棕櫚酸對綿羊單核巨噬細胞炎性因子和氧化應激水平的影響
2021-09-09 08:10:48姚玉昌趙多維肖十雨盧暢亓美玉
東北農業大學學報 2021年7期
姚玉昌,趙多維,肖十雨,盧暢,亓美玉
(1.黑龍江普通高等學校動物遺傳育種與繁殖重點實驗室,哈爾濱 150030;2.東北農業大學動物科學技術學院,哈爾濱 150030;3.黑龍江省農業科學院畜牧研究所,哈爾濱 150086)
脂肪酸(Fatty acid,FA)是由偶數個碳原子組成的直鏈脂肪族羧酸,可調節細胞內信使傳遞[1]、影響部分關鍵基因轉錄[2],對細胞代謝存在直接調控作用。脂肪酸按其碳鏈長度可分為短鏈(2-4 C)、中鏈(6-10 C)及長鏈(12-26 C)脂肪酸,按其飽和度又可分為飽和脂肪酸和不飽和脂肪酸。研究表明,高濃度長鏈飽和脂肪酸具有一定細胞毒性,可誘導炎性因子過量表達[3]、ROS產生、細胞自噬和凋亡通路的激活[4];而長鏈n-3多不飽和脂肪酸則具有一定程度緩解作用[5]。棕櫚酸(Palmitic acid,PA)作為一種典型長鏈飽和脂肪酸,具有較強的脂毒性作用,可引起細胞內過度炎癥反應[6]和氧化應激[7]。亞麻酸(Linoleic acid,LNA)作為一種n-3類多不飽和脂肪酸,可緩解過度炎癥反應[8],改善細胞氧化應激狀態[9]。
Toll樣受體4(Toll-like receptor 4,TLR4)作為一種模式識別受體,可特異性識別革蘭氏陰性菌細胞壁的脂多糖(LPS)成分,激活MyD88依賴和非依賴信號通路,在多種細胞中誘發炎癥、氧化應激、自噬和凋亡等生物學反應[10-12]。TLR4基因除可被LPS激活外,其還可被長鏈飽和脂肪酸激活,進一步活化其下游信號通路[13],誘發細胞中過度炎癥和氧化應激產生[14]。前期研究結果表明,在活體水平上高濃度PA可誘發綿羊單核巨噬細胞和脂肪組織中炎性因子表達上調,激活TLR4基因下游MyD88依賴信號通路,但TLR4基因確切介導作用仍不明確[15]。
因此,本研究分離培養綿羊單核巨噬細胞,揭示PA處理后細胞中炎性因子表達和氧化應激水平變化趨勢,探討TLR4基因在此過程中介導作用,分析LNA可能存在的緩解效應,為闡明長鏈飽和脂肪酸調控免疫細胞代謝相關機制提供幫助。
1 材料與方法
1.1 試驗動物及樣品收集
選擇健康、體況良好18~21月齡德國肉用美利奴公羊6只,飼養于東北農業大學綿羊新品種培育基地,每日提供全價精飼料0.5 kg,自由采食羊草,自由飲水。頸靜脈采集全血50 mL,置于含有肝素的抗凝管中,用于單核巨噬細胞分離。
1.2 脂肪酸:BSA絡合物制備
將0.2 g PA加入至1 mL無水乙醇中溶解,作為儲存液;利用預熱槍頭取10μL儲存液加入至1 mL 10%無脂肪酸的BSA(A6003,Sigma)中,震蕩30 min,42℃加熱30 min,重復此過程至完全溶解。37℃水浴過夜絡合后,0.22μm濾器過濾,-20℃保存待用。
將0.1 g LNA加入至2 mL無水乙醇中溶解,環境中充入氮氣排氧,避免LNA被氧化,作為儲存液;取40μL儲存液加入至1 mL 10%無脂肪酸的BSA(A6003,Sigma)中,震蕩30 min,42℃加熱30 min,重復此過程至完全溶解。37℃水浴過夜絡合后,0.22μm濾器過濾,-20℃保存待用。
1.3 單核巨噬細胞分離及培養
采集的抗凝血用HBSS按1∶1稀釋,小心加至等體積淋巴細胞分離液(購自天津灝洋生物制品科技有限公司)液面上,2 500 r·min-1(×500 g)離心20 min,此時離心管中由上至下細胞分4層,收集第2層絮狀乳白色淋巴細胞至離心管中,2 300 r·min-1(×500 g)離心7 min,棄上清。將所得沉淀細胞利用PBS吹打重懸2次,再用RPMI-1640培養基重懸后,以每個孔1×105個細胞密度接種于12孔板中,待單核巨噬細胞貼壁生長后,再用含0.2% FBS的低血清RPMI-1640培養基在37℃、5%CO2恒溫培養箱培養。
根據文獻報道[16-17]和課題組前期研究,本試驗中給予1 mmol·L-1的PA和LNA處理細胞。設置對照組(含有等濃度BSA,相同體積的培養液)、LPS(100 ng·mL-1)處理組(L4391,Sigma)、PA(1 mmol·L-1)處 理 組(P5585,Sigma)、LNA(1 mmol·L-1)處理組(L1012,Sigma)、PA(1 mmol·L-1)+TAK242(3μmol·L-1)共處理組和PA(1 mmol·L-1)+LNA(1 mmol·L-1)共處理組。其中TAK-242提前預處理6 h,LNA預處理1 h。每組設置3個重復孔,于處理后4 h收集樣品用于后續分析。
1.4 實時熒光定量PCR
收取單核巨噬細胞,按照RNA提取試劑盒(RE-03113,FORE GENE)說明書提取總RNA。取1μL RNA樣品在1.5%瓊脂糖凝膠上120 V電壓電泳10 min,檢測RNA完整度。利用超微量分光光度計(61010-1,DeNovix)檢測RNA樣品純度和濃度,調整RNA濃度至同一水平。按照反轉錄試劑盒(RR037Q,TaKaRa)說明書,反轉錄后得到cDNA。利用ABI7300實時熒光定量PCR檢測系統和FastStart Universal SYBR Green Master(S2024,Roche)試劑盒,檢測TLR4、IFN-γ、TNF-α、IL-1β、IL-6、SOD、iNOS、CAT等基因表達水平,以β-actin為內參基因,采用相對定量法測定基因表達水平。所用的引物信息見表1,由南京金唯智生物技術公司合成。擴增體系為10μL反應混合體系,含有5μL SYBR Green Master,上、下游引物各0.4μL,3.2μL ddH2O和1μL cDNA模板;反應條件為:50℃2 min,95℃10 min;95℃15 s,退火1 min,40個循環;每孔設置3個重復。

表1 PCR引物序列Table 1 PCR primer sequence
1.5 Western blot
收集PA和LNA處理4 h后單核巨噬細胞,利用含有蛋白酶抑制劑(SCP0110,Roche)和PMSF(329-98-6,Roche)的RIPA緩沖液(P0013B,Bey?otime)裂解。隨后,使用BCA蛋白質檢測試劑盒(P0012S,Beyotime)對蛋白質進行定量。將等量蛋白質在12%SDS-PAGE上分離,并轉移到PVDF膜(DVPP00010,Millipore)上,與抗TLR4(1∶1 000;AF7017,Affinity)的一抗孵育后,和辣根過氧化物酶偶聯的二抗(1∶1 000;A0208,Beyotime)孵育,通過化學發光使膜可視化(Thermo Fisher Scientific),利用ImageJ軟件分析蛋白質條帶。
1.6 ROS染色
在各組處理結束前40 min時,于37℃條件下將10μmol·L-1DCFH-DA(E004,南京建成)加入至細胞培養液中,利用熒光顯微鏡(Olympus,X71)觀察細胞內氧化DCF引起的熒光值,綠色熒光信號代表ROS水平。
1.7 氧化和抗氧化指標檢測
于PA和LNA處理4 h后,刮取培養皿上單核巨噬細胞并離心收集沉淀,超聲波破碎。利用總抗氧化能力(T-AOC)測定試劑盒(A015-1,南京建成)、超氧化物歧化酶(SOD)測定試劑盒(A001-3,南京建成)、過氧化氫酶(CAT)測定試劑盒(A007,南京建成)、一氧化氮合酶(NOS)測定試劑盒(A014-1,南京建成)檢測細胞中氧化和抗氧化指標水平,具體操作步驟和判定標準參照說明書執行。
1.8 數據分析
利用SPSS 18.0軟件對數據作單因素方差分析(ANOVA)分析和Duncan多重比較。“*”代表P<0.05,表示差異顯著;“**”代表P<0.01,表示差異極顯著。所有數據均為平均值±標準誤(Mean±SEM)。
2 結果與分析
2.1 單核巨噬細胞中TLR4基因表達變化
利用實時熒光定量PCR和Western blot檢測各處理組中TLR4基因表達水平,結果見圖1。在1 mmol·L-1PA處理4 h后,TLR4基因表達水平上調,與LPS刺激后趨勢相似,均顯著高于對照組(P<0.05);利用TAK242抑制TLR4基因后,其表達水平顯著下調(P<0.05),與對照組間差異不顯著(P>0.05);PA與LNA共處理后,TLR4基因的表達水平也顯著下調(P<0.05),與對照組間差異不顯著(P>0.05);而LNA單獨處理對TLR4基因表達水平無顯著影響(P>0.05)。
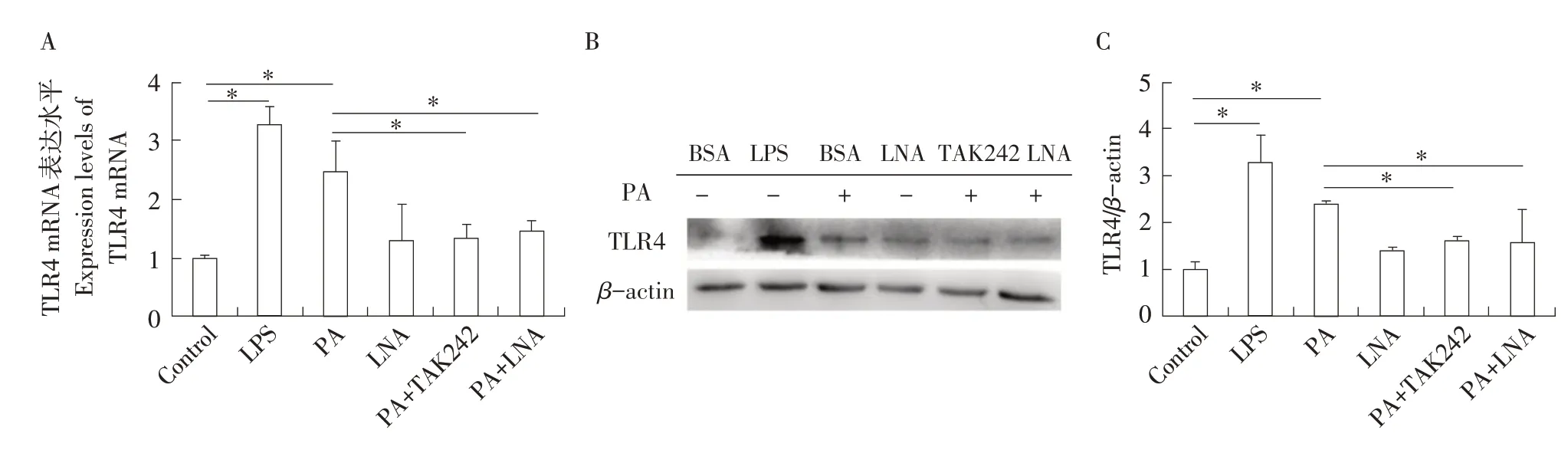
圖1 不同處理條件下綿羊單核巨噬細胞中TLR4基因表達水平Fig.1 Expression of TLR4 gene in sheep mononuclear macrophages under different treatment
2.2 單核巨噬細胞中炎性因子表達變化
利用實時熒光定量PCR檢測各處理組中IFN-γ、TNF-α、IL-1β、IL-6等炎性因子表達水平,結果見圖2。在1 mmol·L-1PA處理4 h后,IFN-γ、TNF-α、IL-1β和IL-6基因表達上調,其中IFN-γ和IL-1β基因表達水平顯著高于對照組(P<0.05),TNF-α和IL-6基因表達水平極顯著高于對照組(P<0.01),整體上各炎性因子表達變化與LPS刺激后趨勢相似;利用TAK242抑制TLR4基因后,TNF-α和IL-6基因表達水平極顯著下調(P<0.01),4種炎性因子表達水平與對照組間均差異不顯著(P>0.05);PA與LNA共處理后,TNF-α和IL-6基因表達水平也極顯著下調(P<0.01),4種炎性因子表達水平與對照組間均差異不顯著(P>0.05);而LNA單獨處理對以上4種炎性因子表達無顯著影響(P>0.05)。
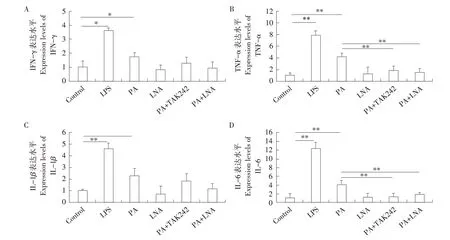
圖2 不同處理條件下綿羊單核巨噬細胞中各炎性因子表達水平Fig.2 Expression levels of inflammatory factors in sheep mononuclear macrophages under different treatment
2.3 單核巨噬細胞中氧化應激相關指標的變化
利用實時熒光定量PCR檢測各處理組中iNOS、SOD、CAT等氧化應激相關基因表達水平,結果見圖3。在1 mmol·L-1PA處理4 h后,iNOS基因表達水平顯著高于對照組(P<0.05),SOD和CAT基因表達水平顯著低于對照組(P<0.05),整體上與LPS刺激后趨勢相似;利用TAK242抑制TLR4基因后,iNOS基因表達水平顯著下調(P<0.05),3個氧化應激相關基因表達水平與對照組間均差異不顯著(P>0.05);PA與LNA共處理后,對3個氧化應激相關基因表達無顯著影響(P>0.05);而LNA單獨處理對以上3種氧化應激相關基因表達也無顯著影響(P>0.05)。
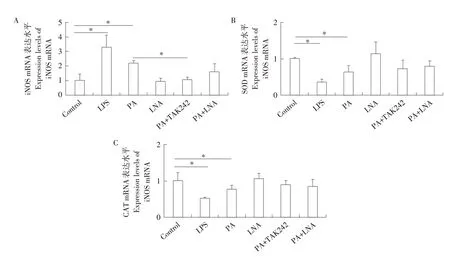
圖3 不同處理條件下綿羊單核巨噬細胞中氧化應激相關基因表達水平Fig.3 Expression levels of oxidative stress related genes in sheep mononuclear macrophages under different treatment
利用氧化應激試劑盒檢測各處理組中iNOS、CAT、SOD、T-AOC等氧化和抗氧化指標變化結果見圖4。在1 mmol·L-1PA處理4 h后,iNOS水平顯著升高(P<0.05),SOD、CAT、T-AOC水平顯著降低(P<0.05),整體上與LPS刺激后趨勢相似;利用TAK242抑制TLR4基因后,iNOS水平顯著下降(P<0.05),T-AOC水平顯著升高(P<0.05),4個氧化應激相關指標水平與對照組間均差異不顯著(P>0.05);PA與LNA共處理后,iNOS水平也顯著下降(P<0.05),T-AOC水平顯著升高(P<0.05),4種氧化和抗氧化指標水平與對照組間均差異不顯著(P>0.05);而LNA單獨處理對以上4種氧化和抗氧化指標水平無顯著影響(P>0.05)。
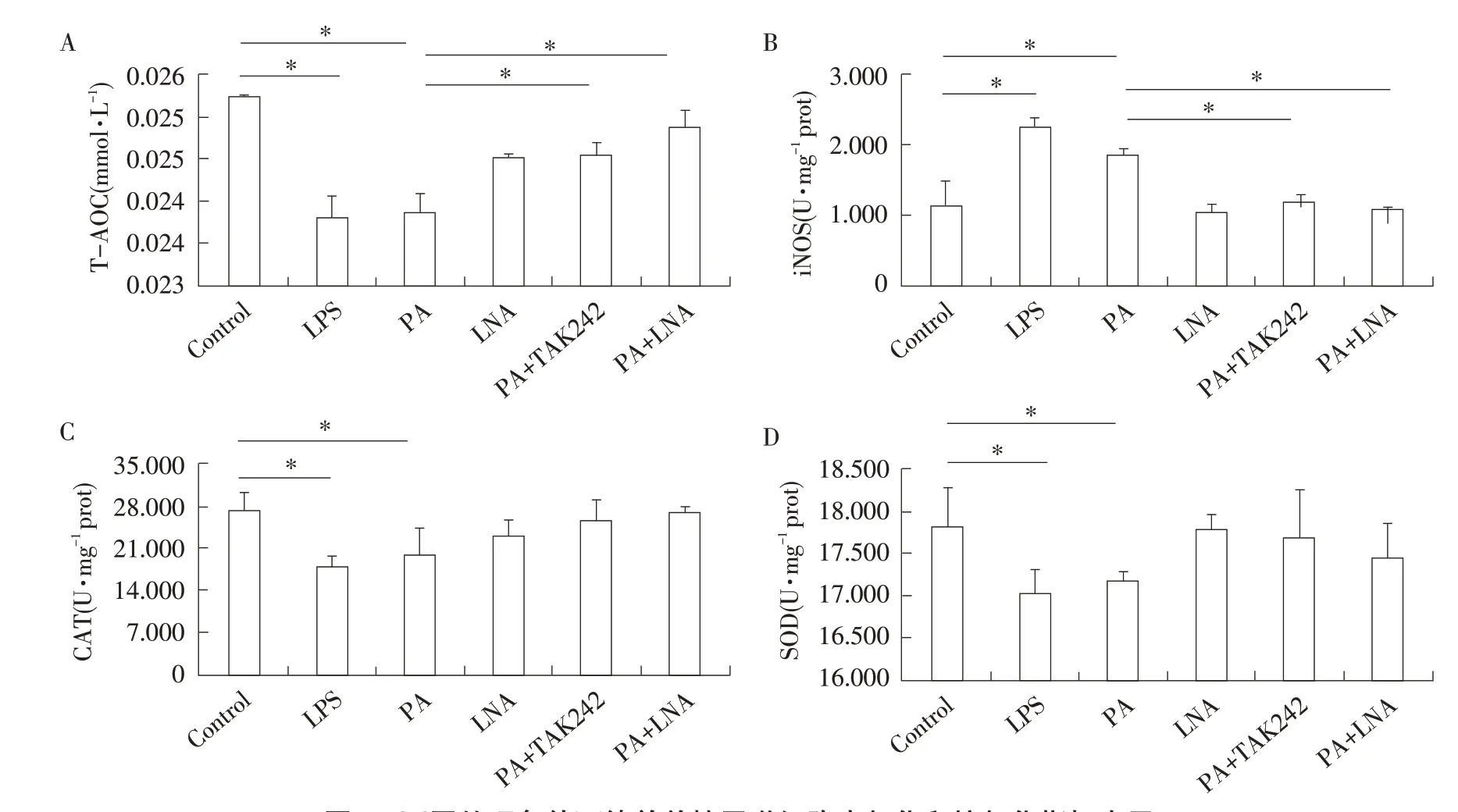
圖4 不同處理條件下綿羊單核巨噬細胞中氧化和抗氧化指標水平Fig.4 Expression levels of oxidation and antioxidant indices in sheep mononuclear macrophages under different treatments
2.4 單核巨噬細胞中ROS水平變化
DCFH-DA染色結果表明見圖5。在1 mmol·L-1PA處理4 h后,ROS的水平顯著升高(P<0.05),與LPS刺激后趨勢相似;利用TAK242抑制TLR4基因后,ROS水平顯著下降(P<0.05),與對照組間差異不顯著(P>0.05);PA與LNA共處理后,ROS水平也顯著下降(P<0.05),與對照組間差異不顯著(P>0.05);而LNA單獨處理對ROS水平無顯著影響(P>0.05)。
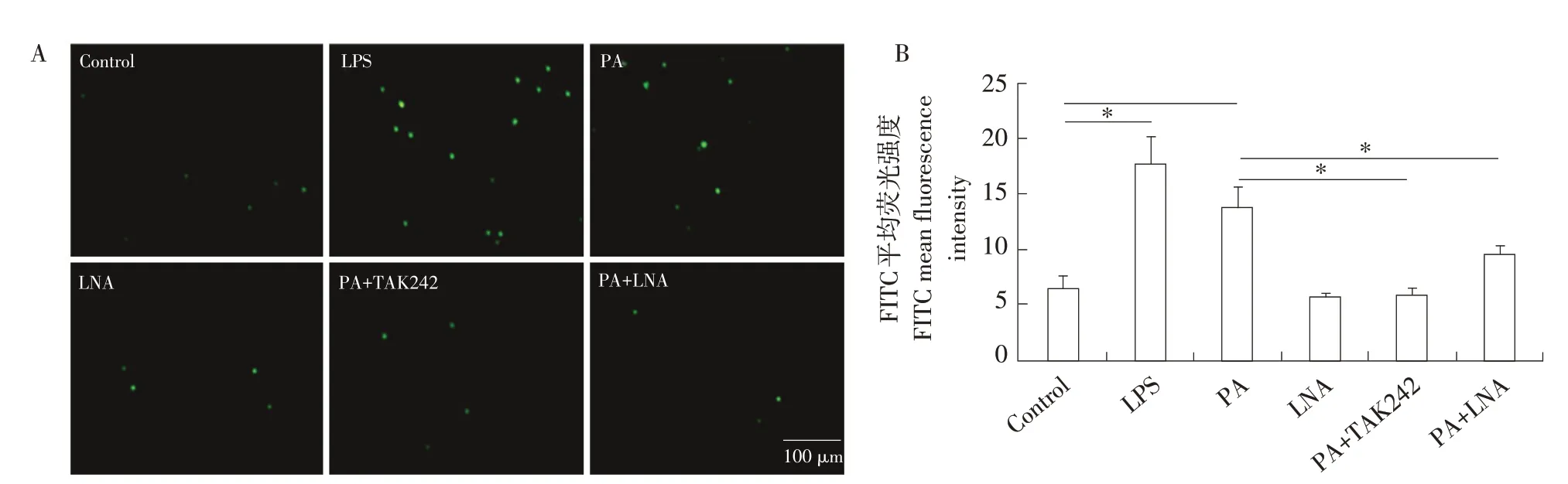
圖5 不同處理條件下綿羊單核巨噬細胞中ROS水平Fig.5 ROS levels in sheep mononuclear macrophages under different treatments
3 討論
脂肪酸對細胞代謝調控作用與其碳鏈長度、雙鍵數量及位置密切相關。目前,研究表明長鏈飽和脂肪酸具有明顯脂毒性,過量累積導致免疫細胞中炎癥激活,促炎因子分泌增加[3]。飽和脂肪酸脂毒性與碳鏈長度呈正比,PA作為含有16個碳的長鏈飽和脂肪酸,可增加巨噬細胞中IL-1β和TNF-α分泌[18]。本研究結果表明,PA處理綿羊單核巨噬細胞后,IFN-γ、TNF-α、IL-1β、IL-6等炎性因子表達水平顯著上調,與陽性對照LPS處理組表現相近趨勢。除LPS以外,脂肪酸也可作為配體激活TLR4基因[13],其中,PA可被TLR4胞膜外區結構識別,并活化其下游信號通路[19]。Suganami等發現飽和脂肪酸處理TLR4基因突變的小鼠巨噬細胞后,TNF-α合成量顯著減少[20]。Sunita等在神經細胞中研究結果也顯示,PA可能通過TLR4引起TNF-α和IL-6釋放,誘發炎癥產生[21]。本研究結果表明,PA處理綿羊單核巨噬細胞后,導致TLR4基因表達上調,炎性因子表達水平升高,而抑制TLR4基因后炎性因子的表達水平顯著下降,進一步證明TLR4基因在介導PA誘發的炎性反應過程中扮演重要角色。部分不飽和脂肪酸可起到一定抗炎作用,亞油酸可以在肝癌細胞系中逆轉PA誘導的炎癥反應,顯著抑制IL-8產生[22];鼠單核細胞研究也表明,n-3類二十碳五烯酸和二十二碳六烯酸可降低IL-1、IL-2、TNF-α等炎性因子表達水平[23]。本研究表明,LNA作為一種n-3類多不飽和脂肪酸,對綿羊單核巨噬細胞中TLR4基因和炎性因子表達無顯著影響,但可在一定程度上緩解PA促炎作用。
一般認為過度氧化應激狀態導致免疫細胞中蛋白質翻譯異常,影響細胞正常代謝過程。PA可通過損傷線粒體氧化呼吸鏈正常功能,誘導細胞發生氧化應激反應[24]。PA可在牛輸卵管上皮細胞中誘導ROS水平升高[18]。Liu等在大鼠心臟細胞研究中發現PA可通過TLR4下游MAPK通路增強ROS產生,進一步說明ROS產生可能與TLR4激活有關[25]。本研究結果表明,PA處理綿羊單核巨噬細胞后,ROS水平顯著升高,而抑制TLR4基因后ROS水平顯著下降。PA刺激炎癥并分泌產生ROS[26],ROS可通過TLR4信號通路激活iNOS,導致NO釋放,NO通過產生過氧化物酶和超氧化物幫助消滅氧化應激源頭[27]。Juntian等發現,PA可以顯著提高iNOS表達,導致細胞氧化應激水平提高[28]。
本研究結果表明,PA處理綿羊單核巨噬細胞后,氧化指標水平提高、抗氧化指標水平降低,導致細胞中整體氧化應激水平提高。而抑制TLR4基因后,iNOS等細胞氧化指標顯著下降,降低氧化應激水平。多不飽和脂肪酸可抑制飽和脂肪酸誘導的TLR4基因表達[29],緩解細胞內氧化應激水平。n-3類多不飽和脂肪酸可以抑制巨噬細胞中iNOS表達[30];攝入n-3類多不飽和脂肪酸后,血液中二十碳五烯酸和二十二碳六烯酸含量增加,誘發抗氧化指標SOD水平升高,緩解氧化應激狀態[31]。本研究表明,LNA對綿羊單核巨噬細胞中TLR4基因表達和抗氧化指標無顯著影響,但可在一定程度上通過降低氧化水平緩解PA誘發的氧化應激反應。
4 結論
PA處理后導致綿羊單核巨噬細胞炎性因子和氧化應激水平升高,TLR4基因在此過程中起重要介導作用,LNA可能在一定程度上緩解PA誘發的細胞炎癥反應和氧化應激。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
西南軍醫(2016年6期)2016-01-23 02:21:19
學苑創造·A版(2015年11期)2016-01-14 09:03:27
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
食品科學(2013年15期)2013-03-11 18:25:48
