茶小綠葉蟬成蟲唾液細菌蛋白的鑒定
2021-09-06 05:39:02鈕羽群夏露霞伍春芳景凱婷程墻義王夢馨韓寶瑜
昆蟲學報 2021年8期
潘 鋮, 鈕羽群, 夏露霞, 伍春芳, 景凱婷,程墻義, 王夢馨, 韓寶瑜,*
(1. 中國計量大學生命科學學院, 浙江省生物計量及檢驗檢疫技術重點實驗室, 杭州 310018;2. 安徽農業大學植物保護學院, 合肥 230000)
在昆蟲進化的生命歷程中,其體內的共生菌扮演著不可替代的作用,能幫助宿主昆蟲更好地進行營養攝取、生長發育、解毒、天敵防御和免疫應答等生命活動(譚周進等, 2005; Engel and Moran, 2013)。刺吸式口器昆蟲,因僅能取食木質部、韌皮部的汁液,極易造成取食營養結構不平衡,其生命活動部分營養功能需依靠共生菌分泌纖維素等碳水化合物酶來維持(Anandetal., 2010; Xueetal., 2014; Douglas, 2015)。唾液作為昆蟲和植物之間互作的基礎性物質,在互作中起著復雜且重要的作用,有研究指出唾液中含有的共生菌來源于與口針相連的唾液管道(唾液腺)和食物管道(腸道)(殷海娣等, 2006; 王雪麗等, 2016; 尚哲明等, 2019; Xuetal., 2019)。煙粉虱Bemisiatabaci唾液腺中被發現存在共生細菌(立克次氏體Rickettsia)(Bruminetal., 2012);張瑛等(2015)以斑透翅蟬Hyalessamaculaticollis唾液腺為對象,利用16S rRNA序列分離鑒定了變形菌門(Proteobacteria)和厚壁菌門(Firmicutes)的7種細菌。直接以唾液為研究對象,唐明等(2014)分離鑒定發現褐飛虱Nilaparvatalugens唾液分泌物中存在4種γ變形菌綱和2種β變形菌綱細菌;苗雨桐等(2017)從褐飛虱成蟲水狀唾液鑒定出屬于變形菌門、放線菌門和厚壁菌門的22種細菌。可見唾液中存在部分昆蟲共生菌,并可能參與宿主昆蟲與植物的互作機制。
茶小綠葉蟬Empoascaflavescens是我國茶區嚴重發生的害蟲,該害蟲種群密度較大,繁殖能力強,極難防治;為害后茶樹嫩梢變紅、卷曲,嚴重影響名優茶的色、香、味和形等品質特征,制約茶葉的產量。幾十年以來依賴化學防治,已致使茶小綠葉蟬對啶蟲脒和聯苯菊酯的抗藥性達高抗水平,對吡蟲啉達中抗水平;同時業內估計每年茶園中40%的施藥量被用于防治茶小綠葉蟬(莊家祥等, 2009)。隨著高通量測序技術、宏基因組學和蛋白質組學等技術的發展,借助新技術分析成為趨勢。劉小改等(2016)和王天召等(2019)分別利用高通量測序分析稻縱卷葉螟Cnaphalocrocismedinalis和褐飛虱腸道細菌,分別得到604和235個分類單元;毛迎新等(2018)利用高通量測序分析了5個地理種群的茶小綠葉蟬成蟲體內共生細菌種類,共鑒定41門116綱105種細菌。目前還沒有葉蟬唾液細菌蛋白的研究報道。本研究以茶小綠葉蟬成蟲唾液的細菌蛋白為切入點,采用液相色譜-串聯質譜(liquid chromatography-tandem mass spectrometry, LC-MS/MS)蛋白分析法鑒定唾液中細菌蛋白,為葉蟬細菌蛋白研究和其在昆蟲取食、致害和解毒等生命過程的作用提供理論基礎。
1 材料與方法
1.1 供試昆蟲
實驗所用的茶小綠葉蟬來自浙江省湖州市安吉縣的露天茶園(安吉白茶品種),飼養于人工氣候室(溫度26±1℃,相對濕度65%±5%,光周期14L∶10D)內,仍用采集的安吉白茶茶枝飼養。
1.2 唾液收集
茶小綠葉蟬水狀唾液的收集方法使用的是Parafilm膜夾營養液法(圖1)。首先使用Parafilm膜在玻璃管(玻璃管兩頭開口,管口內徑5.5 cm,高為10.0 cm,管壁均勻分布小孔用于透氣)的一端制作葉蟬取食裝置,將第1層Parafilm膜均勻拉開到適宜薄度覆蓋于玻璃管一端,使用移液槍吸取3 mL 5%無菌蔗糖溶液添加于膜中間,再將薄度一致的第2層膜覆蓋其上,排出氣泡將液體鎖定于兩膜之間,最后在最外部添加第3層相對較厚的保護膜。從玻璃管另一端將20頭經過1 h饑餓處理的葉蟬成蟲放入,以濕潤的紗布封口。根據上述方法依次制作相同的唾液收集裝置,排列于透氣網盒中,確保取食裝置位于上端,略微傾斜,轉移至人工氣候室(溫度26±0.5℃,相對濕度65%±5%,光周期14L∶10D)內。待葉蟬取食24 h后,使用無菌注射器穿刺收集被取食過的蔗糖溶液,儲存于-80℃的超低溫冰箱。共收集6 000頭葉蟬取食過的蔗糖溶液。
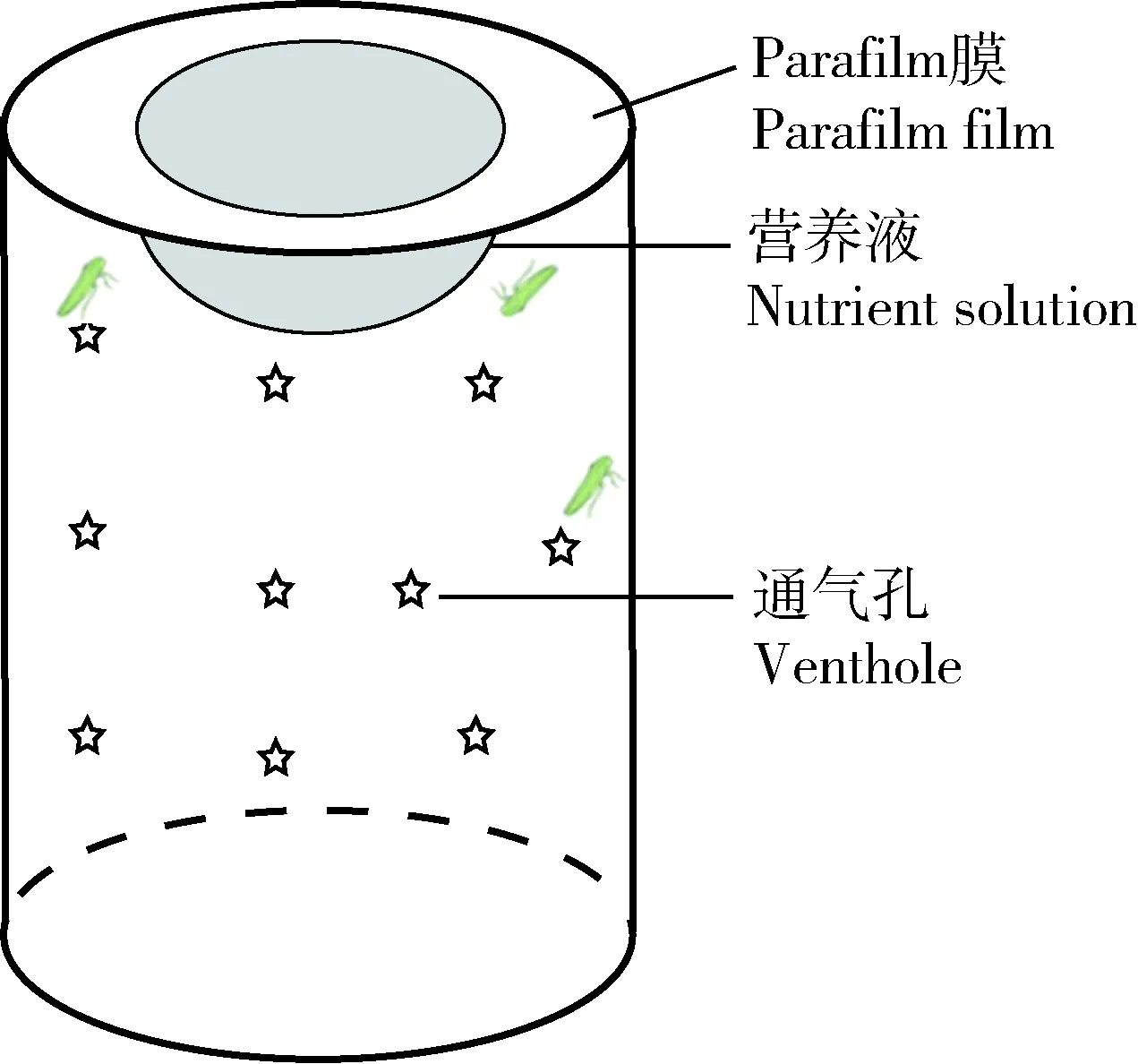
圖1 葉蟬唾液收集裝置示意圖Fig. 1 Schematic diagram of cicada saliva collection device
1.3 唾液蛋白濃縮與酶解
使用超濾方式進行濃縮。首先加入等體積SDS-Tris(2% SDS, 10 mmol/L Tris-HCl, pH 8.0)裂解液,混勻后沸水浴10 min;用3 kD的超濾離心管濃縮收集的樣品,濃縮至終體積200 μL;然后在濃縮樣品中加入適量SDT溶液(4% SDS, 100 nmol/L DTT, 150 mmol/L Tris-HCl, pH 8.0),沸水浴15 min后18 000 g室溫離心15 min,收集上清液;取20 μL樣品上樣,聚丙烯酰胺凝膠電泳(SDS-PAGE),染色;最后采用BCA法進行蛋白質定量,分裝樣品,儲存于-80℃的超低溫冰箱。
使用膜輔助樣品制備技術法(filter-aided sample preparation, FASP)對蛋白進行酶解(Winiewskietal., 2009)。取100 μg蛋白質溶液,加入二硫蘇糖醇(DTT)至終濃度為100 mmol/L。沸水浴5 min后冷卻至室溫。加入200 μL的UA Buffer(0.1 mol/L Tri-HCl含8 mol/L尿素, pH 8.5)混勻,轉移至超濾離心管中,14 000 g 離心15 min,棄廢液,重復一次。加入100 μL的IAA Buffer(100 mmol/L IAA溶解于UA Buffer),600 r/min振蕩1 min,室溫避光反應30 min,14 000 g離心15 min。加入100 μL UA Buffer, 14 000 g離心15 min,重復2次。加入100 μL NH4HCO3(25 mmol/L)溶液,14 000 g離心15 min,重復兩次。加入40 μL Trypsin Buffer(2 μg Trypsin溶于40 μL 100 mmol/L NH4HCO3溶液中),600 r/min振蕩1 min,37℃放置16~18 h。換新收集管,14 000 g離心15 min;再加入40 μL NH4HCO3(25 mmol/L),14 000 g離心15 min,收集濾液。采用C18 Cartridge對肽段進行脫鹽,肽段凍干后加入40 μL濃度為0.1%甲酸溶液復溶,最后對肽段定量(測定OD280值)。
1.4 LC-MS/MS蛋白分析
參照定量結果取適量酶解產物進行LC-MS/MS分析,蛋白檢測分析由上海中科新生命生物科技有限公司進行。
液相部分:采用納升級流速HPLC液相系統Easy nLC進行分離。分別配制0.1%甲酸水溶液和0.1%甲酸乙腈水溶液(乙腈為84%)作為緩沖液A液和B液。色譜柱平衡時使用95%的A液進行。C18上樣色譜柱規格:Thermo Scientific EASY Column(2 cm×100 μm, 5 μm C18)。C18分析柱規格:Thermo Scientific EASY Column(75 μm×100 mm, 3 μm C18)。樣品進上樣色譜柱后,再經過分析柱,流速300 nL/min。洗脫程序見表1。
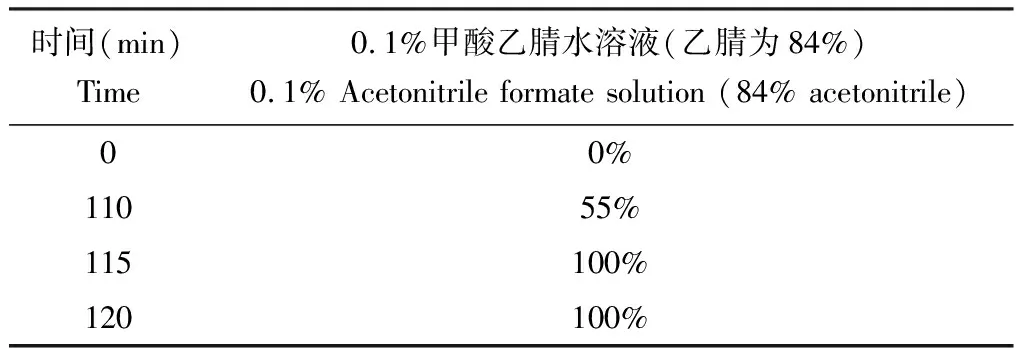
表1 120 min梯度洗脫程序Table 1 120 min gradient elution procedure
1.5 細菌蛋白鑒定
原始文件(raw file) 采用軟件Mascot 2.2(Matrix Science)進行查庫分析。質譜鑒定結果比對UniProt的細菌蛋白數據庫,參數設置參考苗雨桐等(2017),母離子質量差設置為20 mmu,碎片離子誤差為15 μg/g,半胱氨酸上的脲甲基修飾以及蛋氨酸上氧化修飾分別被選定為固定修飾和可變修飾。允許失誤的剪切位點為1。檢索結果以P<0.05為顯著。利用KEGG(Kyoto Encyclopedia of Genes and Genomes)數據庫的KEGG Pathway 數據庫進行信號通路注釋分析。
2 結果
2.1 唾液SDS-PAGE蛋白電泳
本研究對濃縮后的茶小綠葉蟬唾液細菌蛋白進行一維電泳(圖2),蛋白分子量大多數集中分布在35~110 kD之間。

表2 質譜儀參數設置Table 2 Parameter setting of MS
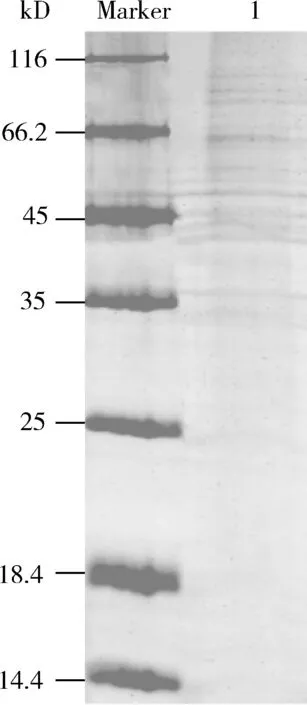
圖2 茶小綠葉蟬成蟲唾液細菌蛋白的分離電泳Fig. 2 Electrophoresis of bacterial proteins in the salivaof Empoasca flavescens adultsMarker: 蛋白質分子量標準Protein molecular weight marker; 1: 目的蛋白Target protein.
2.2 唾液中細菌蛋白的鑒定
本次實驗利用LC-MS/MS蛋白分析法對葉蟬成蟲唾液中的細菌蛋白進行鑒定,利用UniProt公共數據庫進行比對,共鑒定142種細菌蛋白(表3)。分類結果顯示為27個目49種細菌的142種蛋白,隸屬于5個門和11個綱分類階元。以變形菌門(Proteobacteria)細菌蛋白為主,共107個蛋白,占總蛋白數的75%以上,其中又以α-變形菌綱(α-Proteobacteria)51個蛋白和γ-變形菌綱(γ-Proteobacteria)40個蛋白為占比最多的兩類。其次為厚壁菌門(Firmicutes)共28個蛋白,分別屬于桿菌綱(Bacilli)和梭菌綱(Clostrida)。蛋白種類大于10個的目,包括根瘤菌目(Rhizobiales)、鞘脂單包菌目(Sphingomonadales)、腸桿菌目(Enterobacteriales)、假單胞菌目(Pseudomonadale)、芽孢桿菌目(Bacillales)。
在鑒定的142種細菌蛋白中,發現新陳代謝(metabolism)和遺傳信息加工(genetic information processing)是數量最多的兩個類群(圖3)。在新陳代謝類別中,12個蛋白參與氨基酸代謝(amino acid metabolism)通路,涉及天冬氨酸、精氨酸、絲氨酸、賴氨酸、苯丙氨酸、色氨酸、纈氨酸和亮氨酸等生物合成或代謝途徑;11個蛋白參與核苷酸代謝(nucleotide metabolism)通路,包括雙功能嘌呤生物合成蛋白、核苷二磷酸激酶、核苷酸轉移酶、核苷二磷酸激酶等參與嘌呤或嘧啶代謝的蛋白;10個蛋白參與能量代謝(energy metabolism)通路,包括ATP合酶、NADH醌氧化還原酶、甲基四氫甲烷蝶呤環水解酶、絲氨酸羥甲基轉移酶、磷酸烯醇式丙酮酸羧化酶。在遺傳信息加工中, 主要涉及翻譯(translation)通路, 主要以核糖體、氨基酰tRNA生物合成為主。另外,參與膜的形成和運輸、信號傳導(signal transduction)、細胞過程(cellular processes)、蛋白折疊等的蛋白也被鑒定發現。
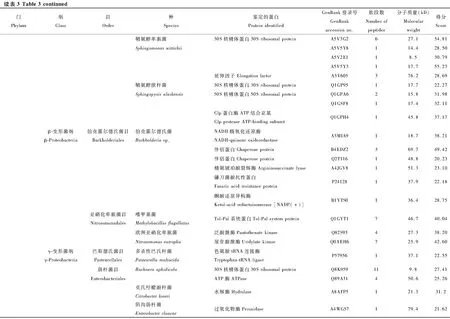

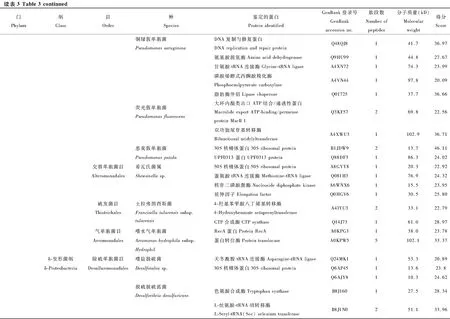
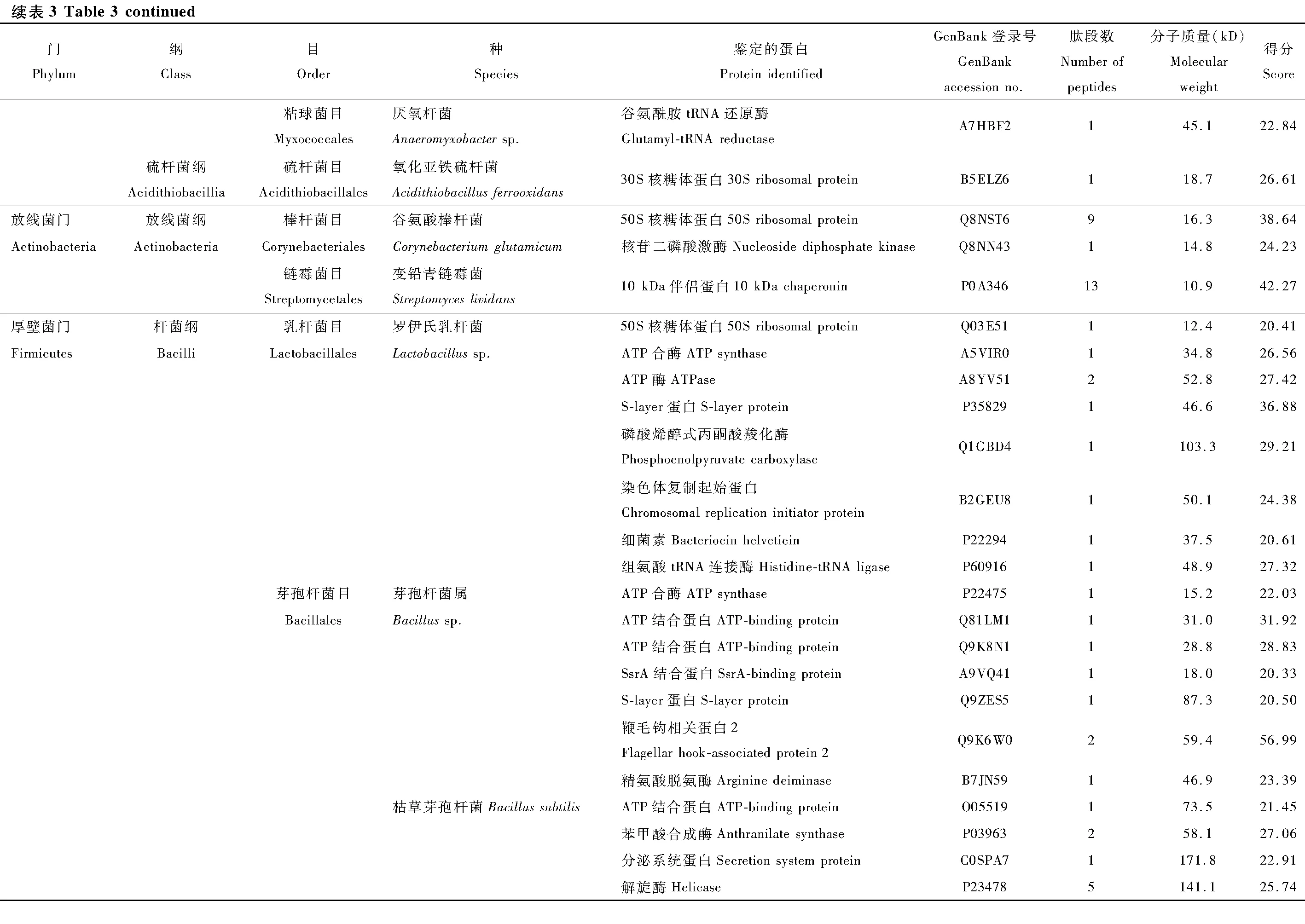
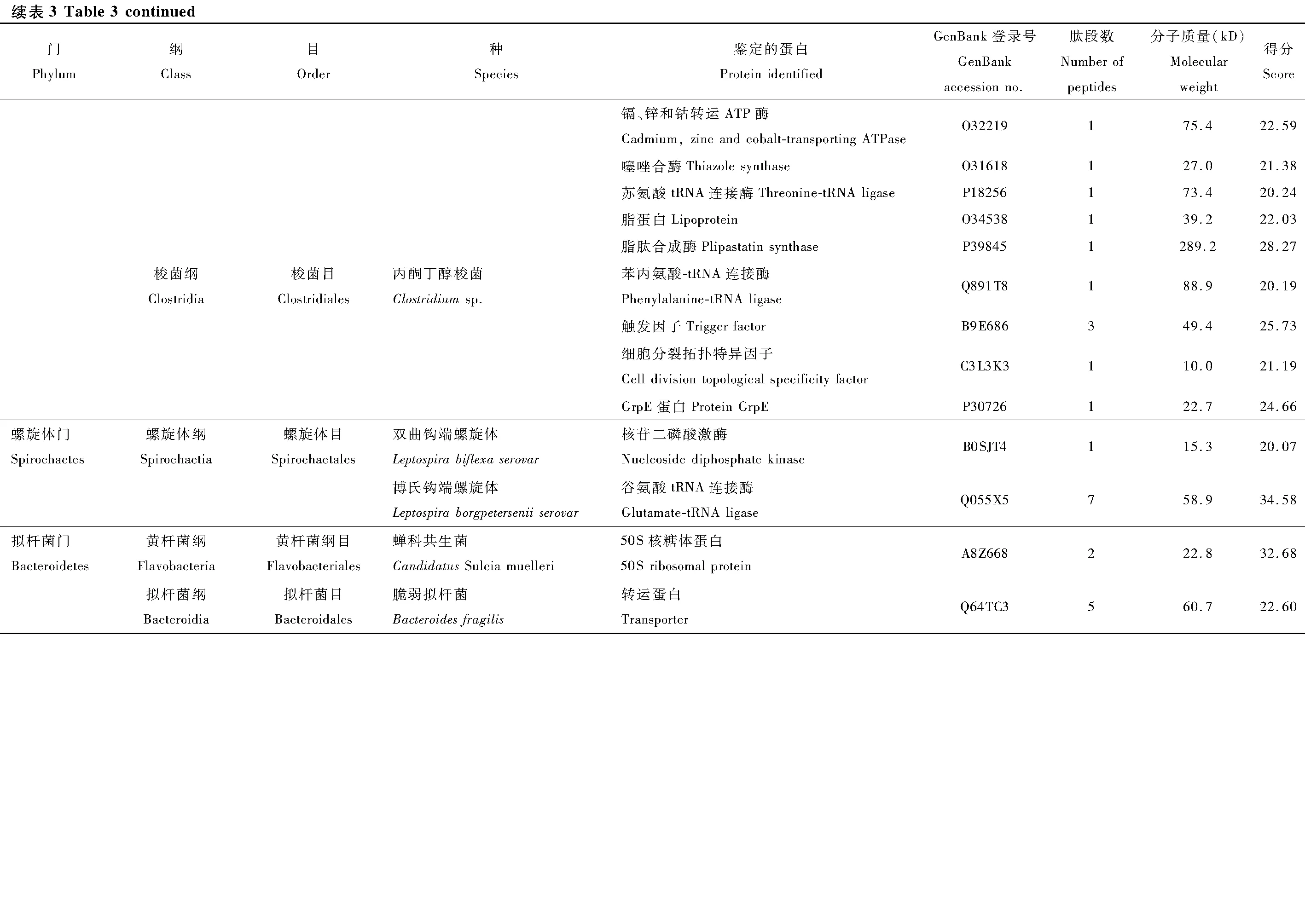
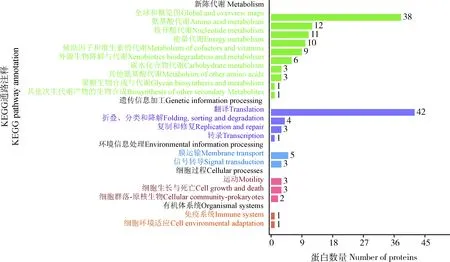
圖3 茶小綠葉蟬成蟲唾液細菌蛋白的KEGG信號途徑分析Fig. 3 KEGG pathway classification of bacterial proteins in the saliva of Empoasca flavescens adults
3 討論
昆蟲共生細菌多樣性分析最常用的鑒定方法有:體外培養鑒定和16S rRNA序列分子標記鑒定。但體外培養需要如特殊營養物質、pH、氧化還原電位等復雜特殊的生存環境,使絕大多數共生菌難以人工培養(Engel and Moran, 2013; 梅承等, 2018),同時利用培養基分離的細菌分類單元相對偏少,效率較低;利用16S rRNA結合限制性片段長度多態性(restriction fragment length polymorphism, RFLP)等技術的分子標記,雖然使鑒定難度降低,鑒定的分類單元增加,但文庫構建和電泳分析均費時費力(Anandetal., 2010; 李香香, 2011; 黃衛強和張和平, 2014)。本研究為解析茶小綠葉蟬在取食過程中唾液內含有的共生菌,首次利用LC-MS/MS法對水狀唾液中的細菌蛋白進行鑒定,并使用細菌UniProt蛋白數據庫進行比對注釋,共發現5門11綱27目49種細菌的142種蛋白。細菌單元分類數量小于利用高通量測序分析結果(毛迎新等, 2018),但結果與利用蛋白質組技術鑒定褐飛虱成蟲唾液發現22種細菌的35種蛋白(苗雨桐等, 2017)類似,唾液細菌數量小于高通量測序分析褐飛虱腸道細菌的結果(王天召等, 2019)。
以韌皮部汁液為食的半翅目昆蟲均有共生微生物,共生菌可為宿主提供氨基酸、維生素和脂肪等必需物質彌補其飲食結構中營養不平衡的缺陷(Feldhaar and Gross, 2009; 李香香等, 2011; Douglas, 2015),同時還對免疫反應、抗病性和抵御天敵等都存在作用(Scarboroughetal., 2005; Vorburger, 2014)。本次共鑒定到精氨琥珀酸裂解酶、精氨酸脫氨酶、天冬氨酸解氨酶、絲氨酸羥甲基轉移酶、色氨酸合成酶、2,3,4,5-四氫吡啶-2,6-二羧酸N-琥珀酰轉移酶、烯醇丙酮酸莽草酸合酶、苯甲酸合成酶、過氧化物酶、吲哚-3-丙酮酸脫羧酶、酮酸還原異構酶涉及8種以上氨基酸的生物合成或代謝途徑,可能為葉蟬的生長發育提供氨基酸(Douglas, 2015)。Wu等(2006)發現Baumanniacicadellinicola被發現可為琉璃葉蟬Homalodiscacoagulata提供維生素,本研究中注釋為參與輔助因子和維生素代謝的9個蛋白涉及根瘤菌目、海洋螺菌目、伯克霍爾德氏菌目、亞硝化單胞菌目、粘球菌目、芽孢桿菌目和硫發菌目等。參與能量代謝的鑒定到甲基四氫甲烷蝶呤環水解酶、磷酸烯醇式丙酮酸羧化酶、ATP合酶和NADH醌氧化還原酶等多種酶類,參與糖酵解、三羧酸循環和電子傳遞鏈等代謝過程可能為宿主提供合成原料和能量。
本次研究所鑒定蛋白均來源于49種細菌,其中假單胞菌目、海洋螺菌目、梭菌目、立克次氏體目、根瘤菌目、伯克霍爾德氏菌和腸桿菌目等均被多次報道在茶小綠葉蟬或頭喙亞目昆蟲體內存在(苗雨桐等, 2017; 毛迎新等, 2018; 王天召等, 2019)。褐飛虱唾液分泌物的16S rDNA檢測發現存在不動桿菌Acinetobactersp.(唐明等, 2014);而唾液細菌蛋白的鑒定發現的22種細菌(苗雨桐等, 2017),其中有17種細菌均在本次的研究中被檢測到。本研究還檢測到頭喙亞目昆蟲的多種常見共生菌,例如:蚜蟲中報道較多的初級共生菌Buchneraaphidicola和次級共生菌Hamiltonelladefensasubsp.Acyrthosiphonpisum(Charles and Ishikawa, 1999; Moran, 2005);在采采蠅中報道的共生菌Sodalisglossinidius和Wigglesworthiaglossinidiabrevipalpis(Chenetal., 1999; Akman, 2001);當然,也包括蟬科昆蟲共生菌CandidatusSulcia muelleri(黃桿菌)、次生共生菌Rickettsia(Nodaetal., 2001; Bruminetal., 2012)。這些結果與毛迎新等(2018)報道的結果一致。有研究指出,H.defensa不但參與2種必需氨基酸和7種非必需氨基酸的合成,還參與大多數必需維生素的合成。也有研究指出,昆蟲體內的初級共生菌會相互配合為寄主提供所需營養元素,葉蟬體內的10種氨基酸,其中Sulcia負責8個氨基酸的合成,另一種共生菌合成其余2種,其中某一氨基酸還有由兩種共生菌共同參與的(McCutcheon and Moran, 2007, 2010; Bennett and Moran, 2013, 2015)。但是本次檢測未發現葉蟬中分布最多的次級共生菌沃爾巴克氏菌Wolbachia(杜慧民等, 2020),與毛迎新等(2018)基于16S rDNA在其所有地理種群均檢測到該菌且為優勢種群的結果存在差異,猜測可能是由于沃爾巴克氏菌與生殖和發育相關且只能進行垂直傳播,在唾液中可能不分布該菌,與苗雨桐等(2017)在褐飛虱唾液中未發現內共生菌Wolbachia和殺雄菌屬Arsenophonus的細菌蛋白的結果一致。本次檢測還發現許多次級共生菌的分布,但是不同種類的葉蟬其體內的次級共生菌需進一步研究。
本研究首次獲得了茶小綠葉蟬唾液共生菌的詳細信息,鑒定多種已報道的葉蟬共生細菌及其他昆蟲的共生菌蛋白,這些蛋白雖然不一定均源于葉蟬唾液細菌群落或腸道細菌代謝,但在葉蟬的生活史中必然發揮著重要的作用,為進一步研究共生菌、茶小綠葉蟬和茶樹之間的關系提供基礎信息。
