絹蒿荒漠糞甲蟲多樣性特征及其對糞便內種子的二次分配
2021-08-21 02:32:22曹佳敏郭亞亞李娜娜孫海榮車昭碧魯為華
草業學報 2021年8期
關鍵詞:物種
曹佳敏,郭亞亞,李娜娜,孫海榮,車昭碧,魯為華,2*
(1.石河子大學動物科技學院,新疆 石河子832003;2.省部共建綿羊遺傳改良與健康養殖國家重點實驗室,新疆 石河子832003)
牲畜糞便是天然草地放牧生態系統中重要的養分來源之一[1]。放牧家畜通過消化只能吸收利用所采食牧草營養成分的一部分,其余60%~90%的養分以糞便和尿液的方式返還到草原生態系統[2],糞便在草地上的堆積能夠有效提高糞堆下土壤水分、有機質、氮、磷和鉀的含量,降低土壤pH值,利于周圍植物生長[3],但是草食動物新鮮糞便質地黏稠,如無食糞昆蟲參與分解,糞便破碎化和分解速度緩慢,會造成糞便下植物缺乏氧氣和光照而死亡,同時,糞便堆積導致糞便下植物生長受阻,甚至會波及糞堆外圍3~5 cm處,造成糞便下及其周邊范圍內的草地生產力降低,并且導致牲畜拒絕取食糞便周圍的牧草[4-5],據統計,家畜排泄物占據放牧地30%~40%的面積[6],如果糞便不能迅速分解,會損失大片的放牧區域,降低草地放牧利用率,因此,排泄到草地上的糞便必須及時分解才能實現草地的有效保護利用,同時實現養分的正常循環。
糞甲蟲作為食糞動物最大的類群,主要包括金龜科(Scarabaeidae)、蜉金龜科(Aphodiidae)和糞金龜科(Geotrupidae),同時,根據糞便處理方式將糞甲蟲分為4個功能類群:a)遠生型糞甲蟲(roller)將糞便團成球狀,滾到一定距離后掘洞并把糞球埋到洞中,作為食物球或孵化球;b)外生型糞甲蟲(tunneler)直接在糞便下掘洞,將糞便搬運到洞中作為食物球,或產卵于其中,作為孵化球,成為后代繁殖場所和食物來源;c)內生型糞甲蟲(dweller)直接在糞中取食,一旦找到糞便便一頭扎進糞便中,其整個生活史中的卵、幼蟲和蛹發育都發生在糞便中[7];d)偷竊寄生型糞甲蟲(kleptoparasites)使用其他糞甲蟲的資源,寄生于糞球中。糞甲蟲通過取食和搬運改變糞便內部形狀,使其內部穿有小孔,加速糞便內部空氣流通,從而有利于微生物的取食和糞便的自然分解,微生物對糞便分解主要是對液體部分進行采食和養分的釋放,此外,大型食草動物對糞便的踩踏有利于糞便破碎使得碎屑進入土壤,進而加速糞便分解,糞便經過分解進入生態系統,從而參與生態系統的物質循環和能量流動[8-10]。盡管不同功能類群的糞甲蟲對糞便處理方式不一樣,但最終能夠快速改變糞便的形狀,使其破碎化,加快糞便的分解速度[11]。
草食動物在采食植物營養體的同時,會將成熟的種子一同吞咽并通過消化道后隨糞便排放到草地上,從而實現種子的消化道傳播(endozoochorous)。糞甲蟲是絹蒿荒漠家畜糞便的主要分解者,在扮演分解者這一角色的同時,它們又會對糞便內的種子進行二次傳播。因此,糞甲蟲會決定糞便內種子的命運和去向。由于糞甲蟲有多個種類和生態功能類型,并且其種類和數量在時間和空間上存在特定的變化規律,這種變化規律又會對糞便的分解及糞便內種子的二次傳播產生影響,在人為建立起來的絹蒿荒漠草地放牧系統中,由于有嚴格的季節選擇性[12],使草食家畜密集性地集中在秋季放牧采食、糞便排泄,這種高強度、密集性的放牧過程顯然也會影響草地植物種子的傳播[13],此外,在組成相對簡單的荒漠生態系統中,不僅糞便的分解過程強烈依賴于糞甲蟲,而且糞便內種子的再分配也由于缺乏其他有效的動物傳播媒介而對糞甲蟲有強烈的依賴性。本研究主要通過不同放牧季節(主要是春秋兩季)的野外調查和模擬控制試驗,以期闡明:1)不同季節絹蒿荒漠的糞甲蟲的種類和數量特征及其數量變化規律是什么?2)各類糞甲蟲對糞便的進駐和取食特征如何?3)通過模擬試驗,探討種子在糞便內被糞甲蟲二次分配的特征。
1 材料與方法
1.1 研究區概況
研究地點選取石河子紫泥泉種羊場的典型絹蒿荒漠放牧地(一大灘)(E 85°49'-85°58',N 43°26'-45°20'),分布海拔980~1120 m。該牧場為重要的春秋牧場,放牧時間包括4-6月(春季)、9-11月(秋季)兩個時間段。屬于典型溫帶荒漠干旱氣候,年平均溫度8℃,月平均最高溫度集中在7月,為26.4℃。年降水集中在4、5、8月,年平均降水量180~200 mm。土壤為淡栗鈣土。植被組成以伊犁絹蒿(Seriphidium transiliense)為建群種,伴生種有草原苔草(Carex liparocarpos)、羊茅(Festuca ovina)、新疆針茅(Stipa sareptana)、駱駝蓬(Peganum harmala)以及一年生草本角果藜(Ceratocarpusarenarius)和豬毛菜(Salsola collina),植被結構組成較為復雜。
1.2 試驗設計及樣品采集
1.2.1 糞甲蟲季節性群落組成和多樣性分析 研究時間為2019年4-10月,研究區為典型的絹蒿荒漠放牧地,為了盡可能覆蓋試驗區域,保證在絹蒿荒漠中對物種多樣性調查的準確性。利用GPS全球定位系統,在空間上將放牧地按海拔每升高20 m為一個調查梯度設置8個不同的海拔。分別為980、1000、1020、1040、1060、1080、1100和1120 m。在4-9月中每個月連續5 d對放牧區域進行調查。調查時首先在放牧區域內的每一調查梯度上設置垂直于調查梯度的50 m×50 m的3個重復樣地,在每個樣地內設置10個5 m×5 m的樣方,對每個樣方內馬、牛的糞便類型、單位面積重量進行計算、統計,然后分別對不同家畜糞便內及糞便下10 cm深土壤內所存在的糞甲蟲種類進行手撿、記數,制作標本帶回實驗室,通過《中國北方常見金龜子彩色圖鑒》[14]和李久文[15]的方法進行分類鑒定。
采用計數法對糞便中糞甲蟲的數量進行確定,將糞甲蟲裝入不密封的管中饑餓12 h排除體內殘渣,在水浴鍋中加熱,待蟲子僵直后取出放置于無水乙醇中,置于4℃冰箱中保存,用體式顯微鏡對糞甲蟲進行拍攝并測量記錄體尺數據。用形態學分類法對糞甲蟲進行分類和鑒定。
1.2.2 糞甲蟲進駐糞便行為觀察 糞甲蟲的行為學研究在典型絹蒿荒漠放牧地進行,時間為6、10月。在清晨收集牛的新鮮糞便,糞便收集后進行充分攪拌混勻。每份糞便為500 g,設置在直徑為30 cm的圓形區域,間隔2 m各6份,共需等量糞便12份。在9:00將糞便放置在指定區域,同時開始用攝影機進行連續拍攝觀察。從9:00開始,每隔15 min對環境條件進行記錄,包括溫度、濕度、風速、光照強度。試驗開始后,在2、6、12、24、48 h時采用全收糞法收集前5份糞便,直至糞便內再無任何糞甲蟲個體存在,將最后一份糞便小心回收裝袋。
通過觀察,明確糞甲蟲的到達時間、到達順序以及不同的糞甲蟲是如何處理糞便的。結合不同時間段對糞甲蟲的收集,確定糞便中糞甲蟲的數量高峰。
1.2.3 糞甲蟲進駐對糞便內種子的分配 用圓形塑料珠作種子模擬物[16]。種子模擬物比真實種子更可取,這樣可以防止糞便中的種子與自然存在的種子相混淆。使用了3種不同直徑的珠子:小(1.3~1.9 mm;每重復n=100),中(3.3~3.7 mm;n=40)和大(4.8~5.8 mm;n=5)。試驗當天,在研究區放置500 g新鮮糞便,其中帶有嵌入的種子模擬物。糞便沉積至無糞甲蟲活動采集土壤樣品。在每個采樣點,在以糞便為圓心半徑為20 cm的圓形區域內以0~1 cm、2~5 cm、5~10 cm深度對土壤層進行采樣。通過篩分土壤樣品對珠子進行計數,以計算在不同深度被糞甲蟲掩埋的珠子的比例。本研究認為未收回的珠子是被糞甲蟲傳播垂直距離大于10 cm或水平距離大于20 cm。
1.3 數據處理
利用Excel軟件計算多樣性指標Margalef豐富度指數(d Ma)、Simpson指數(D)、Shannon-Wiener指數(H')、Pielou均勻度指數(J)。
2 結果與分析
2.1 糞甲蟲季節性群落組成和多樣性分析
在8個海拔梯度覆蓋整個研究區采集到食糞昆蟲14797頭。其中捕獲糞甲蟲分屬3科17種,蜉金龜科9種:黃緣蜉 金龜(Aphodius sublsmbatus)、黑蜉金龜(A.breviassulus)、血斑蜉金龜(A.haemorrhoidalis)、紅亮蜉金龜(A.impunctatus)、黑格(三斑)蜉金龜(Aphodiidae nigrotessellatus)、直蜉金龜(A.rectus)、蜉金龜科一種(Aphodiidae sp.1)蜉金龜科一種(Aphodiidae sp.2)、蜉金龜科一種(Aphodiidae sp.3);金龜科7種:小駝嗡蜣螂(Onthophagusgibbulus)、獨行嗡蜣螂(O.solivagus)、三開蜣螂(Copristripartitus)、北方 蜣 螂(C.lunaris)、粗 紋 彎 邊 蜣 螂(Gymnopleurus flayellates)、金龜科一種(Scarabaeidae sp.1)、金龜科一種(Scarabaeidae sp.2);糞 金 龜 科1種 :糞 金 龜(Geotrupeslaeristriatus)。
研究區4月牛糞生境中的優勢種為金龜科的三開蜣螂,個體數占牛糞生境的34.09%。5月優勢種為蜉金龜科的黃緣蜉金龜,個體數占牛糞生境的22.21%。6月優勢種為蜉金龜科的紅亮蜉金龜,個體數占牛糞生境的21.67%。7月優勢種為金龜科的北方蜣螂,個體數占牛糞生境的30.42%。8-10月優勢種均為蜉金龜科的直蜉金龜,個體數分別占各自生境49.90%、93.55%、99.6%(圖1)。
6月牛糞中捕獲的食糞昆蟲個體數在7個月中僅排第5位,但擁有最多的物種和豐富度(圖2),Simp?son指數、Shannon-Wiener指數、均勻度指數也高于其他月份(表1)。8月牛糞中捕獲食糞昆蟲個體數最少,但各項指數不是最低的。10月牛糞中捕獲的食糞昆蟲個體數量最多(圖1),但各項指數均最低。不同月份物種的豐富度與物種的數量呈正相關,物種的數量越多其測度值也越高。各月牛糞中食糞昆蟲的物種數與各地的豐富度是相一致的:6月>5月>7月>4月>8月>9月>10月(表1)。在物種總數相同的條件下,多樣性指數與各物種個體數量的平均程度呈正相關。物種的均勻度指數則受各物種個體數量分布的均勻性影響。因此,盡管10月有最大的個體數量,但因各物種的個體數量分布極不均勻,即直蜉金龜占了捕獲個體數的絕大部分,致使其多樣性和均勻度指數均最低。
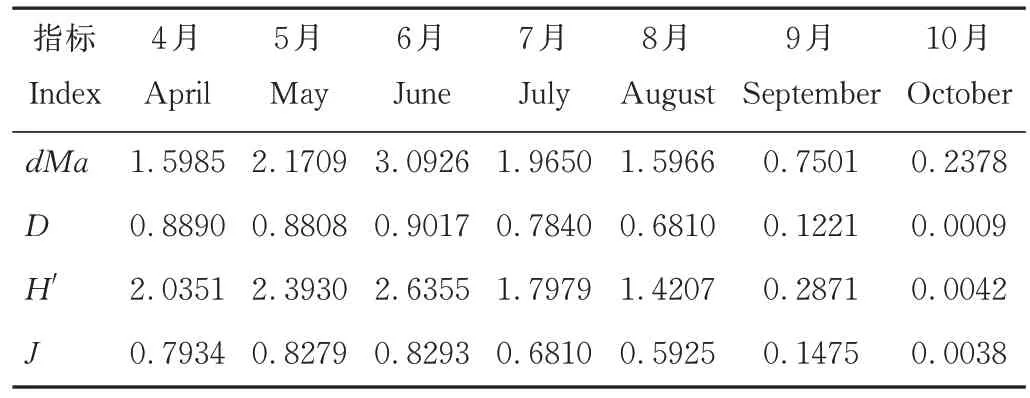
表1 不同月份牛糞中食糞昆蟲的多樣性Table 1 The diversity of insects in cattle dung in different months
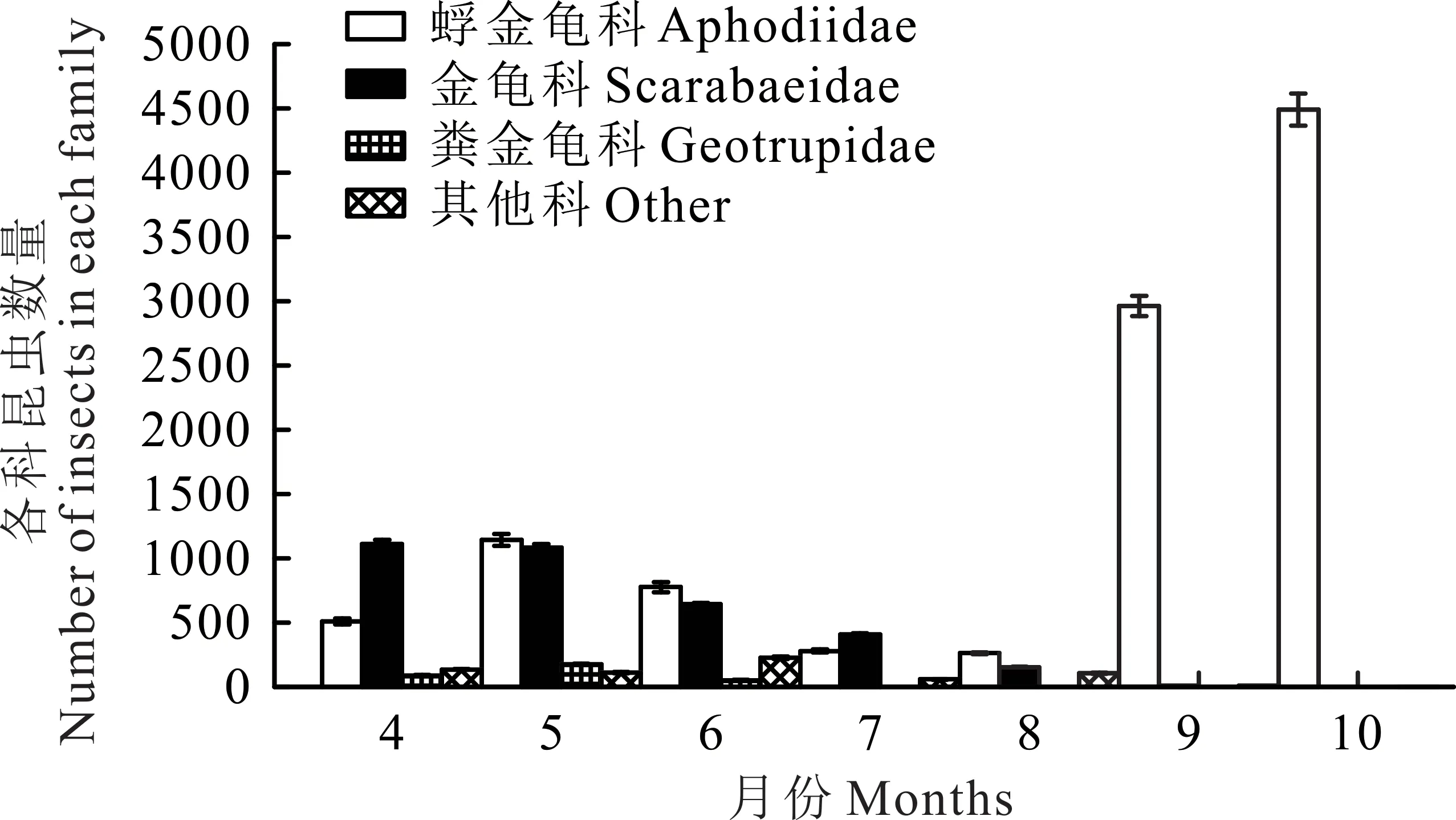
圖1 各科昆蟲不同月份數量變化Fig.1 Insects of different families in different months
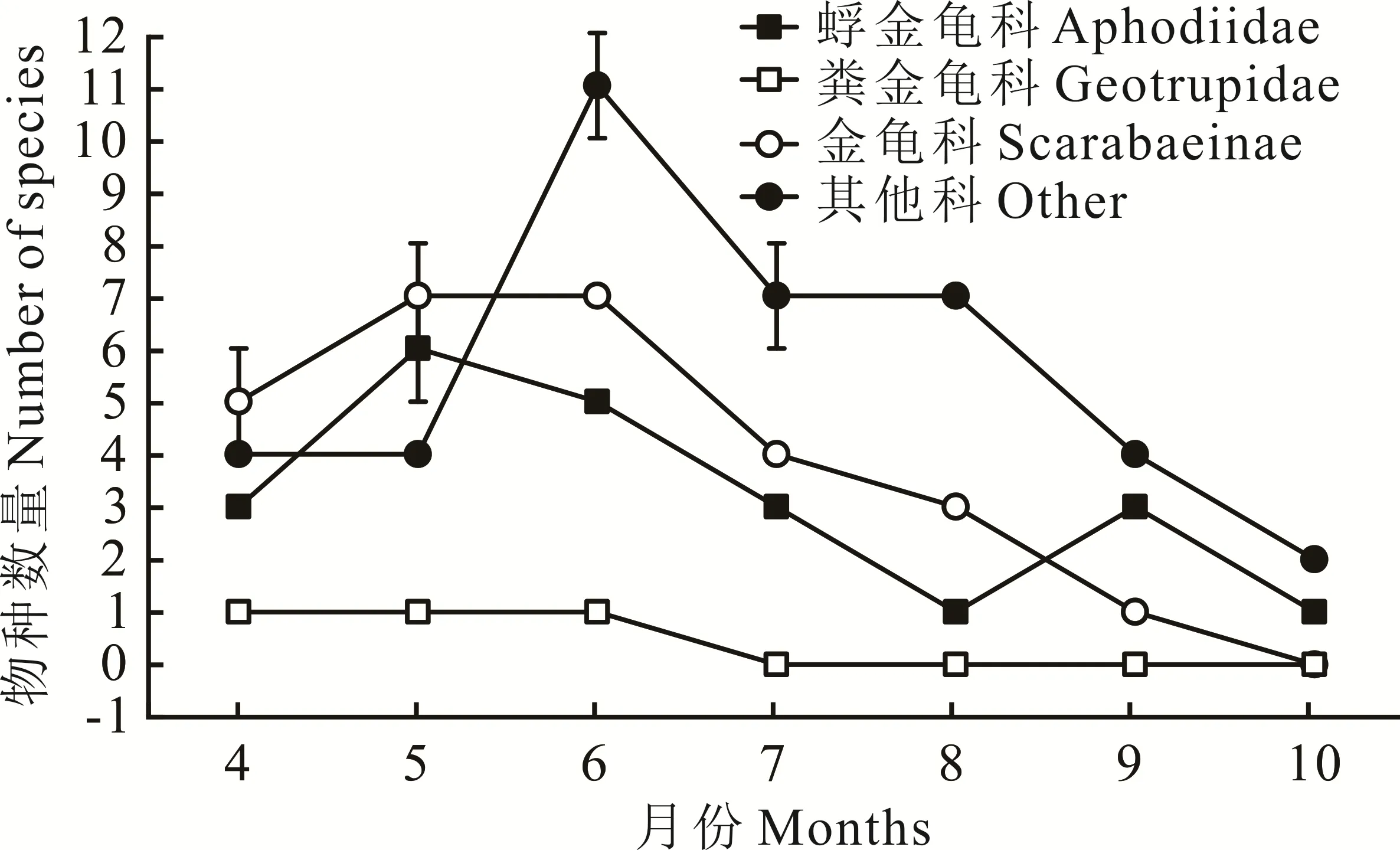
圖2 各科昆蟲不同月份種類動態變化Fig.2 The dynamic change of insects of differ ent families in different months
2.2 糞甲蟲進駐糞便行為觀察
行為觀察試驗在6月糞甲蟲物種數最多和10月糞甲蟲個體數最大的兩個月進行。主要觀察糞甲蟲拜訪糞便的方式,不同種類的糞甲蟲到達糞便的先后順序。不同時間段糞便中累計活動的糞甲蟲數量以及被糞甲蟲分解的糞便狀態。6月糞甲蟲種類多數量少,在糞便中活動時間主要集中在新鮮糞便放置的前24 h,數量高峰在糞便放置8 h左右(累計100+)。優勢種為內生型糞甲蟲紅亮蜉金龜,其次是遠生型糞甲蟲粗紋彎邊蜣螂。10月糞甲蟲種類少數量多,在糞便中活動時間主要集中在新鮮糞便放置的前8 h,數量高峰在糞便放置4 h左右(累計2800+),糞便放置23 h糞甲蟲完全結束在糞便中的活動。優勢種為內生型糞甲蟲直蜉金龜(表2)。

表2 糞甲蟲進駐糞便行為觀察Table 2 Observation on the behavior of dung beetle in d
6月的溫度與糞甲蟲數量沒有顯著相關性(表3);濕度與糞甲蟲數量沒有顯著的相關性;風速與糞甲蟲數量呈顯著負相關(P<0.05);光照強度與糞甲蟲數量呈顯著正相關(P<0.05);10月溫度與糞甲蟲數量呈顯著正相關(P<0.05);濕度與糞甲蟲數量呈極顯著負相關(P<0.01);風速與糞甲蟲數量沒有顯著的相關性;光照強度與糞甲蟲數量呈極顯著正相關(P<0.01)。
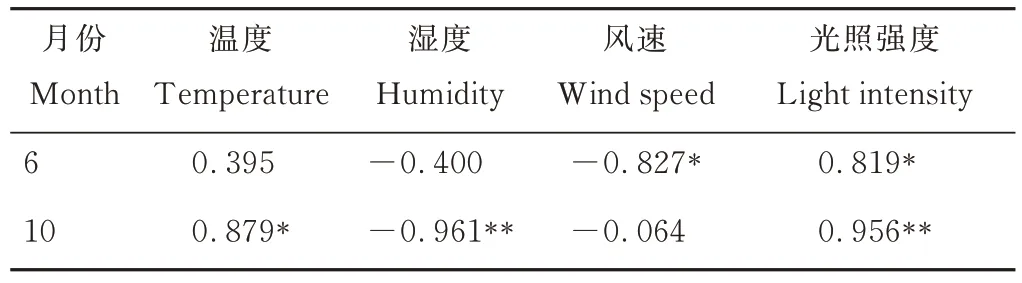
表3 糞甲蟲數量與環境因子之間的皮爾森相關系數Table 3 Pearson’s correlation coefficient between the number of dung beetles and environmental factors
2.3 糞甲蟲進駐對糞便內種子的分配
80%的珠子被埋藏在水平距離20 cm以內及范圍0~10 cm的土壤深度中(圖3),主要處理珠子的糞甲蟲為內生型糞甲蟲、外生型糞甲蟲和遠生型糞甲蟲。有20%的珠子被糞甲蟲埋藏到10 cm以下或移動到水平范圍20 cm以外,主要處理珠子的糞甲蟲為遠生型糞甲蟲。3種尺寸的珠子在0~1 cm、2~5 cm、5~10 cm的埋藏深度中均表現出顯著性差異(P<0.05)。在0~1 cm的埋藏深度中大、中、小3種尺寸的珠子埋藏率呈逐漸降低的趨勢,且差異顯著(P<0.05)。在2~5 cm的埋藏深度中小珠子的埋藏率最高,與大、中珠子差異顯著(P<0.05)。在5~10 cm埋藏深度中,大珠子的埋藏率為0,與中、小珠子差異顯著(P<0.05)。3種尺寸的珠子在>10 cm的埋藏深度中差異不顯著(P>0.05)。
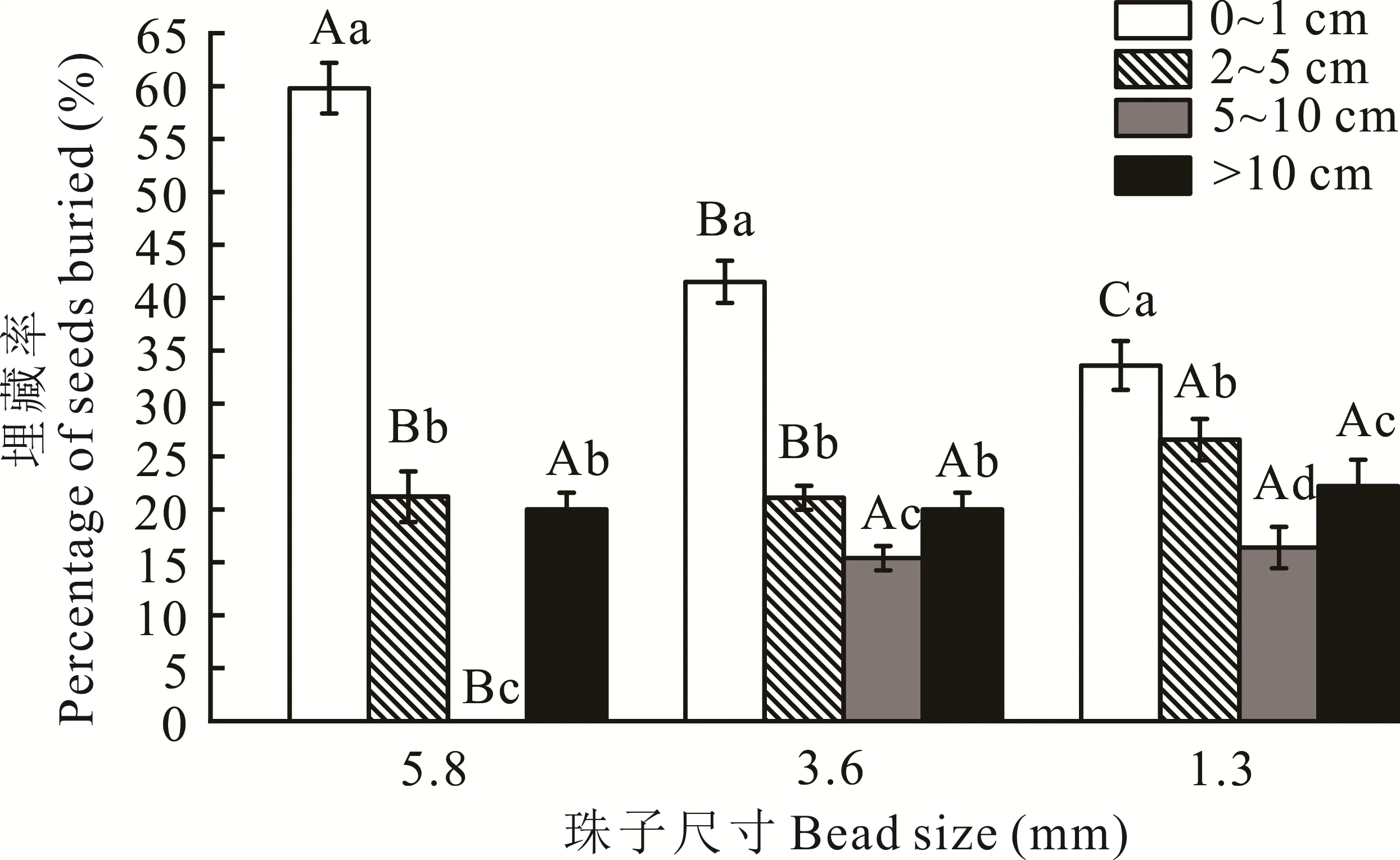
圖3 糞甲蟲對不同尺寸珠子的埋藏率Fig.3 Burial rate of dung beetle to different bead sizes
3 討論
3.1 糞甲蟲的多樣性特征
糞甲蟲多樣性的研究主要集中在熱帶雨林和非洲稀樹草原中,研究發現草原上糞甲蟲的數量是森林的8倍,草原上遠生型糞甲蟲白天主導,外生型糞甲蟲晚上主導。森林里外生型糞甲蟲在白天和晚上都是主導,物種豐富度很低[17]。在印度的牧區研究中,內生型糞甲蟲在春天至秋天活動頻繁,遠生型糞甲蟲在春至夏季以及冬天之前活動頻繁[18]。溫帶草原的研究發現低緯度地區遠生型糞甲蟲和外生型糞甲蟲占主導地位,北部溫帶地區內生型糞甲蟲占主導地位并有少量的外生型糞甲蟲伴隨,本研究結果與此一致。干燥地區的遠生型糞甲蟲數量比潮濕地區的多,可能是因為潮濕地區的土質濕黏不易滾動以及樹葉凋落物過厚阻礙其活動[19]。
本試驗在取樣過程中除糞甲蟲外還在糞便中采集到不少食糞性昆蟲,在數據統計結果中會直接影響到牛糞中物種豐富度,但由于采集到的糞甲蟲數量龐大,所以對其他指數影響較小。作為荒漠草地生態系統的主要成員,糞甲蟲對生態系統的平衡有重要意義。通過本試驗分析認為導致不同月份牛糞中糞甲蟲群落多樣性差異的主要因素有2個。一是氣候因素決定了糞甲蟲群落的主要組成,溫度、濕度、風速和光照強度都對糞甲蟲有影響。二是季節性放牧影響糞甲蟲的種類和數量,研究地作為春秋牧場,承擔了兩季放牧壓力。糞甲蟲依靠嗅覺尋找糞便[20],糞便的數量和密度會直接影響糞甲蟲種類和數量。
3.2 糞甲蟲拜訪行為的日變化及其與環境因子之間的關系
影響6和10月糞甲蟲拜訪行為日變化不同的主要原因是糞甲蟲的種類和數量,6月的牛糞中活動的優勢種為內生型糞甲蟲紅亮蜉金龜,其次是遠生型糞甲蟲粗紋彎邊蜣螂。10月的優勢種為內生型糞甲蟲直蜉金龜。不同功能類群的糞甲蟲對糞便處理方式的差異導致利用時長不一致。6月糞甲蟲數量少競爭小,因而能在糞便中獲得足夠的食物再離開。但10月個體數量大糞便是稀缺資源,當糞便中的養分不能維持取食或產卵育幼時糞甲蟲也會自行離開。其次環境因子也對糞甲蟲的活動有重要影響,隨著氣溫的升高或風速過大糞便失水干燥,開始結塊,糞甲蟲很難利用此類糞便也會離開。糞甲蟲數量與環境因子之間的皮爾森相關系數有明確的季節性差異。6月物種豐富度高,溫度和濕度對糞甲蟲數量沒有表現出顯著相關性。10月的糞甲蟲物種較為單一且數量大,環境因子對其影響較純粹,糞甲蟲表現出喜熱喜干的特性。這與熱帶雨林研究的糞甲蟲喜熱喜濕情況不符,可能的原因是環境和氣候的差異造成不同地區不同物種的聚集,而絹蒿荒漠炎熱干燥,物種不斷適應當地氣候所致。
3.3 糞甲蟲對種子的搬運和埋藏特征
隨著種子大小的增加,埋入種子的比例降低[21]。模擬試驗結束,在土壤表面僅可見少量珠子。不同功能類群的糞甲蟲對糞便處理方式的差異導致不同尺寸珠子的埋藏率不同。本試驗認為0~1 cm層的珠子大部分是由內生型糞甲蟲加工的[22],3種尺寸的珠子在此深度埋藏率最高,與6月的優質種內生型糞甲蟲紅亮蜉金龜相對應。2~10 cm的珠子則由糞金龜加工,多數糞金龜會將珠子埋在2~5 cm糞便下方挖掘的洞中,少數糞金龜會因競爭將珠子深埋。遺失的珠子或被掩埋深度大于10 cm被認為是遠生型糞甲蟲粗紋彎邊蜣螂移出了試驗區。粗紋彎邊蜣螂將糞便團成球狀,滾到一定距離后掘洞并把糞球埋到洞中,作為食物球或孵化球,其中珠子會摻雜在糞球中被二次傳播。
在主要通過哺乳動物排便傳播種子的情況下,糞甲蟲雖然不是專性主動的種子傳播者,但通過利用哺乳動物糞便在多階段種子擴散過程中起關鍵作用。糞甲蟲對于微環境主動而嚴格的選擇性使得糞便內種子的存活率大大增加,具有傳播方式上的優越性。糞甲蟲在將其用于喂養或產卵之前將其掩埋。在此過程中,嵌入糞便中的一粒或幾粒種子可能會意外地埋在糞便下的隧道中或遠離糞便的糞球中[23]。從糞甲蟲的角度來看,糞便中存在的種子是污染物[24]。從種子的角度來看,離開母株和埋葬可能會極大地影響其生存[25-26]。埋在地下的種子比地面上殘留的種子更有可能逃脫捕食[21]。此外,種子最終可能處于潮濕的環境中,更適合發芽。糞甲蟲對種子的埋藏有利于建立土壤種子庫,種子在土中休眠,隨后成功發芽[27]。從植物的角度來看,深度很重要,因為當種子被更深地埋藏時,它們更容易被掠食者發現[28]。然而,糞甲蟲深埋種子也會抑制出苗[29],即使深度小于5 cm的微小增加也可以顯著降低出苗和成功建植的可能性[30]。
4 結論
1)絹蒿荒漠糞甲蟲的物種多樣性特征存在季節性差異,春季糞甲蟲物種豐富度高但個體數量少。秋季糞甲蟲物種豐富度低但個體數量大。
2)6月的糞甲蟲對糞便的拜訪行為數量高峰在8 h,累計100+,24 h后數量開始減少,活動持續48 h完全結束。10月的糞甲蟲對糞便的拜訪行為數量高峰在4 h,累計2800+,6 h后數量開始減少,活動持續23 h完全結束;糞甲蟲的拜訪數量與環境因子之間存在顯著相關性。
3)不同尺寸的珠子之間的埋藏率差異顯著,不同深度之間的埋藏率有顯著性差異。隨著種子大小的增加,埋入種子的比例逐漸降低。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
