腦脊液循環動力學機制研究進展
2021-09-02 03:57:58劉俊張國斌
中國神經精神疾病雜志 2021年5期
劉俊 張國斌
經典腦脊液循環理論認為,腦脊液由脈絡叢產生,側腦室及三腦室中的腦脊液經正中孔及外側孔到達小腦延髓池,其與脊髓腔腦脊液混合流入顱底各腦池,最終流向大腦凸面的蛛網膜下腔,由蛛網膜顆粒吸收入靜脈竇完成循環[1]。然而,隨著膠質淋巴循環理論的提出,腦脊液的產生來源、吸收途徑、與腦組織間液的交換以及腦膜淋巴管等新發現,經典腦脊液循環理論受到了挑戰。本文就中樞神經系統腦脊液循環動力學機制的爭議和最新研究進展作一綜述。
1 腦脊液來源與產生方式
1.1 脈絡叢腦脊液由脈絡叢產生是經典腦脊液循環理論核心之一。最早由CUSHING[2]和WEED[3]先后提出,之后在DANDY切除犬的脈絡叢并堵塞室間孔的實驗中得到證實[4]。WELCH[5]發現脈絡膜靜脈的血細胞壓積要顯著高于脈絡膜前動脈,推測脈絡膜血液生成了腦脊液。SPECTOR等[6]詳細闡明脈絡叢如何產生腦脊液的機制,即血漿在靜水壓作用下,由脈絡叢內的毛細血管內皮細胞滲出到內皮間隙,再經過脈絡叢上皮細胞上遍布的鈉鉀泵和水通道蛋白1(aquaporin 1,AQP1)釋放到腦室中形成腦脊液。最新研究發現脈絡叢可在腦代謝廢物增加時或睡眠周期中的特殊時間點代償性增加腦脊液的生成,以優化腦代謝廢物的清除效率[7]。
然而,許多研究并不支持脈絡叢為腦脊液唯一產生部位,例如BERING同樣在犬身上實施DANDY的實驗操作,卻無法觀察到切除脈絡叢側的腦室發生塌陷[8]。MILHORAT發現切除恒河猴的脈絡叢后,其側腦室內腦脊液生成量較未切除脈絡叢的僅輕微減少。嬰幼兒腦積水患者在接受脈絡叢切除術后,仍有三分之二復發而不得不接受分流手術[9]。之后不斷有研究質疑腦脊液只來源于脈絡叢的這一經典理論,人們越發相信腦脊液產生可能存在其他來源。
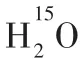
1.3 腦組織間液除了脈絡叢及蛛網膜下腔毛細血管超濾液,腦組織間液也可能是腦脊液來源之一。早在2001年,PéREZ-FíGARES 等[16]即推測,20%腦脊液來源于腦組織間液。RENNELS等[17]發現腦脊液和腦間質液之間交換的最初證據,在犬和貓蛛網膜下腔注入標記物,發現其可沿著血管旁途徑在10 min內進入大腦實質。然而,該研究是基于麻醉這種非生理狀態。
直到2012年,由NEDERGAARD團隊通過雙光子激光掃描顯微鏡首次在活體鼠中發現蛛網膜下腔腦脊液和組織間液之間存在交換,并且是通過AQP4介導,這一機制被稱之為膠質淋巴循環(glympic pathway)[18]。該理論認為,在血管周圍間隙中,腦脊液和組織間液是對流的,組織間液可以通過星形細胞頂端的AQP4快速運輸,在足突端被轉運至血管周圍間隙與腦脊液發生交換,進而隨著腦脊液到達大腦表面被蛛網膜顆粒和靜脈竇吸收,這一機制有力地證明腦組織間液也是腦脊液的重要來源[19-21]。近年來,人們利用核磁示蹤及熒光顯像技術已在活體動物或人體中發現了腦組織間液與腦脊液交換的證據。HAN等[22]利用核磁顯像發現活體大鼠尾狀核處的組織間液可逐漸流向大腦皮層背側和腹側并與其蛛網膜下腔腦脊液匯合。RINGSTAD等[23]在人體腰大池鞘內注射示蹤劑并行強化核磁顯像,發現示蹤劑以一種向心的方式首先在大腦前、中、后動脈的前縱裂區、側裂區、Wills環區富集,且6~9 h后達到高峰,然后示蹤劑逐漸向基底節區、丘腦、中腦等腦深部區域轉移,24 h后基本消失。最近SWEENEY等[20]利用熒光成像技術在活體動物上發現了腦脊液與腦組織間液交換的證據,另外,TAOKA及MESTRE等[24-25]在腦外傷、腦小血管病、梗塞性腦水腫及阿爾茲海默病等中樞神經系統疾病中也發現了腦脊液和腦組織間液交換的證據,只不過這種交換效率會顯著受損。由此可見,膠質淋巴循環理論大大豐富了傳統的腦脊液循環認知,但腦脊液和腦組織間液如何深度地交換融合從而參與腦的代謝和穩態仍舊需要進一步探索。
2 腦脊液循環的流體動力學模式
2.1 從單向流動到雙向流動經典的腦脊液循環模式指出腦脊液循環是沿著腦室-蛛網膜下腔-靜脈竇靜水壓力梯度的單向流動。但是,ORE?KOVIC等[26]認為如果腦脊液循環是單向循環,那么處于平臥位時,由于脈絡叢和靜脈竇的壓力梯度消失,腦脊液循環可能就停滯了,這顯然是矛盾的。另外,一些臨床研究也質疑了脈絡叢是腦脊液生物泵的理論。TAKIZAWA等[27]利用四維相位對比和時空標記反轉脈沖技術對11名健康志愿者側腦室三角區脈絡叢附近的腦脊液活動進行核磁掃描,發現此區域腦脊液流速顯著低于腦干腹側、三腦室和四腦室腦脊液活動,既然脈絡叢推動著腦脊液向前流動,那么此處的腦脊液流速不應該是受到抑制的,這同樣與脈絡叢是腦脊液生物泵的理論相悖。
BULAT-ORE?KOVIC-KLARICA理論認為,不存在脈絡叢的生物泵作用,腦脊液循環是雙向的,即隨著心臟的收縮舒張來回位移,可以從腦室到蛛網膜下腔,亦可以相反從蛛網膜下腔到腦室,腦脊液和腦組織液中任何物質的擴布距離取決于物質的半衰期,半衰期越長,最終將分布到更遠的距離。早在1992年NILSSON等[28]即在核磁研究中觀察到腦脊液循環是雙向的,即導水管的腦脊液發生搏動性流動,流速隨心動周期而變化,在心臟收縮期,腦脊液以大約8 mm/s速度流向后腦(尾端),然后在心臟舒張期間以5 mm/s速度緩慢地回到前腦(頭顱)。因此腦脊液的凈方向是朝向后腦。國內學者[29]同樣驗證了NILSSON發現,他們利用相位核磁技術觀察了凈中腦導水管每搏量,正常人多為頭尾向,而常壓性腦積水患者收縮期和舒張期則均為尾頭向,且呈高動力狀態。該理論可以較好的解釋體位的變化腦脊液循環不會受到明顯的影響,但是雙向循環終歸是一種理想的流動模式。因為雙向循環理論不能很好解釋腦實質內腦脊液與腦組織液的交換,也無法解釋腦代謝廢物如何排出。
2.2 腦脊液和腦組織間液通過膠質淋巴循環深度融合眾所周知,流體的運動既有分子擴散,也有整體流,腦脊液亦是如此。膠質淋巴循環理論認為,蛛網膜下腔的腦脊液與腦組織間液在毛細血管的周圍間隙即Virchow Robin空間(VRS)實現深度交換融合,但基于這種交換的流體動力學諸如驅動力和對流方向等研究目前仍存在爭議。EIDE等[30]在常壓腦積水患者重復了先前RINGSTAD的核磁實驗,他發現腦積水患者較正常人向心性的富集效應顯著減弱,認為這種有規律的富集很難用分子擴散解釋,腦脊液循環可能是在膠質淋巴循環的作用下,以一種整體流的方式運行。
由于分辨率的限制,目前很難利用常規核磁成像觀察VRS中腦脊液與腦組織液交換形式。最近DAVERSINCATTY等[31]利用計算機模擬成像技術發現VRS中腦脊液與腦組織液交換是一種振蕩流和對流的綜合,由腦動脈壁的擴張、剛性運動以及靜態腦脊液壓力梯度共同控制。同樣KOUNDAL等[32]利用動態對比增強核磁顯像技術及數學模型在大鼠中發現,溶質在顱底的大血管旁循環比在中腦和海馬迅速的多,推測蛛網膜下腔和腦池中的腦脊液循環以對流占主導,而在腦實質中,組織液與腦脊液的交換以分子擴散為主,這就能夠解釋腦積水時由于腦室壓升高會導致室管膜通透性增高,腦脊液到組織液的流量增加導致腦白質間質水腫,但是由于蛛網膜下腔腦脊液和組織液的引流效率更高,腦積水卻不會導致大腦皮層灰質水腫。FAGHIH等[33]發現腦脊液-腦組織液的循環不僅通過簡單的分子擴散,而且通過壓力梯度和旋轉重復加速和減速導致非常復雜的運動。由此可見,腦脊液運動不再認為是簡單的循環流動,而是腦室和蛛網膜下腔中各種方向的流動組合,從而適應日常生命活動等復雜的生理過程。
3 腦脊液-組織間液的吸收
3.1 腦膜/脊膜淋巴管腦脊液的產生和吸收是動態平衡的,經典腦脊液循環模式中,腦脊液由硬腦膜上蛛網膜顆粒吸收入靜脈竇。近些年隨著腦膜淋巴管的發現,證實了相當比例的腦脊液是通過腦膜淋巴管引流吸收的。
早在18世紀,PAOLO MASCAGNI首先提到了腦膜淋巴管的存在。由于過于超前,該腦膜淋巴管的存在一直受到質疑。20世紀60年代,F?LDI等[34]重新描述了中樞神經系統與周圍神經系統之間存在淋巴連接。LI等[35]首次在電鏡下觀察到硬腦膜上存在類圓形孔樣的淋巴管,稱為“硬腦膜淋巴孔”,推測該孔可能為蛛網膜下腔腦脊液與顱外結締組織及淋巴系統之間溝通的潛在直截通路。
腦膜淋巴管主要沿著硬腦膜靜脈竇分布。ABSINTA等[36]對人類和非靈長類動物進行硬腦膜淋巴管核磁強化成像,利用兩種不同的增強劑對比呈現了腦膜淋巴管的地形圖,證實了硬腦膜淋巴管的存在,且發現其主要分布在矢狀竇旁附近。NAGANAWA等[37]利用特殊三維實時反轉恢復核磁成像評估內耳淋巴積水時,意外發現在大腦皮質靜脈壁與軟腦膜鞘之間間隙同樣強化,且似乎沿著上矢狀竇與硬腦膜淋巴管相連,推測這一間隙可能是腦脊液吸收入硬腦膜淋巴管的途徑。RINGSTAD等[38]研究同樣支持這種觀點,18例不同腦脊液循環紊亂疾病患者進行鞘內注射腦脊液示蹤劑后行核磁強化,發現對比劑集中在皮質靜脈匯入矢狀竇的竇旁硬腦膜附近,他認為的確存在類似于跨蛛網膜的分子通道,其位于矢狀竇旁的硬腦膜,可能是溝通腦脊液和硬腦膜淋巴管的橋梁。
腦膜淋巴管可通過嗅神經淋巴途徑參與腦脊液吸收。BRADBURY等[39]發現蛛網膜下腔內腦脊液可以穿過篩板沿著嗅神經進入鼻粘膜的淋巴管,最后匯入頸部淋巴結。ASPELUND等[40]和 LOUVEAU等[41]在嚙齒類、魚類、靈長類以及人類發現,腦脊液成分的分子片段會引流到腦膜淋巴管并沿著肩胛后靜脈和乙狀竇以及翼腭動脈的腦膜部分匯入到頸深淋巴結。如前所述,RINGSTAD等[38]研究同樣發現18例患者有2例篩板附近出現了對比劑增強,推測篩板附近的嗅神經途徑參與了腦脊液吸收。
除了硬腦膜及嗅神經,脊神經附近蛛網膜顆粒和硬脊膜淋巴管也可參與腦脊液的吸收。KIDO等[42]及MIURA等[43]發現了脊神經根處存在蛛網膜顆粒和淋巴管。BOZANOVICSOSIC等[44]發現貓有16%的腦脊液通過脊髓周圍流出道排出。KOH等[45]發現脊髓蛛網膜下腔的腦脊液可以沿著脊神經匯入肋間淋巴結。
3.2 蛛網膜顆粒盡管蛛網膜顆粒是腦脊液吸收的重要部位,但最新研究表明,蛛網膜顆粒及靜脈竇可能并非恒定參與腦脊液循環。AKAI等[46]在活體胎鼠上發現,不論在腦室還是在蛛網膜下腔注射熒光劑,均未發現矢狀竇和蛛網膜顆粒發現熒光信號。EIDE在核磁上觀察到示蹤劑是隨著腦脊液向心性的轉移直至消失[30],示蹤劑并不在硬腦膜及靜脈竇旁富集,這進一步質疑了蛛網膜顆粒的吸收功能。ZAKHAROV等[47]研究與這種推論相吻合,他在綿羊小腦延髓池注射放射性示蹤劑后發現,當顱內壓正常時,腦脊液流出的主要途徑是淋巴管,而只有顱內壓升高時,才會觸發向蛛網膜顆粒及顱內靜脈系統的轉運。MURTHA等[48]也在大鼠上發現,腦室注射的造影劑并沒有出現在靜脈竇中,而是大部分很快穿過篩板進入嗅神經周圍鼻粘膜。
除了蛛網膜顆粒和靜脈竇,MALOVESKA等[49]發現腦脊液的引流吸收途徑可能部分是通過膠質淋巴循環進入腦膜淋巴管或者穿過篩板到嗅神經周圍淋巴管或者直接被腦膜淋巴管吸收完成。當然,血管旁路的膠質淋巴循環和腦膜淋巴循環是否存在協同作用目前仍不清楚。關于在不同的生理和病理條件下腦脊液吸收如何有所側重地發揮作用,目前仍缺少研究。另外,MA等[50]報道,寰枕筋膜處枕下小直肌與枕部硬腦膜之間的肌硬膜橋可能參與調節局部腦脊液循環,當刺激犬枕下肌肉收縮或者被動轉頭時局部腦脊液壓力增加。最近發現對美洲短吻鱷枕下肌硬膜橋實施電刺激收縮也出現了局部腦脊液流速增加的現象[51]。
因此,腦脊液循環動力學異常復雜,其來源、循環模式以及吸收部位可能并非一成不變,而是由眾多因素調控。不同的日常生理性活動及病理性疾病狀態下腦脊液循環動力學可能會發生不同變化,后期仍需要深入研究。
