBS—核糖體結合位點;ATG—翻譯起始密碼子Fig.3 Common efficient strategies for promoter engineering?35 and ?10—core elements of promoter;+1—transcription initiation site;RBS—ribosome binding site;ATG—start codon of enzyme translation
在啟動子突變方面,常用方法包括隨機突變與定點突變。例如,可以通過易錯PCR、定點飽和突變乃至創制合成等方式對啟動子的UP 區或間隔區進行隨機突變,創制不同強度的啟動子庫,或者創制合成啟動子庫(synthetic promoter libraries,SPL),用于代謝工程中多基因的差異表達調控;再結合高通量篩選方法獲得優選的啟動子[19-20]。隨機突變方法的優點是能夠得到不同性質及強度存在梯度分布的啟動子;其缺點則是工作量大,需要結合合適的高通量篩選方法才能實現。也可以將目標啟動子的核心?10 區、?35 區等關鍵區域序列向著σ因子識別一致序列的方向突變,從而顯著改變啟動子的活性[14,21-22]。例如,Jiao等通過對枯草芽孢桿菌Pg2 啟動子的核心?10 區、?35 區分別進行面向σ因子識別保守序列的單點突變,突變啟動子的強度提高了15 倍[21];在紅色紅球菌啟動子的研究中,同樣獲得了類似的研究結果[14]。
在啟動子的替換策略中,可供選用的啟動子既可以是自身或者一些種屬相近微生物中已被報道的或被廣泛使用的啟動子(如大腸桿菌中普遍使用的Ptac強啟動子以及枯草芽孢桿菌中常用的P43 啟動子等);也可以通過報告基因表達的方式對一組候選啟動子的強度進行對比分析,從而得到可以使用的優選啟動子[23-24];還可以選用一些噬菌體啟動子、人工構建的雜合啟動子等外源啟動子。其中,優選的噬菌體啟動子,例如來自T7 噬菌體的T7啟動子、來源于噬菌體SPO-1的Pspac啟動子[25],往往具有強轉錄特性。2013 年,Yang等[26]進一步從B.licheniformis基因組中篩選獲得了PluxS 啟動子,活性是P43 啟動子的8 倍。將PluxS 與枯草芽孢桿菌Papr的?10 區結合,構建的雜合啟動子PlapS,具有更高的轉錄活性。尤其是這些外源啟動子不僅可以提高目標基因的轉錄強度,還可以利用大腸桿菌負調控蛋白-操縱子序列(LacI-lacO)等的引入,成功實現人工強啟動子從組成型到誘導型的轉變。例如,LacI-T7 系統誘導型雜合啟動子插入枯草芽孢桿菌基因組,能夠實現高達10 000 倍的啟動強度動態調控范圍[27]。將紅球菌Rhodococcus rhodochrousJ1中的腈水解酶誘導型啟動子與NitR正調控蛋白組合成PnitA-NitR雜合元件,成功構建了鏈霉菌高效表達啟動子——以ε-己內酰胺為誘導劑,鏈霉菌蛋白表達量可以達到可溶蛋白的40%[28]。啟動子替換改造的優勢在于針對性強,有時可以大大減少篩選工作量。但另一方面,可供選用的啟動子仍然非常有限,遠不能滿足快速發展的合成生物學研究的需求。
此外,啟動子改造研究也經常與核糖體結合位點RBS的序列優化相結合。例如,通過3條枯草芽孢桿菌內源組成型強啟動子序列與3條優選RBS序列的隨機突變庫,研究人員篩選獲得了基因表達水平達到約14 000 倍動態調控區間的基因工具箱[29]。基于異源σ70依賴的啟動子可以被鏈霉菌的“看家”σhrdB因子有效識別,Zhao等[30]將大腸桿菌啟動子Ptac的核心區與變青鏈霉菌啟動子PkasO*R15 的5′-非翻譯區(5′-UTRs)融合,獲得了改造啟動子Ptac*,其在鏈霉菌S.lividansTK24中的啟動強度達到Ptac的8.1 倍,以及PkasO*R15的1.7倍。進一步耦合RBS優化,優選啟動子-RBS組合Ptac*RBS3對報告基因的表達強度達到Ptac啟動子的17.6倍。
近年來,隨著合成生物學的興起以及組學分析、高通量篩選、人工智能等新興技術的快速發展,啟動子改造的新思路和新方法也不斷涌現。Li 等[31]及Shen 等[32]發現,針對啟動子核心區的間隔序列進行隨機突變或飽和突變,可以獲得具有310倍轉錄活性差異的啟動子庫,從而獲得優選強啟動子并顯著提高目標產物PHB以及P(3HB-co-4HB)的產量。類似地,啟動子上游序列突變對合成啟動子的轉錄強度的影響也不容忽視[33]。而針對鏈霉菌啟動子調控序列進行隨機突變構建不同強度的啟動子庫,也成功篩選獲得了一系列強、中、弱轉錄活性的鏈霉菌啟動子[34]。進一步地,除了啟動子核心區序列、上游序列及間隔序列以外,研究人員還發現,位于SD 序列上游的5′-UTR序列結構對于轉錄及翻譯效率也具有顯著影響;研究表明,相對于PBAD啟動子,大腸桿菌細胞內104 個天然5′-UTR 序列的啟動子強度在轉錄水平上從0.007%到4630%之間變化,而在翻譯水平上則從0.1%到137%之間變化[35]。此外,基于單個啟動子改造研究的突破,串聯的雙啟動子組合策略也受到重視。研究發現,兩個啟動子之間的間隔序列長度對組合啟動子活性的影響很大,其中80 bp 是優選長度[36]。最后,利用組學數據和生物信息學工具進行啟動子序列識別與改造[37]、強度預測與RBS 序列優選[38]并指導新啟動子發現、高強度新組成型或誘導型啟動子改造以及目標基因的高效表達等研究策略,已經成為今后在合成生物學等生物技術領域開展啟動子研究的必要工具。
3 不同類型誘導型啟動子的研究進展
鑒于誘導型啟動子在合成生物學、代謝工程和工業生物發酵中的重要地位,針對誘導型啟動子的專門研究越來越多,啟動子元件在生物技術研究與生產實踐中的應用前景非常廣闊。研究人員不僅關注經典強啟動子的進一步改造和新用途,比如,T7 轉錄系統與T7 噬菌體啟動子在邏輯門與基因線路中的應用[39],還關注各種新型誘導劑的發現(化學分子)以及新誘導方式的開發(物理信號),以滿足日益增長的工程生物學研究的迫切需要。
迄今為止,從不同微生物來源的菌株中,已經發現大量的新類型誘導型啟動子,受不同化學誘導劑分子或物理誘導信號的調控,對目標基因進行不同強度的表達。相關研究進展總結見表2。
由表2可見,化學誘導劑的種類目前已經增加到了幾十種。但在工業生產中真正實現了應用的,還比較少。總體而言,IPTG 類誘導劑仍然最為高效、嚴謹且應用最為廣泛。但IPTG 的高成本問題已經稱為制約其進一步推廣的核心因素。有鑒于此,研究者們致力于開發全新的,兼具高效、嚴謹且低成本優勢的新型誘導型啟動子,目前已經取得較好進展。例如,對異丙苯甲酸酯誘導啟動子、ε-己內酰胺誘導啟動子等。本文作者課題組正在致力于解析紅色紅球菌中獨特的尿素誘導型強啟動子的誘導機理,一旦獲得成功,有望推廣到其他宿主菌株中獲得普適性應用。而營養源饑餓誘導型啟動子(如氮饑餓、磷酸鹽饑餓)、低溶氧誘導啟動子等,也具有工業化的應用潛力與優勢。
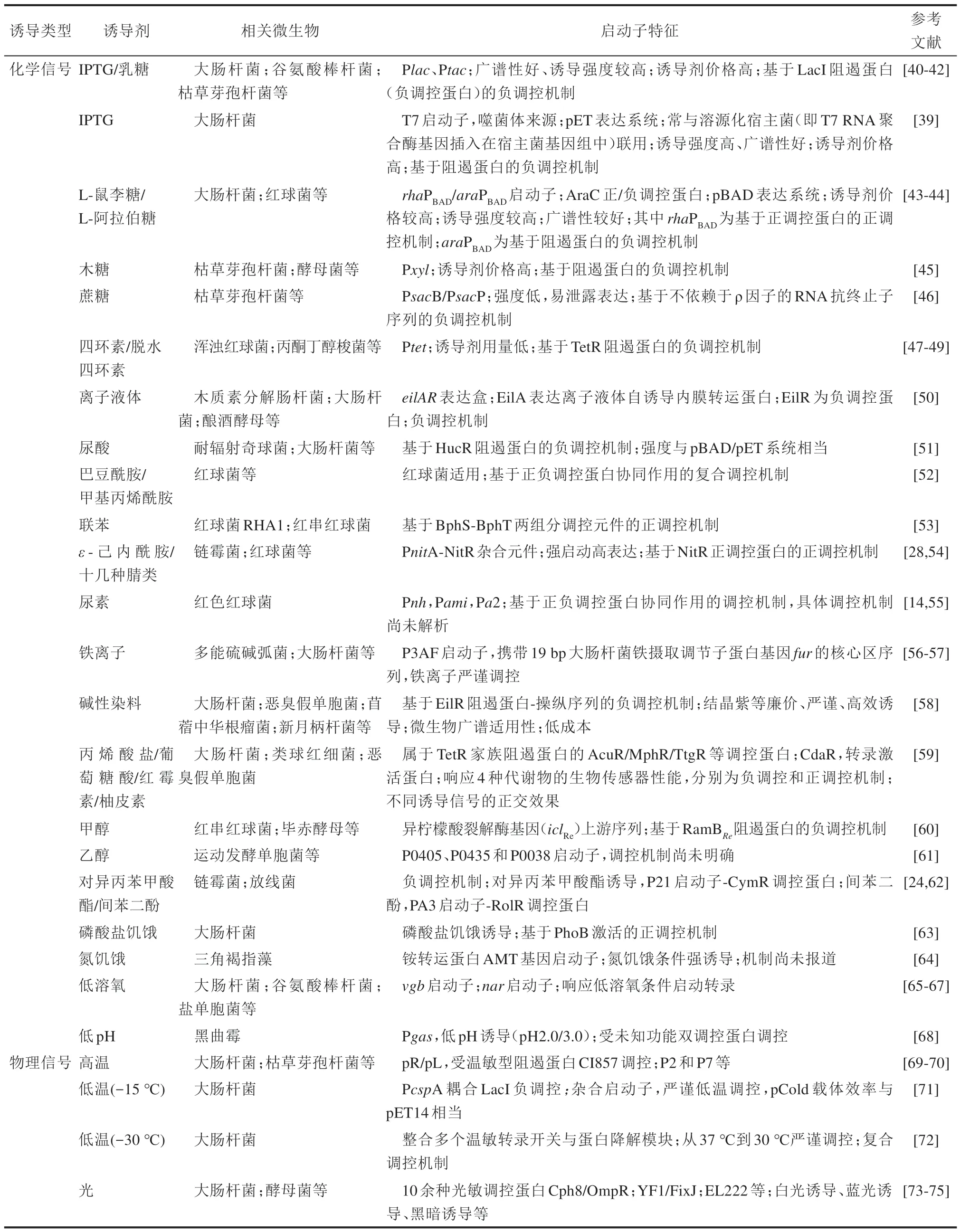
表2 近年來報道的一些不同微生物來源、不同類型的誘導型啟動子Tab.2 Some inducible promoters reported recently from different microorganisms with diverse mechanisms
物理信號誘導的啟動子也吸引了大量研究者的關注。比如,溫度誘導啟動子、光誘導啟動子等。其中,微生物溫度誘導啟動子主要分為高溫(熱)誘導和低溫(冷)誘導兩類,其溫敏開關涵蓋了DNA 型、RNA 型以及蛋白質型[69-72]。而光誘導啟動子則分為白光、藍光、黑暗誘導等多種類型,其調控蛋白等調控因子也各有不同[73-75]。盡管這些啟動子在實驗室規模的研究中已經取得了很多令人振奮的突破,但放大到大規模工業生產水平,變溫操作的能耗問題以及光誘導信號的液面下衰減問題仍然困擾著產業界和學術界。可以預期,新型分子生物技術與新型生物裝備的結合,有望解決上述問題。
4 組成型啟動子的研究進展
相比于誘導型啟動子,組成型啟動子不涉及操縱序列、阻遏蛋白、激活蛋白等轉錄調控元件,因此其工作機制相對簡單,但其在合成生物學、代謝工程等領域高性能細胞工廠以及細胞催化劑的構建研究中同樣具有舉足輕重的作用。就方法學而言,組成型啟動子的改造策略已經在前文中充分討論,在此就不再贅述。
目前,針對組成型啟動子的研究熱點,大致可分為兩類,均是基于目前快速突破的基因組、轉錄組及蛋白質組等組學測序技術以及基于計算機輔助的生物信息學與人工智能技術:一方面是面向基因編輯工具比較匱乏、基因改造相對困難的各種非模式重要工業微生物或特殊微生物,開展不同強度新啟動子、不同表達時序新啟動子的
識別發現及改造研究,乃至不同微生物中RNA 聚合酶σ 因子(或其他重要轉錄元件)的種類、功能、基因轉錄調控規律及其改造方法與應用等研究;另一方面則是面向基因背景清晰、基因編輯工具相對成熟的模式微生物(如大腸桿菌、酵母菌),基于生物信息學及人工智能工具,進行啟動子序列結構基本規律解析、人工預測與設計、人工合成乃至人工智能從頭設計等研究[14,31,76-82]。
例如,Jiao 等[14]針對在生物催化工業中具有重要應用的非模式微生物紅色紅球菌,通過不同生長階段轉錄組的測序分析,獲得了其生長期高表達基因的啟動子序列特征,發現了其σA識別啟動子的保守序列。Li 等[31]則針對嗜鹽菌開展了組成型啟動子庫的研究——從高表達的孔蛋白啟動子出發,針對其核心區間隔序列突變,獲得了梯級強度的組成型啟動子庫,以滿足細胞工廠中大量基因的不同表達強度需要。Trisrivirat 等[78]采用類似的策略構建組成型啟動子庫,實現了生物丙烷產量的提高。Sun 等[79]通過采用生物信息學工具進行RBS 設計與篩選,并耦合啟動子序列改造,獲得了分枝桿菌的高表達元件,同時實現了甾醇產量的顯著提升。Liu等[80]通過對酵母菌中TATA-盒與轉錄起始位點間序列的隨機突變與篩選,獲得了兩個新的強啟動子并提高了β-胡蘿卜素的產量。Zhang 等[81]基于統計模型預測,重構了谷氨酸棒桿菌的人工合成啟動子庫,獲得了優選的串聯啟動子P70,其強度高達tac啟動子的1.21 倍。針對大腸桿菌啟動子,Wang等[82]基于機器學習算法,實現了大腸桿菌啟動子的AI 從頭設計。人工智能、生物信息學與啟動子領域的交叉研究,已經成為啟動子工程的新前沿之一。
5 啟動子工程的新前沿
隨著合成生物學與人工智能工具的快速發展,越來越多的開創性研究在啟動子工程領域涌現。具有動態代謝調控功能的特殊啟動子的發現與改造、新性能啟動子元件的人工智能設計與進化等研究是啟動子工程領域的代表性新前沿。
首先,微生物的胞內代謝網絡十分復雜,各種代謝物以及功能酶的濃度會隨著時間進行動態改變,采用常規的靜態代謝工程策略,如基因的過表達(上調)、敲除或弱化表達(下調)進行代謝流調控,即使在優化調控條件下,也可能造成菌體在某個時間段發生代謝失衡,或者有毒中間代謝產物的積累,不利于菌體生長和目標產物合成,從而導致產物的產量和得率并不理想。動態代謝調控(dynamic regulation)的概念近年來應運而生,其核心思想是讓菌體細胞能夠感應到時時變化的環境狀況或自身的胞內信號,并對其做出響應,調控相關代謝反應以實現代謝途徑的動態平衡,進而達到目標產物合成的高產量、高底物轉化率的統一。動態代謝調控元件的開發是實現動態代謝調控模式的關鍵,它們一方面具有生物傳感器的功能,另一方面也具有執行代謝調控的功能。在目前已經報道的多種元件中,特殊的新模式誘導型啟動子與群體感應信號系統、核糖開關等元件一樣,都是研究人員關注的重點。例如,?ztürk 等[83]闡述了雙啟動表達策略的基本原則,以滿足未來動態代謝工程領域的需求。Landberg 等[84]開發了一種基因組整合型trp?T7 表達系統,可以實現依賴于色氨酸或酵母提取物濃度的自誘導,嚴謹調控目標基因的表達;用該系統生產L-絲氨酸,補料分批發酵培養的絲氨酸產量達到26 g/L。Moreb 等[85]開發了基于E.coliPhoB 調控的磷酸鹽饑餓誘導啟動子,能夠在營養豐富培養基和合成培養基中均實現外源蛋白的自誘導嚴謹調控,使得外源蛋白在磷酸鹽耗盡時自行啟動高表達,從而實現了細胞生長與產物合成解耦的兩階段自誘導發酵培養模式。類似地,Ikegaya 等[86]也報道了來源于紅串紅球菌R.erythropolisN9T-4 的營養匱乏誘導型調控因子。Liang 等[87]則開發了基于產物香草醛以及底物阿魏酸濃度的動態代謝調控回路,實現了香草醛的高產。
其次,人工智能工具已經在越來越多的領域顯示出了強大的功能。在啟動子工程方面,利用機器學習新方法進行人工啟動子設計、創制與優化的研究現已蓬勃興起,并取得了突破性的成果。例如,如前所述,Wang 等[82]發表了基于機器學習的大腸桿菌啟動子的AI 從頭設計框架與活性預測模型,結果表明,經過兩輪優化,70.8%的AI 設計人工啟動子都可以被證明有效,且其序列與大腸桿菌原基因組序列的相似性很低,說明人類已經可以基于機器學習,創造出有效且可能更高效的啟動子新序列。類似的,van Brempt 等[88]則通過卷積神經網絡的機器學習方法,獲得了啟動子序列與功能之間的構效關系信息,成功預測了大腸桿菌σ70啟動子的轉錄起始速率、17 nt 的啟動子間隔區序列,以及枯草芽孢桿菌3 種不同σ 因子特異性的啟動子序列。Zhao 等[89]通過機器學習方法,構建了包含3665 個不同序列的人工啟動子庫,其強度變化范圍達到兩個數量級,且最強啟動子的轉錄活性達到1 mmol/L IPTG 誘導的T7 啟動子的1.52 倍以上。這些研究工作,將成為人工智能工具在啟動子工程領域深度并全面應用的開端。
6 展 望
隨著我國可持續發展戰略的加速推進及二氧化碳減排的迫切需求,綠色生物制造產業正在全面拓展與提升,合成生物學以及面向產業化的工程生物學,已經被認為是本世紀最重要的生物技術平臺[1-2,90]。
在微生物中,轉錄調控是實現目標基因按需表達的最基礎、最重要因素之一[76],啟動子則是轉錄水平調控的核心元件[91-92]。基于啟動子的基因高效及精準表達調控在合成生物學中具有舉足輕重的地位[93-94]。伴隨著合成生物學新技術、高通量生物檢測新裝備、單細胞測序技術、生物信息學乃至人工智能等交叉研究領域的協同發展,啟動子工程已經取得了令人矚目的進展和突破,但同時大量具有挑戰性的科學問題目前還仍然存在。例如,單一啟動子經常發生的基因泄露表達及干擾問題,尚缺少根本性的解決方案;大量組成型與誘導型啟動子的強度響應范圍仍然不能滿足合成生物學研究與產業化生產的需要;眾多有價值的非模式微生物菌株的基本轉錄調控元件、工作原理與調控規律還不十分清楚(比如其看家σ因子的啟動子識別保守序列、調控生長穩定期基因表達及各種環境響應性基因表達的新σ因子及其保守序列、新型正負調控蛋白等);響應更多化學分子與物理信號的新誘導型啟動子的發現、誘導機制解析與改造應用還具有非常廣闊的研究空間;從產業角度,具有安全、高效、低成本特征的新型誘導劑的開發及其產業化應用仍然非常有限。此外,在不同微生物種屬之間具有廣譜通用性的啟動子還不多見;多啟動子串聯協同強化轉錄的研究還剛剛起步;啟動子與反義RNA、核糖開關、群體感應信號等元件協同實現代謝途徑重構優化、基因線路設計構建、新型生物傳感器設計開發、人工多細胞體系以及無細胞體系高效合成目標化學品等研究方興未艾;人工智能從頭設計啟動子的開創性研究還受限于菌種基因背景清晰度、基因編輯工具可用性、高效性以及新算法的性能與精度以及軟件開發等問題。總之,啟動子元件的科學基礎與基因轉錄調控規律的深入研究、其改造方法與手段的全新突破、各種新型誘導劑分子與誘導模式的創制及產業化復雜環境下的高效且低成本應用乃至其調控動態化、設計智能化以及創造性從頭設計的新發展,都將是今后一段時間內微生物啟動子工程領域的重要研究內容[82,88-89,94-100]。
隨著合成生物學的研究對象逐漸從基因元件與模塊走向全細胞復雜代謝線路從頭設計乃至生物新功能從頭創制,可以預見,在如下合成生物學領域,微生物啟動子工程都將發揮越來越重要的作用,包括:新酶的高表達與高效制備;全細胞生物催化;基因線路設計、動態代謝調控以及高性能細胞工廠創建與優化;高靈敏度生物檢測及多功能無細胞合成體系開發;高效、穩定的人工多細胞體系創制及復雜環境應用以及各種新興交叉領域,如生物-金屬復合催化、生物新材料、生物系統自動控制、生物信息學與AI新工具等等。
綜上所述,啟動子工程,尤其是微生物啟動子工程,將驅動合成生物學研究與綠色生物制造技術的持續快速發展,從而為我國綠色化工、環境保護、醫藥與大健康、食品及能源等領域的發展做出不可替代的重要貢獻。
致謝:謹以此文致敬Daniel I.C.Wang 教授在基因工程及生化工程等領域的開創性貢獻。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
現代畜牧科技(2021年4期)2021-07-21 06:13:00
科技傳播(2019年22期)2020-01-14 03:06:54
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
數學物理學報(2017年2期)2017-06-05 09:12:30
