家蠶BmlncR2036調控P25基因啟動子活性及參與中腸發育調控
2021-09-06 05:39:00霍春月常美玲闞云超李丹丹
昆蟲學報 2021年8期
關鍵詞:檢測
霍春月, 程 浩, 付 裕, 常美玲, 王 藝, 闞云超, 李丹丹
(南陽師范學院, 河南省伏牛山昆蟲生物學重點實驗室, 河南南陽 473061)
長鏈非編碼RNA (long non-coding RNA, lncRNA)是長度大于200 nt不編碼蛋白質的一大類RNA。lncRNA依據其在基因組上的位置可分為五大類(Jarrouxetal., 2017):反義lncRNA (Katayamaetal., 2005; Heetal., 2008; Faghihi and Wahlestedt, 2009; Weryetal., 2018)、雙向lncRNA(Hamazakietal., 2017)、增強子相關lncRNA (Kimetal., 2010; Wangetal., 2011; Dingetal., 2018)、基因間lncRNA(Guttmanetal., 2009; Cabilietal., 2011; Ulitskyetal., 2011; Ulitsky and Bartel, 2013)和假基因lncRNA (Balakirev and Ayala, 2003; Pinketal., 2011; Tutar, 2012)。lncRNA能夠在表觀遺傳學水平調控基因表達,參與生物體生長、發育、生殖、分化等過程(Engreitzetal., 2016b; Kim and Shiekhattar, 2016; Robinsonetal., 2020),如:改變DNA甲基化、組蛋白修飾狀態(Akhadeetal., 2017; Dykes and Emanueli, 2017),作為酶的輔因子調控轉錄因子活性,與編碼蛋白質的mRNA形成雙鏈雜交體干預mRNA的剪切,參與調節mRNA穩定性(Engreitzetal., 2016a),作為內源競爭RNA (competing endogenous RNAs, ceRNA)與miRNA競爭性結合mRNA,作為小分子RNA的前體(Tayetal., 2014)等。最近研究表明,一些lncRNA能夠編碼小肽,表明它們可能兼具肽編碼能力和非編碼RNA功能(Rohrigetal., 2002; Andersonetal., 2015; Nelsonetal., 2016; Legninietal., 2017; Matsumotoetal., 2017; Razookyetal., 2017; Linetal., 2020; Wuetal., 2020; 趙念念等, 2020)。
家蠶Bombyxmori是鱗翅目的模式昆蟲,絲腺作為重要的產絲場所,在其經濟價值實現中具有重要作用。蠶絲蛋白由絲素和絲膠兩種成分組成,絲素在后部絲腺合成,編碼絲素蛋白的基因主要有Fib-L,Fib-H和P25;絲膠在中部絲腺分泌,編碼絲膠蛋白的基因主要有Ser1,Ser2和Ser3 3種 (Yamaguchietal., 1989; Inoueetal., 2000; Asakuraetal., 2007)。宋菲等(2014)發現miR-2739對絲素重鏈基因Fib-H的表達具有調控作用;Huang等(2011)發現miR-2b與P25基因在表達上存在著相反的關系,可能調控P25基因的表達。另外Qian等(2018, 2020)發現miR-2780a和miR-275能夠特異性存在于中部絲腺中,靶向調控絲膠蛋白基因Ser1和Ser2的表達。長鏈非編碼RNA作為一種重要的調控因子,在家蠶變態發育及抗病毒過程中發揮重要作用,目前在家蠶中已發現10 000余條lncRNA序列(Wuetal., 2016; Zhouetal., 2016, 2018; Dykes and Emanueli, 2017; Yinetal., 2020)。來源于家蠶性別決定基因dsx反義鏈上的lncRNA Bmdsx-AS1能夠參與dsx基因的選擇性剪接(Xuetal., 2019)。感染核型多角體病毒(nuclear polyhedrosis virus, NPV)后,家蠶中4 450個lncRNAs出現顯著差異表達,其中上調表達2 837個和下調表達1 613個,表明lncRNA參與了家蠶病毒感染過程(Yinetal., 2020)。另外,lncRNA可作為miRNA的ceRNA發揮調控作用,Wu等(2016)從家蠶基因組中獲得11 810個lncRNA,其中104個lncRNA具有內源競爭性RNA的潛力,可能作為ceRNA發揮作用。
在前期研究中我們構建了家蠶4齡眠和5齡第3天幼蟲絲腺非編碼RNA文庫,發現了大量lncRNA在這兩個齡期的絲腺中存在顯著差異表達,其中在5齡第3天幼蟲絲腺中高表達的一個lncRNA BmlncR2036位于絲素基因P25上游,可能參與P25基因的調控。為進一步探索BmlncR2036在家蠶絲腺及變態發育中的功能,本研究對其在家蠶幼蟲不同組織和不同發育階段絲腺中的表達譜進行了鑒定,同時對BmlncR2036作為ceRNA的功能和參與P25基因啟動子調控的機制進行了初步研究,最后在家蠶個體中干涉BmlncR2036表達,對其參與家蠶絲腺及中腸發育調控的機制進行了初步探索,為進一步研究lncRNA功能提供了實驗證據。
1 材料與方法
1.1 試蟲來源
家蠶品系為大造P50,由中國農業科學院蠶業研究所李木旺研究員惠贈。
1.2 家蠶飼養及取樣
家蠶飼養在溫度25℃,相對濕度60%,光周期12L∶12D,用新鮮桑葉飼養。取5齡第3天家蠶幼蟲,解剖后收集體壁、腦、神經、精巢、卵巢、絲腺、馬氏管、血淋巴、脂肪體和中腸10種組織,分別置于液氮中速凍保存。取4齡第3天幼蟲、4齡眠和5齡第1-2天幼蟲,解剖后取整個絲腺速凍于液氮中;取5齡第3-5天幼蟲及5齡熟蠶解剖蟲體,將絲腺分為前絲腺、中絲腺和后絲腺3個部分,分別置于液氮中速凍保存。
1.3 qPCR檢測BmlncR2036的表達譜
在前期構建的家蠶4齡眠和5齡第3天絲腺非編碼RNA文庫中,發現一個高表達的lncRNA BmlncRNA2036位于P25上游9.47 kb的互補鏈上,轉錄方向與P25相反(王立方, 2015)。BmlncR2036及P25基因表達譜檢測同楊宗霖等(2019)方法:將1.2節各樣品用TRIzol法提取總RNA,取總RNA 2 μg使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit(TaKaRa)合成cDNA第1鏈。基于內參基因actin, BmlncR2036及P25基因序列用Primer Premier 5.0軟件設計引物,PCR反應體系: cDNA 0.5 μL, 上下游引物終濃度為0.5 μmol/L, FastStart Universal SYBR Green Mix (Roche) 5 μL。PCR反應條件: 95℃變性30 s, 5℃退火30 s, 72℃延伸30 s, 40個循環。所用引物及miRNA模擬物見表1。實驗設置生物學重復3次,每個樣品技術重復3次。
1.4 靶向P25基因和BmlncR2036的miRNA預測
使用miRanda(Enrightetal., 2003)和RNAhybrid(Rehmsmeieretal., 2004)軟件預測能夠同時靶向P25基因和BmlncR2036的miRNA。
1.5 熒光素酶檢測法檢測miRNA與P25基因的體外互作
P25基因3′UTR熒光素酶報告基因載體psiCHECK-2[P25]構建同楊宗霖等(2019)。以家蠶5齡幼蟲后絲腺RNA為模板,使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit (TaKaRa)合成cDNA第1鏈,擴增P25基因3′UTR 413 bp序列,引物序列見表1。PCR反應體系(20 μL): cDNA 1 μL, 上下游引物終濃度為0.5 μmol/L, Premix Ex TaqTMHot Start Version (TaKaRa) 10 μL。PCR反應條件: 95℃變性30 s, 55℃退火30 s, 72℃延伸1 min, 30個循環。使用瓊脂糖凝膠DNA回收試劑盒(Axygen)回收目的條帶,連接pMD19-T Simple載體(TaKaRa)。轉化后,菌液PCR檢測連接效果,送菌液至蘇州泓迅生物科技股份有限公司測序。測序正確的菌液提質粒,用XhoⅠ和NotⅠ雙酶切后連接到psiCHECK-2熒光素酶報告基因載體(Promega)上。經轉化、菌液PCR檢測、測序等步驟確定載體是否構建成功。測序正確的菌液重新搖菌,用去內毒素質粒提取試劑盒(Omega)提取質粒。
基于miR-2739和miR-279a成熟miRNA序列設計引物,由上海吉瑪制藥技術有限公司合成miRNA mimics和陰性對照(NC),引物序列見表1。
細胞轉染及熒光素酶測定方法同楊宗霖等(2019)。人胚胎腎細胞HEK293T細胞(由衡陽師范學院唐青海博士惠贈)在含有10%胎牛血清(Gibco)的DMEM培養基(Biological Industry)中培養瓶傳代培養,溫度37℃,CO2濃度5%。按每孔2×105個細胞鋪24孔板,使用X-tremeGENE siRNA Transfection Reagent(Roche)轉染miRNA mimics和P25基因熒光素酶報告基因載體。miRNA mimics轉染濃度為100 nmol/孔,P25基因熒光素酶報告基因載體質粒轉染濃度為100 ng/孔。實驗設置5組:無任何處理的HEK293T細胞;轉染psiCHECK-2[P25]空載體;共轉染psiCHECK-2[P25]與miR-2739 mimics;共轉染psiCHECK-2[P25]與NC mimics;共轉染psiCHECK-2[P25]與miR-279amimics。每組設樣品重復3個,實驗重復3次。轉染48 h后,用細胞裂解液裂解細胞,取20 μL細胞裂解液按照雙熒光素檢測試劑盒Steady-Glo? Luciferase Assay System(Promega)步驟,檢測樣品熒光值。P25基因位于Firefly熒光素酶基因下游,因此用獲得的Firefly熒光素酶活性(F)/內參Renilla熒光素酶活性(R),獲得每組3個樣品及3個重復的R/F值,均一化后用Excel軟件進行作圖(楊宗霖等, 2019)。
1.6 P25基因啟動子活性檢測
BmlncR2036位于P25基因上游,那么其可能通過影響P25基因啟動子活性參與P25基因表達調控。取家蠶5齡熟蠶后絲腺,用DNAzol(MRC)提取基因組DNA,參照Nony等(1995)方法,使用表1引物擴增P25基因上游1 251 bp序列,構建P25基因啟動子區熒光素酶報告基因載體。PCR反應體系及反應條件同1.5節,目的條帶回收,連接pMD19-T Simple載體,測序,測序正確的菌液提質粒,用KpnⅠ和HindⅢ雙酶切后連接在pGL3-Enhancer載體(Promega)上,構建P25基因啟動子載體pGL3-Enhancer[P25-promoter],經轉化、菌液PCR檢測、測序等步驟確定載體是否構建成功。測序正確的菌液重新搖菌,用去內毒素質粒提取試劑盒(Omega)提取質粒備用。
取家蠶5齡熟蠶后絲腺RNA反轉錄cDNA,使用表1引物擴增BmlncR2036基因全長704 bp序列。PCR反應體系及反應條件同1.5節,目的條帶回收,連接pMD19-T Simple載體,測序,測序正確的菌液提質粒,用KpnⅠ和EcoRⅠ雙酶切后連接在pcDNA3.1(+)載體(Promega)上構建BmlncR2036過表達載體pcDNA3.1(+)[BmlncR2036],經轉化、菌液PCR檢測、測序等步驟確定載體是否構建成功。測序正確的菌液重新搖菌,用去內毒素質粒提取試劑盒(Omega)提取質粒備用。
HEK293T細胞培養及鋪板同1.5節,實驗設置2組:共轉染pGL3-Enhancer [P25-promoter]和pcDNA3.1(+)空載體組;共轉染pGL3-Enhancer [P25-promoter]和pcDNA3.1(+)[BmlncR2036]組。每組設樣品重復3個,實驗重復3次。轉染48 h后,取100 μL細胞,按照Steady-Glo? Luciferase Assay System (Promega)步驟檢測螢火蟲熒光素酶活性,檢測BmlncR2036對P25基因啟動子活性的影響,P25基因啟動子連接在螢火蟲熒光素酶基因上游,因此比較獲得的每組3個樣品及3個重復的Firefly熒光素酶活性(F),均一化后用Excel軟件進行比較作圖。
1.7 RNAi干涉BmlncR2036
取家蠶5齡熟蠶后絲腺RNA,使用表1帶有T7啟動子的引物擴增BmlncR2036基因351 bp片段,PCR反應體系(20 μL): cDNA 1 μL, 上下游引物終濃度為0.5 μmol/L, Premix TaqTM(Ex TaqTMVersion 2.0, TaKaRa) 10 μL。PCR反應條件: 95℃變性30 s, 55℃退火30 s, 72℃延伸30 s, 30個循環。用PCR產物純化試劑盒(Axygen)純化目的片段,用T7體外轉錄試劑盒(Promega)轉錄合成dsRNA,體系(20 μL):帶T7啟動子的BmlncR2036基因PCR產物0.5~1 μg, 2×T7 Buffer 10 μL, Enzyme Mix 2 μL。37℃反應30 min,70℃ 10 min,室溫20 min使反轉錄的RNA形成雙鏈。DNase (Thermo Fisher) 37℃ 30 min去除模板DNA。體外轉錄dsRNA用等體積的異丙醇進行濃縮純化。陰性對照為EGFP基因,以piggyBac[EGFP]載體為模板擴增攜帶有T7啟動子的EGFP基因,其dsRNA合成方法同上。
選擇家蠶5齡第3天幼蟲,利用顯微注射針將dsRNA沿背部血淋巴注射入家蠶體內,每頭注射5 μg,每個基因注射3組,每組5頭。48 h后取家蠶個體,解剖蟲體,除去腸道內容物,將蟲體分為絲腺和非絲腺兩部分,液氮研磨,TRIzol法提取RNA,qPCR檢測dsRNA干涉效果。qPCR檢測同楊宗霖等(2019)方法:分別取注射BmlncR2036和EGFPdsRNA的絲腺或非絲腺樣品總RNA 2 μg,使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit(TaKaRa)合成cDNA第1鏈。內參基因為actin。PCR反應體系(10 μL): cDNA 0.5 μL, 上下游引物終濃度為0.5 μmol/L, FastStart Universal SYBR Green Mix (Roche) 5 μL。PCR反應條件: 95℃變性30 s, 55℃退火30 s, 72℃延伸30 s, 40個循環。所用引物見表1。qPCR實驗設置生物學重復3次,每樣品技術重復3次。未取樣幼蟲飼養至羽化,發現長勢緩慢或形態異常幼蟲,解剖蟲體,觀察RNAi處理后家蠶絲腺及中腸形態及內容物的變化。
1.8 數據分析
數據分析同楊宗霖等(2019)方法。PCR結果采用2-ΔΔCt法計算相對表達量高低,Ct為循環閾值,表示每一個反應管內的熒光信號到達設定的閾值時所經歷的循環數,ΔCt=目的基因的平均Ct值-內參基因的平均Ct值,ΔΔCt=ΔCt處理樣本-ΔCt對照樣本。實驗獨立生物學重復3次,每樣品測定技術重復3次,數據為3次獨立生物學實驗平均值分析結果。數據采用SPSS Statistics 22.0軟件進行處理,以平均值±標準誤表示,并采用單因素方差分析和t檢驗檢測不同處理間的差異顯著性。
2 結果
2.1 BmlncR2036在家蠶幼蟲不同組織及不同發育階段絲腺中的表達譜
qPCR檢測結果表明BmlncR2036在家蠶5齡第3天幼蟲和5齡熟蠶的后絲腺中高表達(圖1: B),與P25基因在后絲腺高表達(Zhongetal., 2005; Maetal., 2013)呈現較一致的表達趨勢。BmlncR2036在家蠶5齡第3天幼蟲不同組織中的表達譜表明,BmlncR2036除在精巢和中腸中高表達外,在其他組織中表達量較低(圖1: A),表明lncRNA BmlncR2036可能具有多樣化的功能。
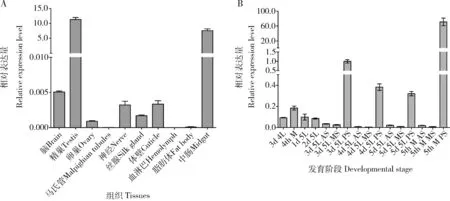
圖1 qPCR檢測BmlncR2036在家蠶5齡幼蟲不同組織(A)和不同齡期絲腺(B)中的表達譜Fig. 1 Expression profiles of BmlncR2036 in different tissues of the 5th instar larvae (A) and silk glandsat different developmental stages (B) of Bombyx mori by qPCR3d 4L: 4齡第3天幼蟲Day-3 4th instar larva; 4th M: 4齡眠4th instar molting larva; 1-5d 5L: 分別為5齡第1-5天幼蟲Day-1-5 5th instar larvae, respectively; AS: 前絲腺Anterior silk gland; MS: 中絲腺Middle silk gland; PS: 后絲腺Posterior silk gland; 5th M: 5齡熟蠶Mature 5th instar larva. 4齡第3天、4齡眠和5齡第1-2天幼蟲取整個絲腺。The whole silk gland was sampled from the day-3 4th instar larvae, 4th instar molting larvae and day-1-2 5th instar larvae. 圖中數據為平均值±標準誤。Data in the figure are mean±SE.
2.2 靶向P25基因和BmlncR2036的miRNA
預測結果表明,在靶向P25基因的miRNA中,能與BmlncRNA2036匹配的主要有miR-2739和miR-279a。為進一步驗證P25基因與兩個miRNA是否存在互作,我們利用熒光素酶檢測系統體外驗證二者的互作。首先構建靶標基因P25 3′UTR區熒光素酶報告基因過表達載體psiCHECK-2[P25],P25 3′UTR擴增結果如圖2(A)所示,PCR產物電泳條帶單一(條帶1),413 bp位置正確。psiCHECK-2[P25]雙酶切結果為條帶2,能夠酶切出正確的P25 3′UTR條帶,測序結果正確。共轉染miR-2739和miR-279a模擬物和P25 3′UTR過表達載體psiCHECK-2[P25]后,細胞熒光素酶活性與NC相比無顯著變化 (圖2: B),表明P25不是miR-2739和miR-279a的真實靶標。
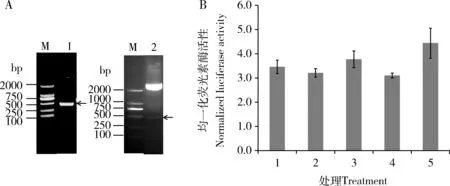
圖2 熒光素酶檢測系統體外驗證P25基因與miRNAs的互作Fig. 2 Verification of the interaction between P25 gene and miRNAs by luciferase assay in vitroA: P25 3′UTR PCR擴增結果及psiCHECK-2[P25]酶切結果PCR amplification result of P25 3′UTR and the double digestion result of psiCHECK-2[P25]; M: DL2000 marker; 1: P25 3′UTR擴增產物 Amplification product of the 3′UTR of P25 gene; 2: 載體psiCHECK-2[P25]雙酶切結果Double digestion result of psiCHECK-2[P25] vector. B: HEK293細胞系中熒光素酶活性實驗檢測miR-2739和miR-279a與靶基因P25的互作Verification of interaction between target gene P25 and miR-2739 and miR-279a by luciferase assay in HEK293 cells; 1: 無任何處理的HEK293細胞對照The normal HEK293 cells as the control; 2: 轉染psiCHECK-2[P25]載體Transfected with psiCHECK-2[P25]; 3: 共轉染psiCHECK-2[P25]+miR-2739 mimics Cotransfected with psiCHECK-2[P25] and miR-2739 mimics; 4: 共轉染psiCHECK-2[P25]+NC mimics Cotransfected with psiCHECK-2[P25] and NC mimics; 5: 共轉染psiCHECK-2[P25]+miR-279a mimics Cotransfected with psiCHECK-2[P25] and miR-279a mimics. 圖中數據為平均值±標準誤。Data in the figure are mean±SE.
2.3 BmlncR2036對P25啟動子活性的影響
P25啟動子序列和BmlncR2036擴增結果如圖3(A)所示,PCR產物電泳條帶單一,位置正確。pGL3-Enhancer [P25-promoter]和pcDNA3.1(+) [BmlncR2036]雙酶切結果如圖3(B),能夠酶切出正確的P25啟動子條帶和BmlncR2036全長,測序結果正確。在HEK293T細胞中共轉染pGL3-Enhancer [P25-promoter]和pcDNA3.1(+) [BmlncR2036]后,細胞中BmlncR2036實現顯著過表達(圖3: C)。細胞熒光素酶活性比單獨轉染pGL3-Enhancer [P25-promoter]細胞的下降了52% (圖3: D),達極顯著水平(P<0.01),表明BmlncR2036對P25啟動子區存在負調控作用。
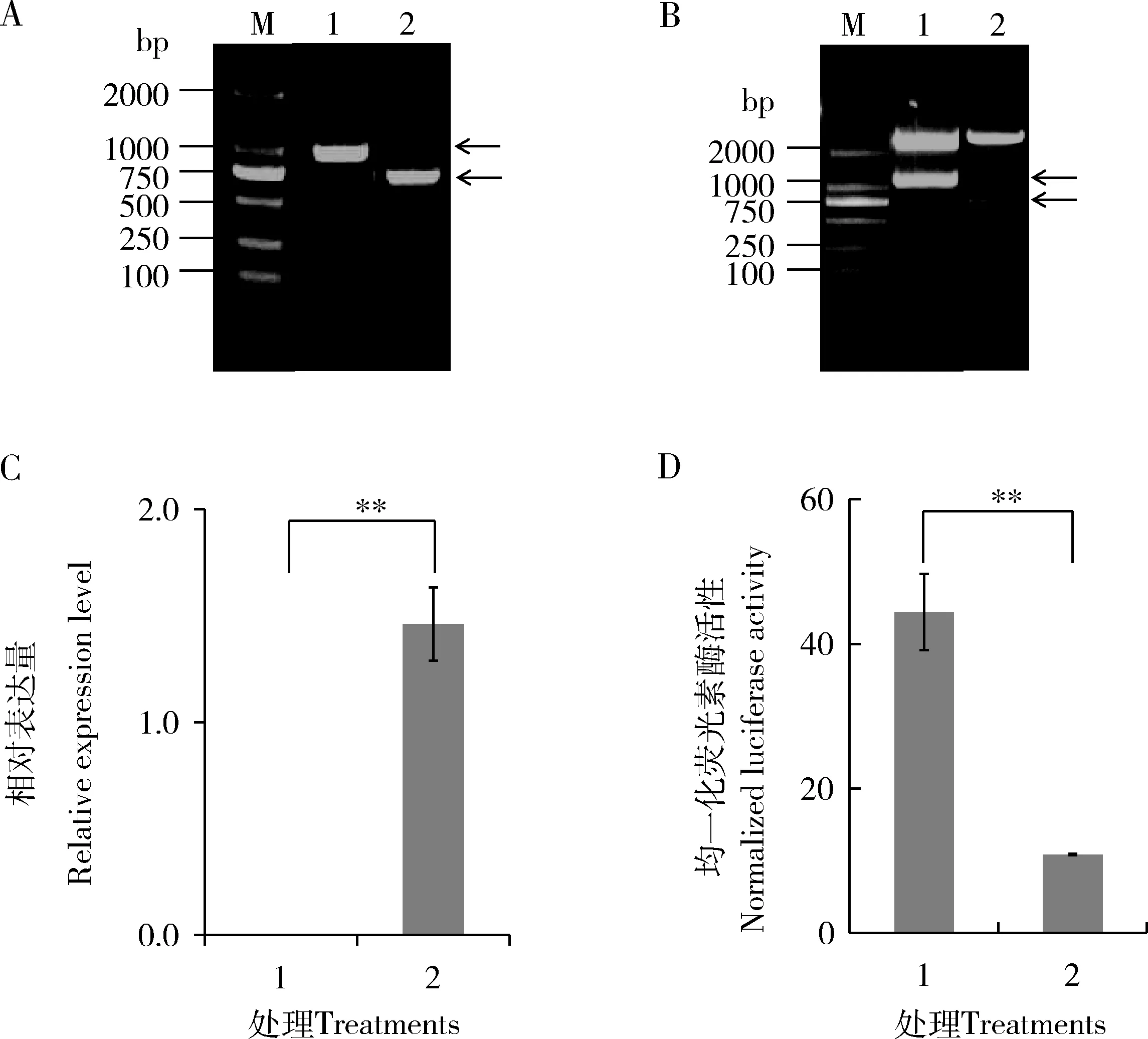
圖3 BmlncR2036和P25啟動子互作結果Fig. 3 Interaction result of BmlncR2036 and the promoter of P25A: P25啟動子和BmlncR2036全長PCR擴增結果 PCR result of P25 promoter and the full-length of BmlncR2036; M: DL2000 marker; 1: P25 啟動子擴增產物 Amplification product of the promoter of P25 gene; 2: BmlncR2036全長擴增產物 Amplification product of the full-length of BmlncR2036. 箭頭示PCR擴增產物Arrows show PCR products. B: 載體pGL3-Enhancer [P25-promoter]和pcDNA3.1(+)[BmlncR2036]雙酶切鑒定Double digestion of vectors pGL3-Enhancer [P25-promoter] and pcDNA3.1(+)[BmlncR2036]; M: DL2000 marker; 1: pGL3-Enhancer [P25-promoter]雙酶切產物Double digestion product of pGL3-Enhancer [P25-promoter]; 2: pcDNA3.1(+)[BmlncR2036]雙酶切產物Double digestion product of pcDNA3.1(+)[BmlncR2036]. 箭頭分別示酶切獲得的P25啟動子序列 1 251 bp和BmlncR2036 704 bp全長序列Arrows show the digested 1 251 bp P25 promoter sequence and 704 bp BmlncR2036 full-length sequence, respectively. C: HEK293細胞系中BmlncR2036過表達結果Overexpression result of BmlncR2036 in HEK293 cells; D: HEK293細胞系中熒光素酶活性實驗檢測BmlncR2036對P25基因啟動子活性影響 Verification of the effect of BmlncR2036 on the promoter activity of P25 by detection of the luciferase activity in HEK293 cells; 1: 轉染pGL3-Enhancer [P25-promoter]載體和pcDNA3.1(+)空載體Transfected with vector pGL3-Enhancer [P25-promoter] and blank vector pcDNA3.1(+); 2: 轉染載體pGL3-Enhancer [P25-promoter]和pcDNA3.1(+)[BmlncR2036] Transfected with vectors pGL3-Enhancer [P25-promoter] and pcDNA3.1(+)[BmlncR2036]. 圖中數據為平均值±標準誤;柱上雙星號表示差異極顯著(P<0.01, 單因素方差分析, t檢驗)。Data in the figure are mean±SE. Double asterisk above bars represents extremely significant difference (P<0.01, one-way ANOVA, t-test). 下同The same below.
2.4 RNAi干涉家蠶幼蟲中BmlncR2036表達
在家蠶5齡第3天幼蟲血淋巴中注射BmlncR2036 dsRNA發現,48 h后BmlncR2036在非絲腺組織中的表達量下降了84%,絲腺組織中表達量無明顯變化(圖4: A)。注射BmlncRNA2036 dsRNA的家蠶個體發育遲緩,上簇后體色變暗,后逐漸死亡(圖4: B),解剖后發現,中腸有大量內容物殘留,馬氏管顏色發白,但絲腺無明顯異常(圖4: C),表明BmlncR2036可能在家蠶中腸發育過程中發揮調控作用。
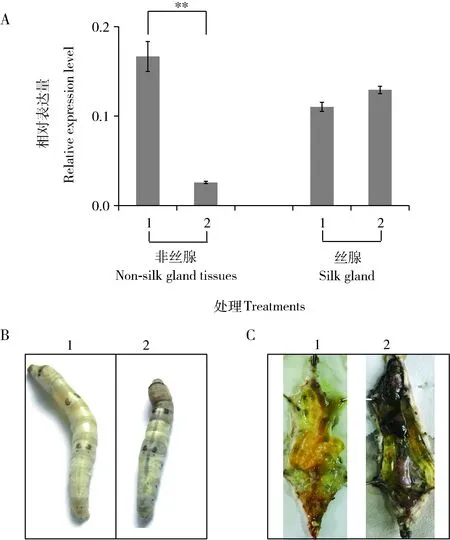
圖4 家蠶5齡幼蟲中通過RNAi敲低BmlncR2036表達后的結果Fig. 4 Result of knocking down the expression of BmlncR2036 in the 5th instar larvae of Bombyx mori by RNAiA: 注射dsRNA后不同組織中BmlncR2036表達結果 Expression result of BmlncR2036 in different tissues after dsRNA injection; B: 注射BmlncR2036 dsRNA后家蠶表型Phenotype of B. mori after dsRNA injection of BmlncR2036; C: 注射BmlncR2036 dsRNA后家蠶解剖結果Dissection result of B. mori after injection of BmlncR2036 dsRNA. 1: 注射EGFP dsRNA的對照Injection of EGFP as the control; 2: 注射BmlncR2036 dsRNA Injection of BmlncR2036 dsRNA.
3 討論
我們在前期研究中從家蠶幼蟲絲腺中鑒定到大量新的長鏈非編碼RNA,并對其在絲腺發育過程的表達譜進行了研究(王立方, 2015),發現一個在絲素P25基因上游存在的lncRNA BmlncR2036在家蠶后絲腺中高表達。BmlncR2036在幼蟲的后絲腺中高表達(圖1: B),已知P25基因也在幼蟲尤其是5齡幼蟲后絲腺中富集(Zhongetal., 2005; Maetal., 2013),表明其可能作為P25的競爭性內源RNA發揮作用。通過分析其作為內源性競爭miRNA發現,同時靶向BmlncR2036和P25基因的miRNAmiR-2739和miR-279a并不能在體外與P25基因產生互作(圖2: B),表明BmlncR2036可能不是通過作為miR-2739和miR-279a這兩個miRNA的ceRNA對P25發揮作用,即miR-2739和miR-279a可能有其他靶標,而P25也有可能存在其他BmlncR2036起ceRNA作用的miRNA分子,具體機制還需進一步實驗驗證。
分析BmlncR2036對P25基因啟動子活性影響發現,BmlncR2036對P25基因啟動子的轉錄存在負調控(圖3: C, D)。基因上下游存在為數眾多的lncRNA,包括增強子RNA,啟動子相關lncRNA,上游反義鏈RNA,啟動子上游lncRNA,端粒重復區lncRNA等(St Laurentetal., 2015)。Wang等(2015)在骨骼肌成肌細胞中鑒定出一個位于發育多能性相關基因2 (developmental pluripotency-associated 2,Dppa2)上游的發育多能性相關基因2上游肌相關lncRNA (developmental pluripotency-associated 2 upstream binding muscle lncRNA, lncRNA-Dum),該lncRNA能夠順式募集DNA甲基轉移酶1(DNA methyltransferase1, DNMT1), DNMT3A和DNMT3B等至Dppa2基因的啟動子區,并造成該區域甲基化沉默,從而抑制Dppa2的表達,造成骨骼肌成肌細胞向肌細胞分化。BmlncR2036是位于P25基因上游的lncRNA,其是否具有類似的機制,還需進一步垂釣能夠與BmlncR2036結合的蛋白,判斷其是否通過募集蛋白至P25基因的啟動子區從而參與P25基因的表達調控。因此對BmlncR2036功能的探索還需更多的實驗證據。
個體中RNAi干涉BmlncR2036表達(圖4: B, C),發現家蠶發育受阻,中腸內容物滯留,最后產生死亡現象,表明BmlncR2036可能具有除調控P25基因外多樣化的功能,其可能參與家蠶中腸的發育調控。本研究從ceRNA和調控啟動子活性兩個角度探索了BmlncR2036參與家蠶發育的可能機制,為研究lncRNA的調控機制提供了實驗依據,但對BmlncR2036詳細的功能探索還需要更多的實驗證據。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
