西花薊馬GABA受體基因鑒定及FoRDL在多殺霉素抗性中的作用
2021-09-06 05:42:38何秉青華登科袁江江鄭曉斌徐寶云張友軍吳青君
昆蟲學報 2021年8期
王 京, 何秉青, 華登科, 張 坤, 袁江江, 鄭曉斌,徐寶云, 張友軍, 吳青君,*
(1. 中國農業科學院蔬菜花卉研究所, 北京 100081; 2. 北京市昌平區農業技術推廣站, 北京 102200)
γ-氨基丁酸(γ-aminobutyric acid, GABA)是動物神經系統中一種重要的神經遞質,通過與位于突觸后膜的受體結合,使突觸后神經元處于保護性抑制狀態(Bown and Shelp, 1997; Ashbyetal., 2012)。昆蟲的GABA受體(GABA receptors, GABAR)按其結構特征和藥理學性質可分為離子型受體(GABAAR)和代謝型受體(GABABR)兩類。GABAAR是一個由5個亞基組成的寡聚體蛋白(Chebib and Johnston, 2000),昆蟲的GABAAR是環戊二烯類、苯基吡唑類以及大環內酯類殺蟲劑的分子靶標(Buckinghametal., 2005; Rahmanetal., 2012; Nakaoetal., 2015),目前已知有3類亞基:抗狄氏劑亞基(resistance to dieldrin, RDL)(Ffrench-Constantetal., 1991)、配體門控氯離子通道同系物3(ligand-gated chloride channel homologue 3, LCCH3)(Hendersonetal., 1993)和果蠅的GABA和甘氨酸類似受體(GABA and glycine-like receptor ofDrosophila, GRD)(Harveyetal., 1994)。RDL亞基最初得名于黑腹果蠅Drosophilamelanogaster,因其與環戊二烯類殺蟲劑狄氏劑抗性形成相關(Ffrench-Constantetal., 1991),隨后發現該亞基也在昆蟲對阿維菌素和氟蟲腈抗性中起重要作用(Guillemaudetal., 2003; Weietal., 2015; Mengetal., 2019)。GABABR屬于G蛋白偶聯受體(G protein-coupled receptor)家族,該受體通過G蛋白介導連接第二信使,調節控制神經細胞內環腺苷酸(cyclic adenosine monophosphate, cAMP)含量,通過cAMP與細胞內的K+和Ca2+通道相偶聯,進行信號轉導(Kuriyamaetal., 2000; Labouèbeetal., 2007)。
西花薊馬Frankliniellaoccidentalis是一種重要的世界性農業害蟲(Kirk and Terry, 2003),目前,使用殺蟲劑是防治西花薊馬的主要手段,但其不合理使用導致西花薊馬的抗藥性問題日趨嚴重(Demirozeretal., 2012; Gaoetal., 2012; 萬巖然等, 2016)。 據Arthropod Pesticide Resistance Database(Sanches and Wise, 2021)最新統計,西花薊馬已對包括多殺霉素類藥劑在內的至少7類藥劑產生了抗性。多殺霉素是一種大環內酯類生物源農藥(Williamsetal., 2003),主要作用于昆蟲的煙堿型乙酰膽堿受體(nicotinic acetylcholine receptor, nAChR)(Salgado, 1997),GABAAR可能是其次要靶標(Watson, 2001)。已有研究表明西花薊馬等多種昆蟲對多殺霉素的高水平抗性與nAChR的α6亞基變異密切相關(Puineanetal., 2013; Silvaetal., 2016; Wanetal., 2018),但作為靶標之一的GABAAR是否在西花薊馬對多殺霉素抗性形成中起作用尚未見報道。
本研究基于本實驗室西花薊馬的基因組和轉錄組數據(未發表),首先對西花薊馬GABAAR和GABABR基因進行了鑒定、克隆驗證以及生物信息學分析;為探究GABAAR是否參與西花薊馬對多殺霉素的抗性,進一步比較了西花薊馬多殺霉素敏感和抗性品系中GABAAR亞基基因FoRDL,FoLCCH3和FoGRD的序列和表達差異,利用RNAi結合生物測定,研究FoRDL在西花薊馬對多殺霉素抗性中的作用。研究結果將有助于了解西花薊馬GABAR家族基因,為GABAR基因功能研究提供參考,為闡明西花薊馬對多殺霉素的抗性機制提供依據。
1 材料與方法
1.1 供試昆蟲
西花薊馬多殺霉素敏感品系(Ivf03)和抗性近等基因系品系(NIL-R)是通過多世代回交法建立(Yuanetal., 2017)。NIL-R對多殺霉素的抗性倍數為36 000倍(Yuanetal., 2017)。Ivf03使用新鮮四季豆Phaseolusvulgaris飼喂,不接觸任何殺蟲劑。養蟲室溫度為27±1℃,相對濕度為65%,光周期為16L∶8D(Zhangetal., 2007)。NIL-R用2.5%多殺霉素懸浮劑(SC)(spinosad, 美國陶氏益農公司)處理的四季豆飼喂(侯文杰等, 2013),飼養條件與敏感品系一致。
1.2 GABAR家族基因的鑒定
從黑腹果蠅基因組數據庫和GenBank中下載黑腹果蠅和意大利蜜蜂Apismelliferaligustica的GABAR家族基因蛋白序列,并以其為參照,對本實驗室西花薊馬基因組進行GABAR家族基因檢索。然后以本實驗室西花薊馬轉錄組序列為參照,對預測到的西花薊馬GABAR家族基因序列進行校正,同時在NCBI nr數據庫中用BLASTX的方法進行序列比對。根據本實驗室西花薊馬基因組,查詢8個GABAR基因所在染色體及其位置,使用在線工具MG2C(http:∥mg2c.iask.in/mg2c_v2.0/)對西花薊馬GABAR基因進行染色體定位分析。
1.3 GABAR家族基因的克隆及測序
采用Trizol(Trizol?Reagent RNA Extraction Kit, Invitrogen)法分別提取Ivf03和NIL-R品系 3日齡成蟲總RNA,經瓊脂糖凝膠電泳和NanoDrop 2000對RNA的完整性、濃度和純度進行檢測,使用cDNA合成試劑盒(PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit, TaKaRa)分別合成cDNA。根據1.2節獲得的GABAR家族基因序列信息設計引物(表1),根據Q5超保真聚合酶(New England BioLabs)說明書分別進行PCR擴增。 PCR反應體系(25 μL): 5×Q5 Reaction Buffer 5 μL, dNTPs(10 mmol/L)0.5 μL, 正反向引物(10 μmol/L)各1 μL, cDNA模板1 μL, Q5 High-Fidelity DNA Polymerase 0.25 μL, 5×Q5 High GC Enhancer(Optional)5 μL, Nuclease-Free Water 11.25 μL。PCR反應程序: 98℃ 30 s; 98℃ 10 s, 60℃ 30 s, 72℃ 90 s~4 min, 30個循環; 72℃ 10 min。PCR擴增產物送北京擎科新業生物技術有限公司進行測序。
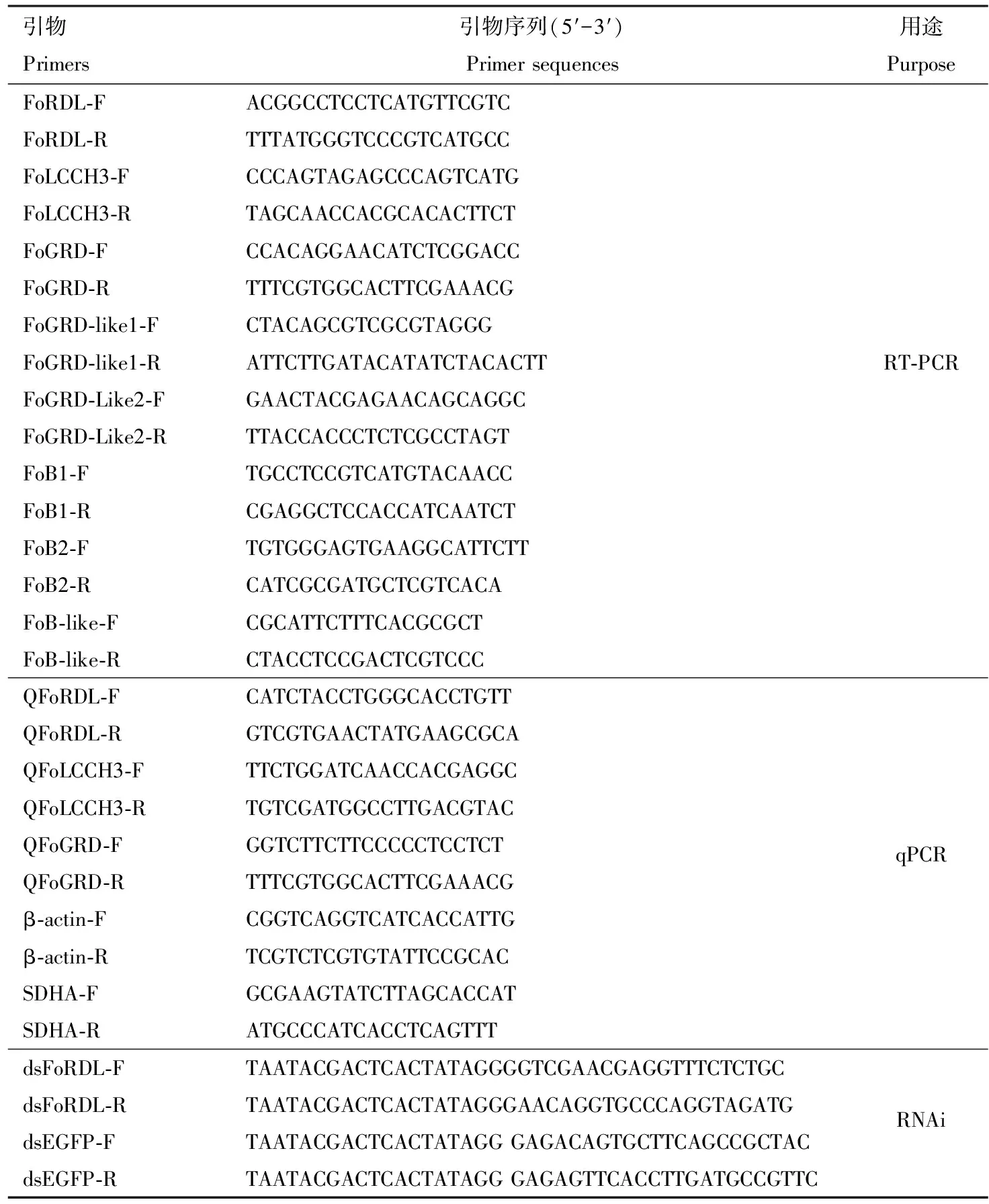
表1 引物信息Table 1 Primer information
1.4 生物信息學分析
采用MEGA6.0軟件鄰接法(neighbor-joining method)(Tamuraetal., 2013)對1.3節克隆驗證的西花薊馬GABAR基因構建系統進化發育樹,bootstrap值設為1 000。通過NCBI數據庫下載膜翅目、鞘翅目、雙翅目、鱗翅目、同翅目和纓翅目等其他物種的GABAR基因,選取黑腹果蠅、二化螟Chilosuppressalis和東亞飛蝗Locustamigratoria的nAChR基因作為外圍參照基因。利用Meme在線程序(http:∥meme-suite.org/tools/meme)對西花薊馬GABAR蛋白保守功能基序(motif)進行分析。使用ExPASy(Gasteigeretal., 2003)和PSORT(Tamura and Akutsu, 2007)工具對西花薊馬的GABAR基因所編碼蛋白質的氨基酸數目、分子量、等電點、帶負電荷的殘基總數(Asp+Glu)、帶正電荷的殘基總數(Arg+Lys)、不穩定指數(instability index)、脂肪族氨基酸指數(aliphatic index)和親疏水性進行分析以及對蛋白的亞細胞定位進行預測。采用DANMAN軟件對Ivf03和NIL-R品系克隆獲得的FoRDL, FoLCCH3和FoGRD基因序列進行比對,同時以已知黑腹果蠅、意大利蜜蜂和赤擬谷盜Triboliumcastaneum相應基因序列為參照。利用軟件ClustalW2(http:∥www.ebi.ac.uk/Tools/msa/clustalw2/)進行基因序列相似性分析,使用在線軟件SignalP(http:∥www.cbs.dtu.dk/services/SignalP/)進行信號肽預測,采用TMHMM軟件(http:∥www.cbs.dtu.dk/services/TMHMM/)進行蛋白質跨膜區域預測。
1.5 GABAAR亞基基因的表達動態
利用qPCR技術測定Ivf03品系不同發育階段(1-2齡若蟲、蛹和3日齡成蟲)和NIL-R品系3日齡成蟲的FoRDL,FoLCCH3和FoGRD相對表達量。在解剖鏡下用解剖針將若蟲、蛹和成蟲分別挑至1.5 mL離心管中,樣品收集后立即置于液氮中冷凍,提取總RNA,cDNA合成方法同1.3節。FoRDL的定量引物參照Meng等(2018),FoLCCH3和FoGRD的定量引物采用Primer Premier 5.0設計(表1)。qPCR在 Applied Biosystems 7500 Real-Time PCR System上進行,反應體系(20 μL): 2×FastFire qPCR PreMix(SYBR Green)10 μL, cDNA模板1 μL, 正反向引物(10 pmol/μL)各0.6 μL, 50×ROX Reference Dye 0.4 μL, RNase-Free ddH2O 7.4 μL。反應程序: 95℃ 10 min; 95℃ 15 s, 60℃ 30 s, 72℃ 30 s, 40個循環。選擇琥珀酸脫氫酶(succinate dehydrogenase, SDHA)和β-肌動蛋白(β-actin)基因作為內參基因(Cifuentesetal., 2012),以兩個內參基因Ct值的幾何平均值校正目的基因的表達水平(Vandesompeleetal., 2002),采用2-ΔΔCt計算基因的相對表達量。每個蟲態設置4個生物學重復,每個生物學重復1-2齡若蟲和蛹約200頭,成蟲約100頭;每個生物學重復設置4個技術重復。
1.6 FoRDL的RNAi
根據1.5節定量測定的結果,FoLCCH3和FoGRD的表達量在敏感和抗性品系中無顯著差異,而FoRDL在抗性品系中表達量顯著低于敏感品系,因此對FoRDL進行了RNAi實驗。使用 T7 RiboMAX Express RNAi試劑盒(Promega, Madison, WI美國),按照說明書合成dsFoRDL(處理組)和dsEGFP(對照組),引物序列見表1。取敏感品系西花薊馬3日齡成蟲約50頭置于干擾裝置內,采用飼喂法進行干擾,具體操作方法參照袁江江等(2018)。分別在干擾12, 24, 36, 48和72 h后收集樣品,檢測沉默效率。處理組和對照組均設置4個生物學重復,每個生物學重復為50頭成蟲;每個重復設置4個技術重復。FoRDL的表達量的檢測方法參照1.5節。采用葉管藥膜法(龔佑輝等, 2010; 王澤華等, 2011)收集處理組和對照組干擾24 h存活的成蟲,使用含有0.1%曲拉通X-100的蒸餾水將2.5%多殺霉素分別配制成0.250和0.400 mg/L兩個濃度,藥劑處理48 h后分別統計并計算處理組和對照組的死亡率。處理組和對照組均設計10個重復,每個重復處理約20頭成蟲。
1.7 數據分析
采用SPSS Statistics 19軟件對實驗數據進行統計分析。采用獨立樣本t檢驗對FoRDL,FoLCCH3和FoGRD在西花薊馬敏感和抗性品系中的表達量以及RNA干擾FoRDL后基因相對表達量和成蟲死亡率差異進行分析;采用單因素方差分析及最小顯著差法(LSD)分析FoRDL,FoLCCH3和FoRGD在不同發育階段的相對表達量差異顯著性。
2 結果
2.1 西花薊馬GABAR家族基因鑒定及生物信息學特征
克隆獲得8個GABAR基因FoRDL,FoLCCH3,FoGRD,FoGRD-like1,FoGRD-like2,FoB1,FoB2和FoB-like(GenBank登錄號: MH148151-MH148158)的全長序列,ORF長度介于1 080~3 720 bp之間,包括5個GABAAR亞基基因和3個GABABR基因,分布于6條染色體上,其中FoLCCH3,FoGRD和FoGRD-like2分布在3號染色體上,FoGRD-like1,FoB2,FoB-like,FoB1和FoRDL依次分布在6, 7, 10, 13和14號染色體上(圖1)。
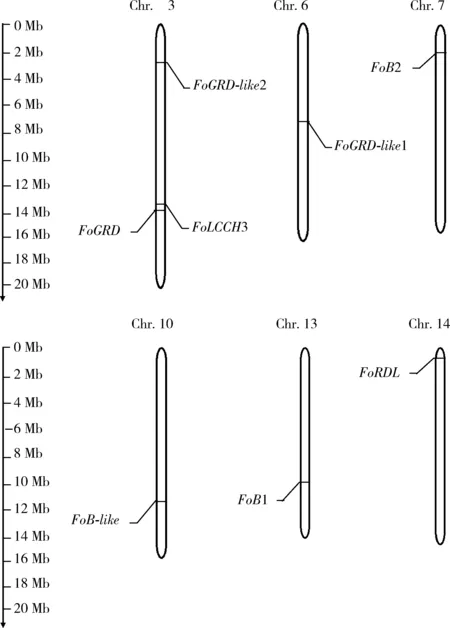
圖1 西花薊馬GABAR基因在染色體上的分布Fig. 1 Distribution of GABAR genes on chromosomes of Frankliniella occidentalis
8個GABAR基因所編碼的氨基酸數目介于360~1 240之間,所編碼的蛋白分子量介于41.88~137.84 kD之間,等電點介于8.04~9.62之間,表明這8個蛋白呈堿性。此外,蛋白帶負電荷的殘基總數范圍為33~138,帶正電荷的殘基總數范圍為44~141,不穩定指數范圍為37.81~49.75,脂肪族氨基酸指數范圍為73.18~97.81;除FoRDL和FoB-like為疏水性蛋白質,其余的均為親水性蛋白質。亞細胞定位預測分析表明,所有GABAR蛋白均分布在內質網。
2.2 西花薊馬GABAR基因系統進化樹
由系統進化分析可知,西花薊馬8個GABAR基因與其他昆蟲物種相對應的GABAR基因聚類,具有很高的保守性。西花薊馬GABAR不同基因的親緣關系不同:對于RDL亞基,西花薊馬所屬的纓翅目與膜翅目昆蟲的親緣關系較近,而與鱗翅目昆蟲的親緣關系較遠(圖2);對于LCCH3亞基,西花薊馬與等翅目白蟻Cryptotermessecundus親緣關系較近,與膜翅目麗蠅蛹集金小蜂Nasoniavitripennis親緣關系較遠(圖2);對于GRD亞基,西花薊馬與雙翅目果蠅Drosophilanavojoa親緣關系較近,FoGRD-like1和FoGRD-like2與鱗翅目玉米螟Ostriniafurnacalis親緣關系較近(圖2);對于B亞基,西花薊馬的B1和B2亞基均與同屬薊馬科的棕櫚薊馬Thripspalmi親緣關系最近(圖2)。
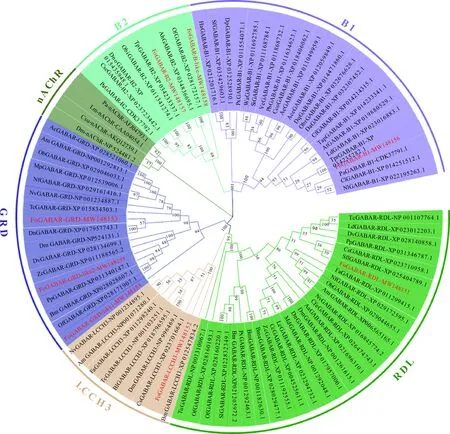
圖2 鄰接法構建的基于氨基酸序列的昆蟲GABAR蛋白系統進化樹(1 000次重復)Fig. 2 Phylogenetic tree of insect GABAR proteins based on amino acid sequences by neighbor-joining method (1 000 replicates)GABAR蛋白來源物種Origin species of GABAR proteins: Fo: 西花薊馬Frankliniella occidentalis; Dm: 黑腹果蠅Drosophila melanogaster; Md: 家蠅Musca domestica; Cc: 實蠅Ceratitis capitata; Zc: 瓜實蠅Zeugodacus cucurbitae; Dn: Drosophila navojoa; Lc: 銅綠蠅 Lucilia cuprina; Tc: 赤擬谷盜Tribolium castaneum; Fa: 阿里山潛蠅繭蜂 Fopius arisanus; Am: 意大利蜜蜂Apis mellifera ligustica; Nv: 麗蠅蛹集金小蜂Nasonia vitripennis; Ob: 雙角壁蜂Osmia bicornis; Bm: 家蠶Bombyx mori; Bma: 野桑蠶Bombyx mandarina; Px: 小菜蛾Plutella xylostella; Sl: 斜紋夜蛾Spodoptera litura; Aa: 黑斑蚊Aedes aegypti; Ld: 馬鈴薯甲蟲Leptinotarsa decemlineata; Cfe: 貓櫛頭蚤Ctenocephalides felis; Of: 亞洲玉米螟Ostrinia furnacalis; Pp: 螢火蟲Photinus pyralis; Dv: 西方玉米根蟲Diabrotica virgifera; Sfl: 蔗黃偽毛蚜Sipha flava; Nf: 黃褐尼氏蟻Nylanderia fulva; Bl: 銹腹實蠅Bactrocera latifrons; Cs: 白蟻Cryptotermes secundus; Ac: 中華蜜蜂Apis cerana; Mp: 法老小家蟻Monomorium pharaonis; Da: 嗜鳳梨果蠅Drosophila ananassae; Tpr: 短管赤眼蜂Trichogramma pretiosum; Sf: 草地貪夜蛾Spodoptera frugiperda; Ha: 茄夜蛾Helicoverpa armigera; Dp: 黑脈金斑蝶Danaus plexippus; Tp: 棕櫚薊馬Thrips palmi; Pa: 美洲大蠊Periplaneta americana; Cf: 多胚跳小蜂Copidosoma floridanum; Cl: 溫帶臭蟲Cimex lectularius; Dq: 方頭恐猛蟻Dinoponera quadriceps; Wa: 金刻沃氏蟻Wasmannia auropunctata; Obr: 鋸針蟻Odontomachus brunneus; Ve: 扁胸切葉蟻Vollenhovia emeryi; Si: 紅火蟻Solenopsis invicta; Tz: 皺切葉蟻Trachymyrmex zeteki; Cco: 駝切葉蟻Cyphomyrmex costatus; Ae: 頂切葉蟻Acromyrmex echinatior; Ace: 六刺芭切葉蟻Atta cephalotes; Ld: 馬鈴薯甲蟲Leptinotarsa decemlineata; At: 小蜂窩甲蟲Aethina tumida; Nl: 稻褐飛虱Nilaparvata lugens; Dno: 汗蜂Dufourea novaeangliae; Ah: 小斑草眼蝶Aphantopus hyperantus; Cse: 白蟻Cryptotermes secundus; Obi: 畢氏卵角蟻Ooceraea biroi; Csu: 畢氏卵角蟻Ooceraea biroi; Lm: 飛蝗Locusta migratoria.
通過motif分析和進化樹聚類結果可知,西花薊馬GABAAR和GABABR氨基酸序列均存在高度一致性。西花薊馬GABAAR亞基FoGRD, FoGRD-like1和FoGRD-like2聚類在一起,FoRDL和FoLCCH3聚類在一起,這5個亞基氨基酸序列的保守基序類型、排序均一致。GABABR亞基FoB1, FoB2和FoB-like氨基酸序列的保守基序一致,但是FoB2在其蛋白質后端又分布4個Motif 8保守基序(圖3)。
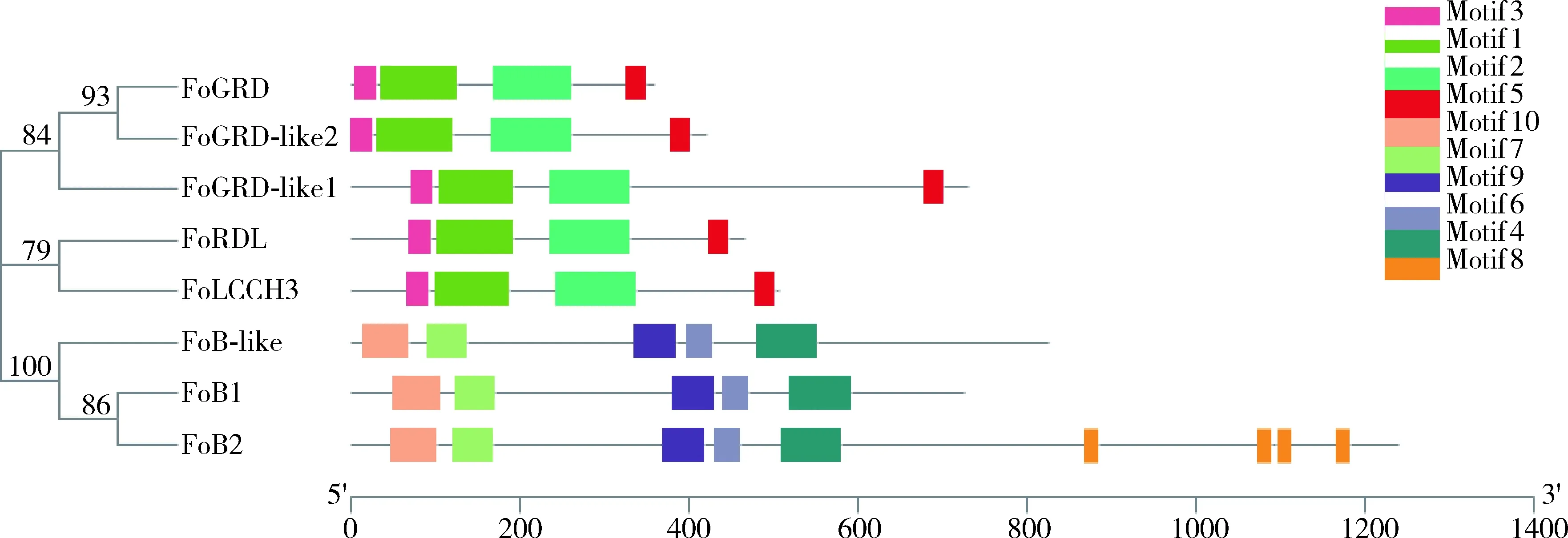
圖3 西花薊馬GABAR基因保守功能基序分析Fig. 3 Analysis of conserved motifs of GABAR genes in Frankliniella occidentalis
2.3 FoRDL, FoLCCH3和FoGRD氨基酸序列比對與結構分析
通過比較Ivf03和NIL-R品系FoRDL, FoLCCH3和FoGRD的氨基酸序列,未發現序列差異。西花薊馬與其他昆蟲的GABAR亞基RDL, LCCH3和GRD氨基酸序列比對分析結果表明,3個GABAAR亞基均屬于半胱氨酸環配體門控離子通道超家族,3個亞基之間具有典型的共同特征,每個亞基均具有6個N端胞外區環結構(loop A-F),以及4個跨膜區(TM 1-4),在不同昆蟲間結構相似度高,且這些環結構、跨膜區的氨基酸序列具有很高的保守性(圖4-6)。此外,RDL在loop D結構的上游存在可變剪切位點(圖4),對Ivf03和NIL-R品系的FoRDL測序結果進行多序列比對發現,在兩個品系中均存在3種形式剪切體,結合實驗室基因組數據表明,此3種形式是由于FoRDL外顯子3的互斥剪切所致。

圖4 西花薊馬FoRDL與其他昆蟲的RDL亞基的氨基酸序列比對Fig. 4 Amino acid sequence alignment of FoRDL of Frankliniella occidentalis and RDL subunits from other insectsRDL亞基來源物種和GenBank登錄號Origin species and GenBank accession numbers of RDL subunits: DmRDL: 黑腹果蠅Drosophila melanogaster, NP_729461; TcRDL: 赤擬谷盜Tribolium castaneum, NP_001107808; AmRDL: 意大利蜜蜂Apis mellifera ligustica, XP_006565167.1. ☆表示相同的氨基酸殘基Showing the identical amino acid residues; ■表示相似的氨基酸殘基Showing the similar amino acid residues.

圖5 西花薊馬FoLCCH3與其他昆蟲的LCCH3亞基的氨基酸序列比對Fig. 5 Amino acid sequence alignment of FoLCCH3 of Frankliniella occidentalis and LCCH3 subunits from other insectsLCCH3亞基來源物種和GenBank登錄號Origin species and GenBank accession numbers of LCCH3 subunits: DmLCCH3: 黑腹果蠅Drosophila melanogaster, NP_996469.1; TcLCCH3: 赤擬谷盜Tribolium castaneum, NP_001103251.1; AmLCCH3: 意大利蜜蜂Apis mellifera ligustica, XP_026298398.1. ☆表示相同的氨基酸殘基Showing the identical amino acid residues; ■表示相似的氨基酸殘基Showing the similar amino acid residues.

圖6 西花薊馬FoGRD與其他昆蟲的GRD亞基的氨基酸序列比對Fig. 6 Amino acid sequence alignment of FoGRD of Frankliniella occidentalis and GRD subunits from other insectsGRD亞基來源物種和GenBank登錄號Origin species and GenBank accession numbers of GRD subunits: DmGRD: 黑腹果蠅Drosophila melanogaster, NP_524131.1; TcGRD: 赤擬谷盜Tribolium castaneum, NP_001107772.1; AmGRD: 意大利蜜蜂Apis mellifera ligustica, NP_001292813.1. ☆表示相同的氨基酸殘基Showing the identical amino acid residues; ■表示相似的氨基酸殘基Showing the similar amino acid residues.
2.4 FoRDL, FoLCCH3和FoGRD在不同齡期的表達模式
FoRDL,FoLCCH3和FoGRD在西花薊馬敏感品系的1-2齡若蟲、蛹和成蟲中均有表達,且均在成蟲中表達量最高。其中FoRDL的表達量隨著西花薊馬的發育而顯著升高(P<0.05)(圖7: A);FoLCCH3在1-2齡若蟲和蛹中表達水平相近,都顯著低于成蟲(P<0.05)(圖7: B);FoGRD在成蟲和蛹中表達水平較高且二者水平相近,在1-2齡若蟲中表達水平相近且均顯著低于蛹或成蟲(P<0.05)(圖7: C)。FoRDL在抗性品系中表達量顯著低于敏感品系(P<0.05)(圖7: D),而FoLCCH3和FoGRD的表達量在兩品系間無顯著差異(P>0.05)(圖7: E, F)。
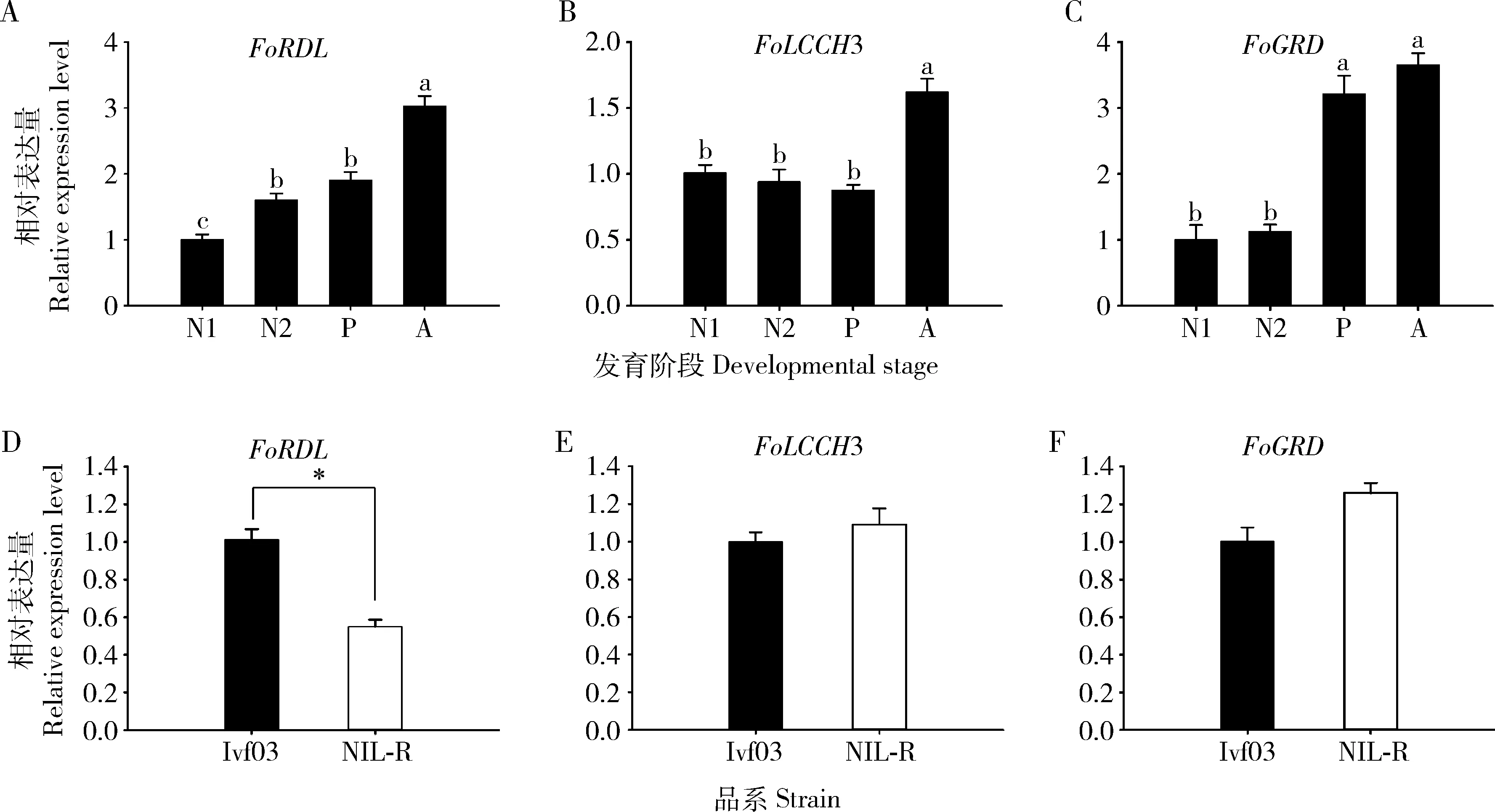
圖7 FoRDL, FoLCCH3和FoGRD在西花薊馬多殺霉素敏感品系Ivf03不同發育階段(A, B和C)及敏感品系Ivf03和抗性品系(NIL-R)成蟲期(D, E和F)的相對表達量Fig. 7 Relative expression levels of FoRDL, FoLCCH3 and FoGRD in the spinosad susceptible strain ofFrankliniella occidentalis (Ivf03) at different developmental stages (A, B and C) and in the Ivf03 strainand the spinosad resistant strain (NIL-R strain) at the adult stage (D, E and F)Ivf03: 西花薊馬多殺霉素敏感品系Spinosad susceptible strain of F. occidentalis; NIL-R: 西花薊馬多殺霉素抗性近等基因系品系Near-isogenic line of the spinosad resistant strain of F. occidentalis; N1: 1齡若蟲1st instar nymph; N2: 2齡若蟲2nd instar nymph; P: 蛹Pupa; A: 成蟲Adult. 圖中柱表示平均值±標準誤;柱上不同字母表示不同發育階段基因表達量顯著性差異(P<0.05, LSD),柱上星號表示品系間差異顯著(P<0.05, 獨立樣本t檢驗)。Data in the figures are mean±SE. Different letters above bars indicate significant difference in the gene expression level (P<0.05, LSD) among different developmental stages, and asterisk above bars indicates significant difference (P<0.05, independent samples t-test) between the strains.
2.5 FoRDL的RNAi及干擾后對多殺霉素的敏感性
西花薊馬敏感品系3日齡成蟲飼喂dsFoRDL12和24 h后,FoRDL的表達量分別下調34.11%和45.16%,與對照組dsEGFP比較差異顯著(P<0.05),在飼喂36, 48和72 h后,FoRDL的表達量無顯著種群差異(P>0.05)(圖8: A)。干擾24 h后,西花薊馬對多殺霉素的敏感性顯著降低,在0.250和0.400 mg/L處理濃度下,其死亡率分別從對照組的38.80%和50.12%顯著降低到17.15%和28.57%(P<0.05),即分別比對照下降了55.80%和43.00%(圖8: B)。
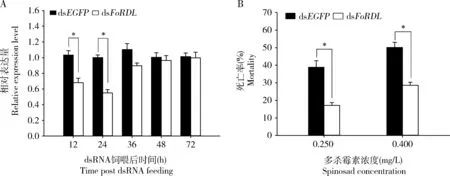
圖8 西花薊馬3日齡成蟲中FoRDL RNAi后FoRDL的相對表達量(A)和FoRDL RNAi后再進行多殺霉素處理后的成蟲死亡率(B)Fig. 8 Relative expression levels of FoRDL after RNAi of FoRDL in the 3-day-old adults of Frankliniella occidentalis (A)and the mortality rate of adults after treatment with spinosad following RNAi of FoRDL (B)dsEGFP: 對照組Control group; dsFoRDL: 處理組Treatment group. 圖中柱表示平均值±標準誤;柱上星號表示差異顯著(P<0.05, 獨立樣本t檢驗)。Data in the figures are mean±SE. Asterisk above bars indicates significant difference (P<0.05, independent samples t-test).
3 討論
本研究通過檢索西花薊馬基因組GABAR家族基因,鑒定到5個GABAAR亞基基因和3個GABABR基因;西花薊馬FoRDL,FoLCCH3和FoGRD編碼的氨基酸序列具有昆蟲GABAAR亞基的典型結構;FoRDL外顯子3存在著3種互斥剪切,它們可以形成3種不同的剪接體。RDL亞基的選擇性剪切存在于多種昆蟲中,黑腹果蠅體內的RDL亞基,外顯子3和外顯子6可通過選擇性剪接編碼形成4個不同的多態片段(Ffrench-Constant and Rocheleau, 1993)。在家蠶、意大利蜜蜂、赤擬谷盜和麗蠅蛹集金小蜂等昆蟲中都存在RDL亞基外顯子3和外顯子6的不同的剪接形式(Jones and Sattelle, 2008; Jonesetal., 2010; Yuetal., 2010)。mRNA選擇性剪切豐富了生物進化的方式,是形成蛋白質結構和功能多樣性的重要機制,然而對昆蟲RDL亞基不同剪接體的功能有待進一步研究。
昆蟲的GABAAR作為殺蟲劑的重要作用靶標,與乙酰膽堿酯酶、鈉離子通道類似,一直是毒理學研究的熱點。昆蟲的GABAR主要存在于中樞神經系統和神經肌肉連接處(Bloomquist, 2001),介導神經和肌肉細胞中快速的抑制性突觸傳遞。西花薊馬GABAAR亞基基因FoRDL,FoLCCH3和FoGRD在敏感品系的各個發育階段均有表達。雖然3個基因的表達模式有所不同,但是隨著齡期的增長,表達量均呈現升高的趨勢(圖7)。Wei等(2015)研究發現,灰飛虱Laodelphaxstriatellus的RDL,GRD和LCCH3在其整個發育階段均有存在且隨著齡期的增長,相對表達量有著先降低后升高的趨勢,表明GABAAR亞基基因在不同昆蟲中表達模式有所不同。此外,GABAAR亞基基因在昆蟲不同組織部位表達量也存在較大差異。小菜蛾和甜菜夜蛾兩種昆蟲頭部的GABAAR亞基基因相對表達量顯著高于胸部和腹部(周小毛等, 2006; Shangetal., 2009),灰飛虱頭部GABAAR亞基基因的表達量也顯著高于胸部、腹部和足部(Weietal., 2015)。這可能與昆蟲GABAAR主要存在于昆蟲中樞神經系統有關。
對于昆蟲GABAR在抗藥性形成中的作用研究表明,黑腹果蠅對狄氏劑產生抗性與GABAR的RDL亞基A2′S和A2′G突變有關(Leeetal., 1993)。且在抗狄氏劑等環戊二烯類殺蟲劑的德國小蠊Blattellagermanica、赤擬谷盜、小菜蛾等昆蟲的RDL亞基中均發現了類似黑腹果蠅的A2′S和A2′G突變(Miyazakietal., 1995; Yuanetal., 2010; Gondhalekar and Scharf, 2012)。灰飛虱對氟蟲腈產生抗性與GABAAR RDL高頻率的A2′N突變有關(Nakaoetal., 2011)。Wei等(2015)對灰飛虱氟蟲腈敏感品系和抗性品系的GABAAR的RDL亞基進行基因沉默后,使其對氟蟲腈的敏感性分別降低了30%和27%,驗證了RDL亞基作為氟蟲腈靶標受體的功能。本研究比較了西花薊馬多殺霉素敏感和抗性品系的GABAAR亞基(FoRDL, FoLCCH3和FoGRD)氨基酸序列,未發現這些亞基在敏感和抗性品系中存在序列差異,但FoRDL的表達量在抗性品系中顯著低于敏感品系(圖7)。同時研究結果發現,干擾西花薊馬敏感品系FoRDL后進行生物測定,其成蟲死亡率顯著降低(圖8),由此推測西花薊馬GABAAR 的RDL 亞基可能與多殺霉素抗性有關。
目前昆蟲對多殺霉素的抗性機制主要包括細胞色素P450s、多功能氧化酶(MFO)介導的代謝增強(Wangetal., 2006; Reyesetal., 2012; Rehan and Freed, 2014)和nAChR α6亞基的變異(Puineanetal., 2013; Silvaetal., 2016; Wanetal., 2018)。GABAR是多殺霉素的作用靶標之一,本研究表明西花薊馬GABAAR的RDL亞基基因可能參與了其對多殺霉素的抗性形成。這為我們深入了解西花薊馬對多殺霉素的抗性機制,及為西花薊馬的抗性治理和科學防控提供了理論依據。
