敏感型和耐藥型HIV毒株感染模型的全局動(dòng)力學(xué)
2021-09-13 14:13:30滕志東
陳 偉, 滕志東
(新疆大學(xué) 數(shù)學(xué)與系統(tǒng)科學(xué)學(xué)院,新疆 烏魯木齊830046)
眾所周知,HIV病毒侵入人體后,人體的免疫功能遭到破壞,最終導(dǎo)致人體死亡,1982年被正式命名為艾滋病.艾滋病毒有3種傳播方式:血液傳播、母嬰傳播和性接觸傳播.目前,許多具有特異性免疫反應(yīng)的病毒動(dòng)力學(xué)研究日益受到重視,這些研究對(duì)預(yù)防和控制艾滋病有一定的指導(dǎo)作用[1-6].
在治療HIV感染患者方面,有4類抗逆轉(zhuǎn)錄病毒(ARV)藥物:
1)核苷/核苷酸逆轉(zhuǎn)錄酶抑制劑(NRTIs),
2)非核苷逆轉(zhuǎn)錄酶抑制劑(NNRTIs),
3)蛋白酶抑制劑(PIs),
4)進(jìn)入/融合抑制劑(EIs)[7].每種藥物針對(duì)病毒生命周期的不同階段.HIV耐藥的出現(xiàn)最早是由一些核苷抑制劑記錄的,這些抑制劑通過其作為DNA鏈終止劑的活性抑制病毒復(fù)制[8].在臨床中,艾滋病毒耐藥性可由耐藥突變體向易感個(gè)體傳播或通過在治療期間產(chǎn)生的突變而產(chǎn)生.Rong等[9]提供了一個(gè)理論框架,用于探索在使用其他HCV蛋白酶抑制劑或聚合酶抑制劑治療期間,預(yù)先存在的突變變體的患病率和耐藥性的演變.
為了研究HIV感染患者在治療過程中產(chǎn)生耐藥性的機(jī)制,Rong等[7]提出了如下包括敏感型和耐藥型毒株的動(dòng)力學(xué)模型
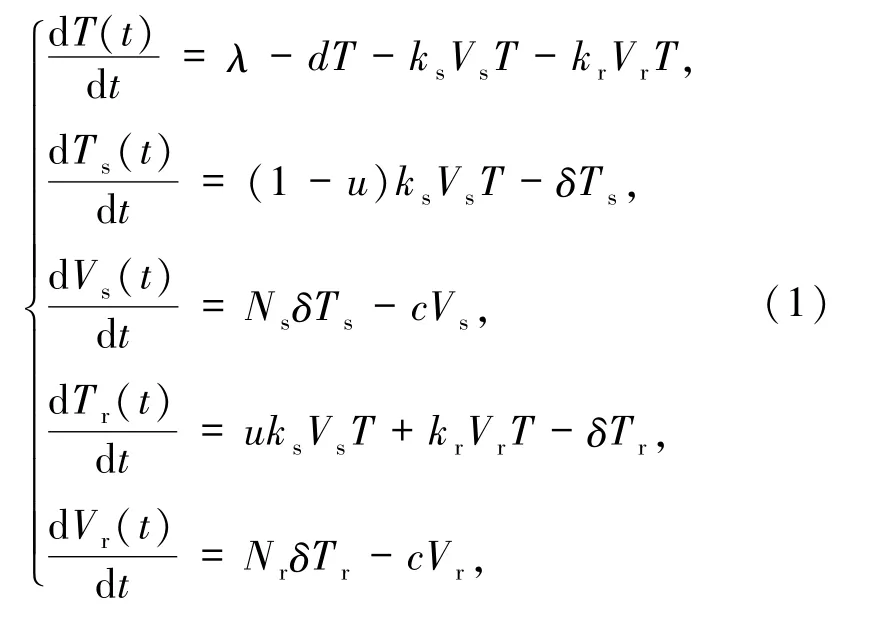
其中,T(t)為未感染細(xì)胞在時(shí)間t的濃度,Ts(t)為敏感毒株感染細(xì)胞在時(shí)間t的濃度,Tr(t)為耐藥毒株感染細(xì)胞在時(shí)間t的濃度,Vs(t)為敏感毒株在時(shí)間t的濃度,Vr(t)為耐藥毒株在時(shí)間t的濃度,λ為未感染細(xì)胞的補(bǔ)充率,d為未感染細(xì)胞的病死率,ks為敏感毒株感染細(xì)胞的感染率,kr為耐藥毒株感染細(xì)胞的感染率,δ為感染細(xì)胞的病死率,Ns為敏感毒株的釋放率,Nr為耐藥毒株的釋放率,c為游離病毒的清除率.
在模型(1)中,參數(shù)u(0<u<1)是病毒RNA逆轉(zhuǎn)錄成原病毒DNA(簡(jiǎn)稱SR轉(zhuǎn)化)過程中,敏感型毒株發(fā)生突變并且產(chǎn)生耐藥性的轉(zhuǎn)化率.需要注意的是,在開始治療之前,宿主體內(nèi)就存在大量的敏感毒株,因此從耐藥型向敏感型毒株的后向突變就可被忽略[10-11].為了準(zhǔn)確地計(jì)算出模型的平衡點(diǎn),以及確定模型平衡點(diǎn)的穩(wěn)定性,生物動(dòng)力學(xué)的入侵閾值,即傳染病中基本再生數(shù)的概念起著決定性的作用[12].
文獻(xiàn)[7]僅研究了模型(1)中無病平衡點(diǎn),具有耐藥型毒株感染的邊界平衡點(diǎn)以及共存平衡點(diǎn)的存在性和局部穩(wěn)定性[7](命題1和命題2),全局動(dòng)力學(xué)包括平衡點(diǎn)的全局漸近穩(wěn)定性和HIV感染模型的一致持續(xù)性都沒有考慮.因此,在本文中,將對(duì)這些問題進(jìn)行深入研究.對(duì)無病平衡點(diǎn)、具有耐藥型毒株平衡點(diǎn)、共存平衡點(diǎn)的局部及全局穩(wěn)定性和HIV感染模型的一致持續(xù)性建立了一系列的閾值準(zhǔn)則.
1 基本性質(zhì)
對(duì)任意整數(shù)n>0,定義
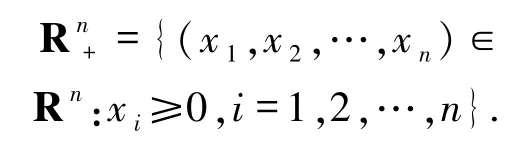
模型(1)的初始條件可表示為
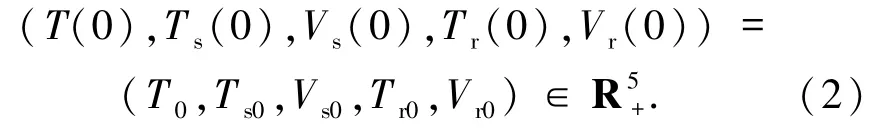
關(guān)于模型(1)解的正性和有界性,有定理1.
定理1對(duì)任何初始值
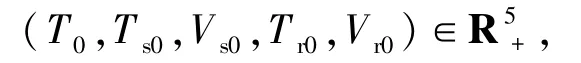
模型(1)具有初始條件(2)的解
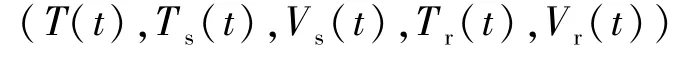
在t∈[0,∞)上有定義,非負(fù)且最終有界.
證明首先,證明解的非負(fù)性.顯然,只需要證明對(duì)滿足任意正初始值(T0,Ts0,Vs0,Tr0,Vr0)的解(T(t),Ts(t),Vs(t),Tr(t),Vr(t))也是正的.定義m(t)=min{T(t),Ts(t),Vs(t),Tr(t),Vr(t)},則有m(0)>0.用反證法.假設(shè)存在一個(gè)t*>0使得m(t*)=0,并且對(duì)所有的t∈[0,t*)有m(t)>0,則有以下5種情形:
1)m(t*)=T(t*),2)m(t*)=Ts(t*),
3)m(t*)=Vs(t*),
4)m(t*)=Tr(t*),
5)m(t*)=Vr(t*).
對(duì)于情形1),由于當(dāng)t∈[0,t*)時(shí)有m(t)>0,則從模型(1)的第一個(gè)方程有

因此,可以得到0=Ts(t*)≥Ts(0)e-δt*>0,矛盾.與上述論證類似,可以得到情形3)、4)和5)都是矛盾的,這表明m(t)總是正的.因此,解(T(t),Ts(t),Vs(t),Tr(t),Vr(t))是非負(fù)的.
下面證明解的最終有界性.定義Lyapunov函數(shù)

這說明解(T(t),Ts(t),Vs(t),Tr(t),Vr(t))也是最終有界的.定理得證.
根據(jù)在文獻(xiàn)[12]中提出的傳染病基本再生數(shù)的概念,定義如下敏感型毒株感染基本再生數(shù)Rs和耐藥型毒株感染基本再生數(shù)Rr為:
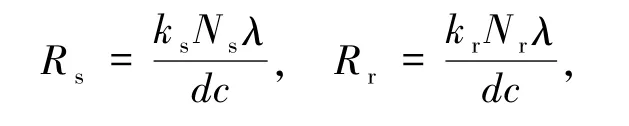
其中,1/c表示毒株i(i=r,s)的平均生命周期;λ/d表示毒株i在感染過程開始時(shí),少量病毒感染易感細(xì)胞的感染細(xì)胞數(shù)量(接近于無感染穩(wěn)態(tài));ki Ni代表一個(gè)i型毒株在其平均存活時(shí)間內(nèi)產(chǎn)生的病毒顆粒數(shù)量.將這些量全部乘積起來就可以得到一個(gè)i型毒株感染細(xì)胞產(chǎn)生新的感染細(xì)胞的數(shù)量,也就是Ri.
下面討論模型(1)平衡點(diǎn)的存在性.平衡點(diǎn)可以由下列方程組得到
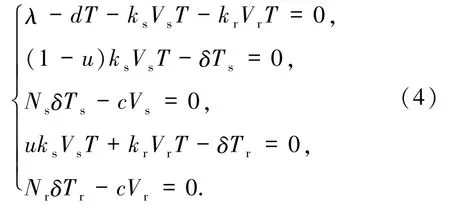
通過求解方程組(4),很容易得到定理2.

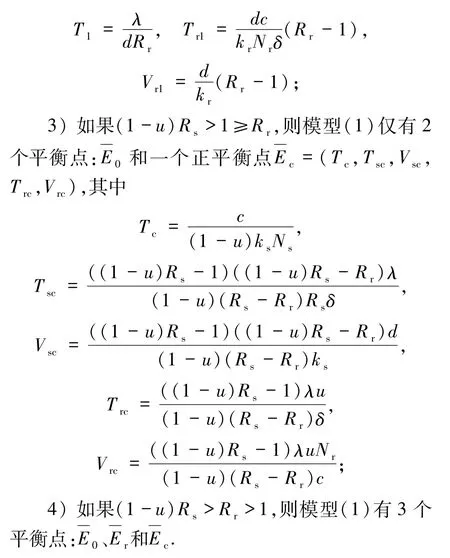
2 全局穩(wěn)定性
令E=(T,Ts,Vs,Tr,Vr)是模型(1)的任意平衡點(diǎn),則在平衡點(diǎn)E處的Jacobi矩陣如下
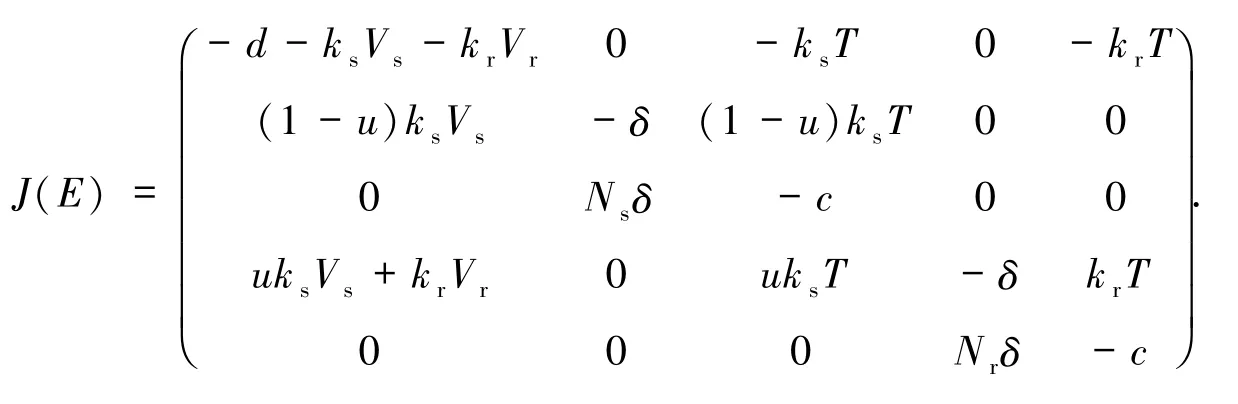

(5)式的一個(gè)根為X1=-d.當(dāng)(1-u)Rs<1且Rr<1時(shí),方程




3 一致持續(xù)性
定理6 如果(1-u)Rs>max{1,Rr},則模型(1)是一致持續(xù)的.也就是說,存在一個(gè)正常數(shù)δ,使得對(duì)模型(1)的任意正解(T(t),Ts(t),Vs(t),Tr(t),Vr(t)),都有
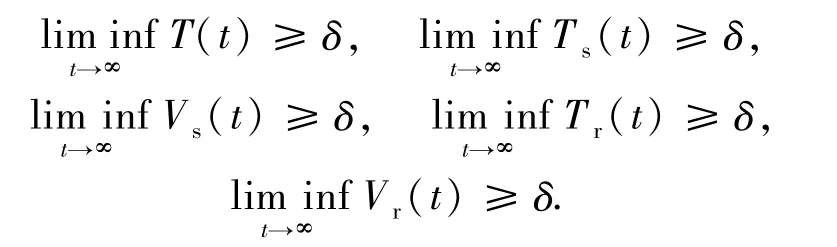
證明對(duì)任意
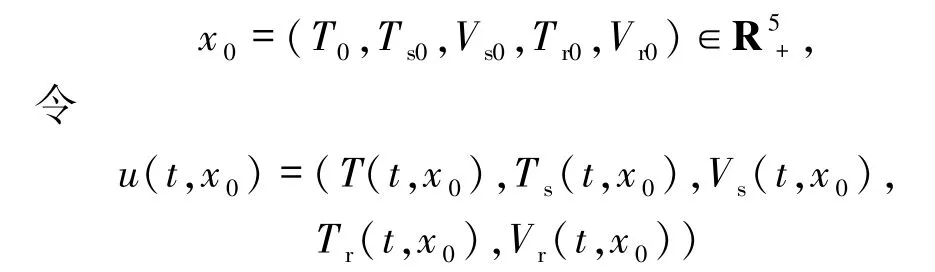
是模型(1)具有初始條件u(0,x0)=x0的解.從定理1的證明可以得到





4 數(shù)值模擬

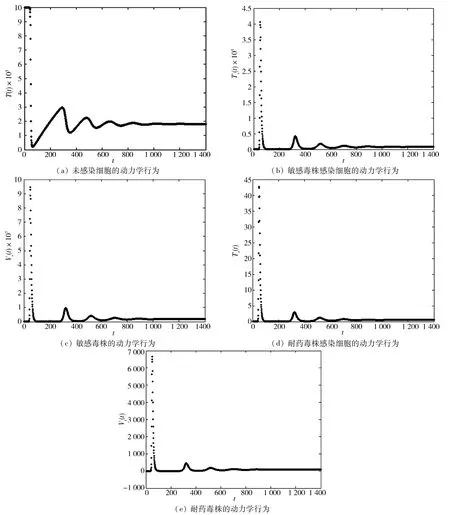
圖1 HIV感染模型的動(dòng)力學(xué)行為Fig.1 Dynamical behaviors of HIV infection model



表1 模型(1)的初始值Tab.1 The initial values of model(1)
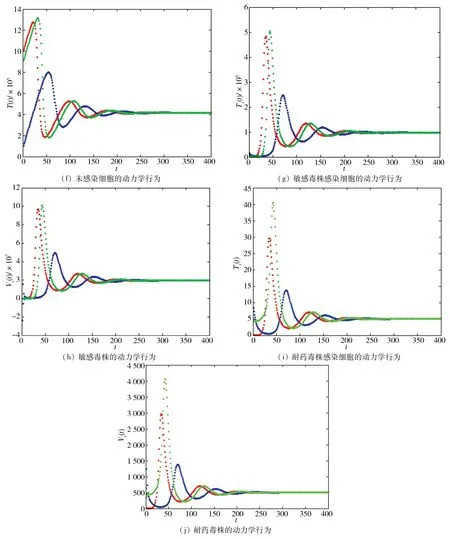
圖2 HIV感染模型的動(dòng)力學(xué)行為Fig.2 Dynamical behaviors of HIV infection model
5 結(jié)論
本文主要研究了敏感型和耐藥型2種毒株的HIV感染模型的全局動(dòng)力學(xué).敏感毒株在逆轉(zhuǎn)錄過程中會(huì)發(fā)生突變并產(chǎn)生耐藥性.主要結(jié)果在定理1~定理6中給出.具體地說,在定理1中得到解的正性和有界性,在定理2中計(jì)算出耐藥型毒株感染平衡點(diǎn)和共存平衡點(diǎn),定理3~定理5中分別得到每個(gè)平衡點(diǎn)的局部和全局漸近穩(wěn)定的充要閾值條件,在定理6中建立HIV感染模型的一致持續(xù)性.
同樣也有很多問題值得更深地去研究.首先,注1~注3考慮了一個(gè)有趣的開問題就是能否在對(duì)應(yīng)的條件下建立平衡點(diǎn)的全局漸近穩(wěn)定性.其次,研究更加復(fù)雜的模型,比如具有飽和發(fā)生率的2種毒株感染模型[15-16].此外,具有空間擴(kuò)散的多毒株感染模型平衡點(diǎn)全局穩(wěn)定問題和具有年齡結(jié)構(gòu)感染模型的動(dòng)力學(xué)行為[17-18]等問題仍然值得去研究.


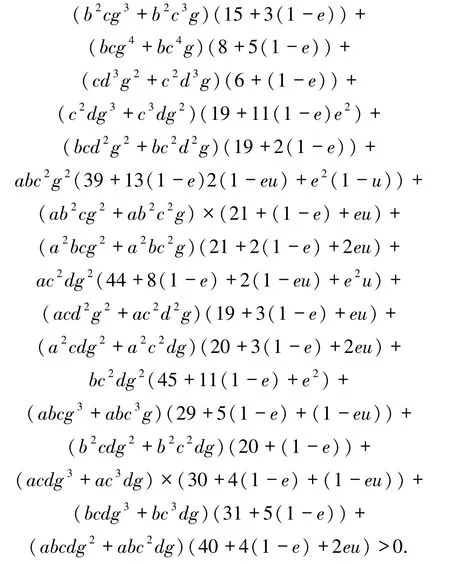
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:40
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
核科學(xué)與工程(2015年4期)2015-09-26 11:59:03
中國當(dāng)代醫(yī)藥(2015年17期)2015-03-01 02:03:58
