不同載畜率對短花針茅荒漠草原優(yōu)勢物種間關(guān)系及其空間分布特征的影響
2021-08-21 02:32:00張峰孫嘉偉孫宇鄭佳華喬薺瑢趙萌莉
草業(yè)學報 2021年8期
關(guān)鍵詞:物種
張峰,孫嘉偉,孫宇,鄭佳華,喬薺瑢,趙萌莉*
(1.內(nèi)蒙古農(nóng)業(yè)大學草原與資源環(huán)境學院,內(nèi)蒙古 呼和浩特010019;2.內(nèi)蒙古農(nóng)業(yè)大學草地資源教育部重點實驗室,內(nèi)蒙古呼和浩特010019)
種間關(guān)系是決定草地群落組成結(jié)構(gòu)、動態(tài)變化的重要因素[1],是形成草地群落空間格局的主要動力[2]。目前,對草地群落物種間關(guān)系的研究較多[3-4],但在反映物種在群落中的相對重要性及物種間親和或競爭的相對關(guān)系時,卻無法直觀反映種群在空間上的動態(tài)變化規(guī)律[5]。借助地統(tǒng)計學對不同草地類型的不同指標測度進行分析[6-8],能反映植物種群空間分布格局的動態(tài)變化,并能模擬還原物種分布及其種間關(guān)系。近年來,由于過度放牧造成各草地類型出現(xiàn)不同程度的退化,使得群落蓋度、多樣性及生產(chǎn)力降低[9];而荒漠草原是我國北方天然的生態(tài)屏障,是以放牧為主的畜牧業(yè)基地[10],其氣候干燥、生境脆弱且對干擾響應(yīng)敏感[11]。但目前對荒漠草原放牧干擾的相關(guān)研究主要集中于群落結(jié)構(gòu)、物種多樣性、生產(chǎn)力、性狀屬性及土壤理化性質(zhì)等方面[12-13],對多物種間空間分布關(guān)系的研究較少;而張峰等[14]對大針茅(Stipa grandis)典型草原空間分布關(guān)系的研究發(fā)現(xiàn),放牧會導致種間關(guān)系改變;而Callaway等[15]和Armas等[16]的研究表明,植物的相互作用影響著種群動態(tài)。
以短花針茅(Stipa breviflora)荒漠草原為對象,結(jié)合常規(guī)分析方法與地統(tǒng)計學方法,分析建群種短花針茅與優(yōu)勢種無芒隱子草(Cleistogenes songorica)和冷蒿(Artimisia frigida)種間關(guān)系及空間分布特征,闡明優(yōu)勢物種的種間關(guān)系及其作用強度,并結(jié)合空間模擬揭示物種間的動態(tài)變化,為草地合理利用提供理論基礎(chǔ)。本研究旨在回答以下3個問題:1)優(yōu)勢種無芒隱子草和冷蒿與建群種短花針茅之間是競爭關(guān)系還是親和關(guān)系?不同載畜率是否會改變物種之間的關(guān)系?2)物種之間的作用強度是怎樣的?不同載畜率是否會改變物種間的作用強度?3)不同載畜率下優(yōu)勢物種間的空間分布特征是怎樣的?對其進行空間模擬是否具有實際意義?
1 材料與方法
1.1 研究區(qū)概況
試驗區(qū)位于內(nèi)蒙古農(nóng)牧業(yè)科學院綜合試驗示范中心四子王旗基地(41°46′44″N,111°53′42″E),海拔1456 m;典型的中溫帶大陸性季風氣候,春季干旱多風,夏季炎熱,而冬季寒冷漫長。>10℃的年積溫為2200~2500℃,氣溫平均日差為13~14℃;平均無霜期約為108 d。在2004-2018年期間,年均溫3.4℃,最冷月在1月,均溫約為-16.03℃,最熱月在7月,均溫約為20.82℃,極值為-20.38和23.73℃;年均降水量為234 mm,降水主要集中在4-9月;年均風速為4~5 m·s-1。地帶性植被類型為短花針茅荒漠草原,建群種為短花針茅,主要優(yōu)勢種為無芒隱子草和冷蒿,伴生種為銀灰旋花(Convolvulus ammanni)、阿爾泰狗娃花(Heteropappus altaicus)、小葉錦雞兒(Caragana microphylla)、木地膚(Kochia prostrata),其他常見植物有糙蘇(Phlomis umbrosa)、駝絨藜(Ceratoideslatens)等,還有一、二年生植物地錦(Euphorbia humifusa)、刺穗藜(Chenopodum aristatum)等;植被稀疏低矮,群落高度為8~10 cm,蓋度為17%~20%。土壤為淡栗鈣土,土層厚度約為1 cm,較貧瘠,有機質(zhì)含量較低,約為14 g·kg-1。
1.2 試驗設(shè)計
試驗區(qū)于2004年進行圍建,設(shè)置對照(control,CK)、輕度放牧(light grazing,LG)、中度放牧(moderate grazing,MG)和重度放牧(heavy grazing,HG)4個載畜率水平,放牧家畜為當?shù)爻赡昝晒鹏删d羊,分別為0、4、8、12只,載畜率分別為0(CK)、0.15(LG)、0.30(MG)和0.45(HG)羊單位·hm-2·月-1;3次重復,共12個小區(qū),各小區(qū)面積均為4.4 hm2;在圍建前,草地放養(yǎng)綿羊且草地利用強度較大,但圍建時各小區(qū)草地及土壤狀況較一致。自2004年起,放牧期于6月初開始并于11月底結(jié)束。
由于12個小區(qū)均呈現(xiàn)南北長、東西窄的特點,本試驗于2020年7-9月在每個小區(qū)分別設(shè)置1個25 m×100 m的大樣方,均等劃分為50個5 m×10 m的中樣方(南北長10 m,東西寬5 m)(圖1),在每個中樣方東南角設(shè)置1個1 m×1 m的小樣方;各小區(qū)小樣方均為50個,共計600個。以西南角作為起點(0,0),自西向東為X軸,自南向北為Y軸,最西南角坐標為(0,0),最東北角坐標為(20,90);記錄各小樣方中短花針茅、無芒隱子草及冷蒿的株叢數(shù)(多度)。
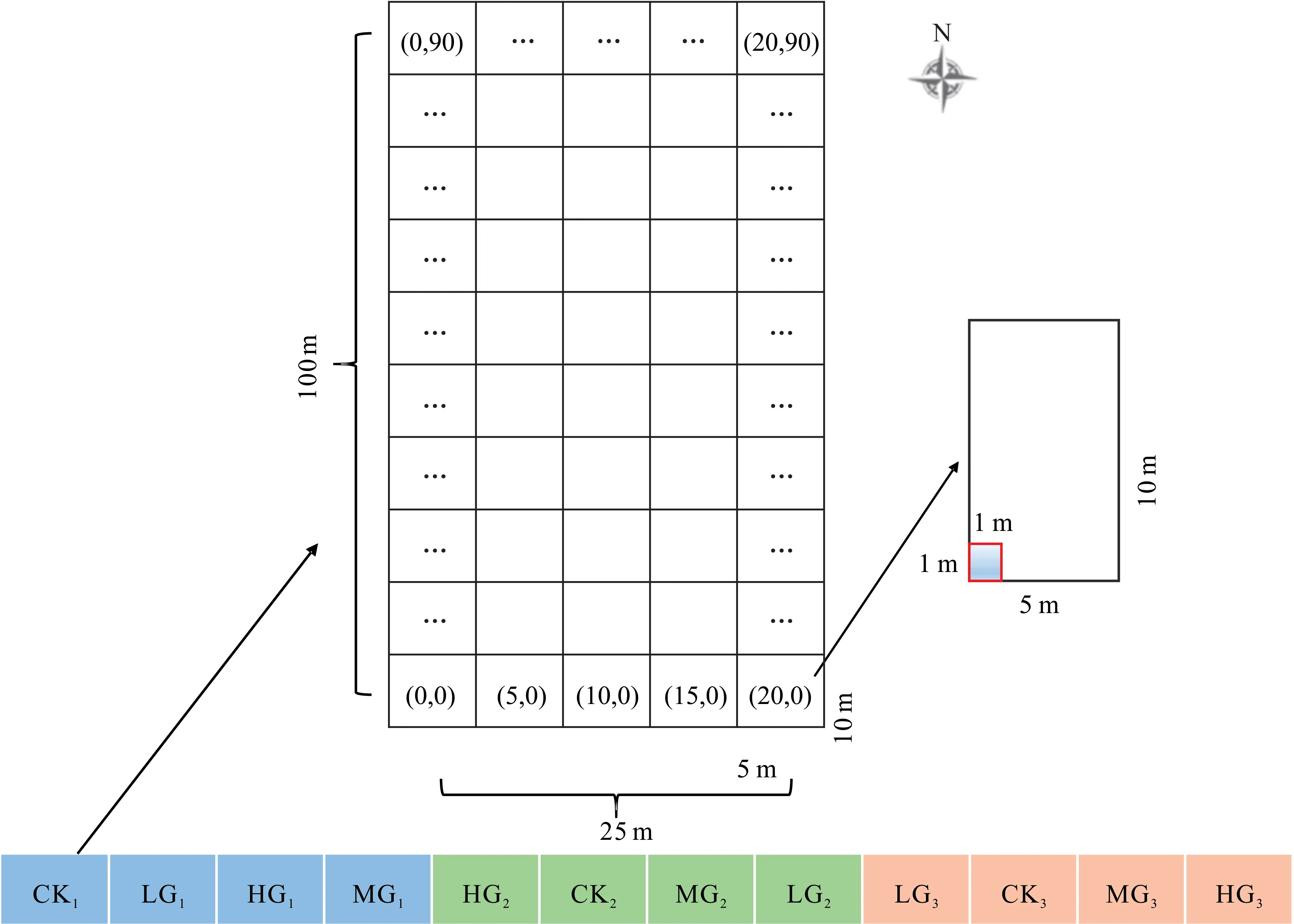
圖1 樣地及取樣示意圖Fig.1 Schematic diagram of plot and sampling
1.3 數(shù)據(jù)處理
首先,對建群種短花針茅與優(yōu)勢種無芒隱子草和冷蒿種群出現(xiàn)頻率及多度進行處理及月份下雙因素方差分析;其次,以建群種短花針茅為自變量,以優(yōu)勢種無芒隱子草和冷蒿為因變量分別進行回歸分析;再者,對優(yōu)勢種兩兩進行物種對同分布指數(shù)分析,得出6個物種對的物種作用強度;最后,將優(yōu)勢種無芒隱子草(X)和冷蒿(Y)種群多度定義為有序的坐標變量,分析建群種短花針茅(Z)受優(yōu)勢種無芒隱子草和冷蒿種群的影響特點和程度。物種多度數(shù)據(jù)均經(jīng)過正態(tài)轉(zhuǎn)化,符合正態(tài)分布后使用地統(tǒng)計學軟件GS+9.0進行半方差函數(shù)分析,得到最優(yōu)模型及優(yōu)勢種群間空間分布關(guān)系的分形維數(shù)后通過克里格(Kriging)插值法繪制其3D立體圖,通過立體圖峰谷之間的過渡及凹凸情況來判斷優(yōu)勢種群間分布關(guān)系情況。所有數(shù)據(jù)分析基于SPSS 19.0軟件,所有圖表制作基于SigmaPlot 14.0。顯著性水平為α=0.05。此外,頻率(frequency,f)及同分布指數(shù)公式(co-occurrence index,CO)[17]如下:

式中:N i代表物種出現(xiàn)的樣方數(shù);N為總樣方數(shù);O1,2代表兩個物種同時出現(xiàn)的樣方數(shù);O1代表只有物種1出現(xiàn)的樣方數(shù)。如果CO1,2值很大,說明物種2在物種1出現(xiàn)的大多數(shù)樣方都出現(xiàn),物種1受到物種2影響的可能性比較大;如果CO1,2值很小,說明在物種1分布的大多數(shù)樣方內(nèi)都沒有物種2個體,二者發(fā)生作用的可能性很小。
2 結(jié)果與分析
2.1 優(yōu)勢種群密度
對建群種短花針茅及優(yōu)勢種無芒隱子草和冷蒿種群進行分析表明,不同放牧處理、月份及其交互作用均對短花針茅和無芒隱子草出現(xiàn)頻率無顯著影響(P>0.05);而冷蒿出現(xiàn)頻率僅在處理間差異顯著(P<0.05)。然而,不同放牧處理、月份及其交互作用對短花針茅、無芒隱子草及冷蒿種群多度均存在顯著影響(P<0.05)(表1)。
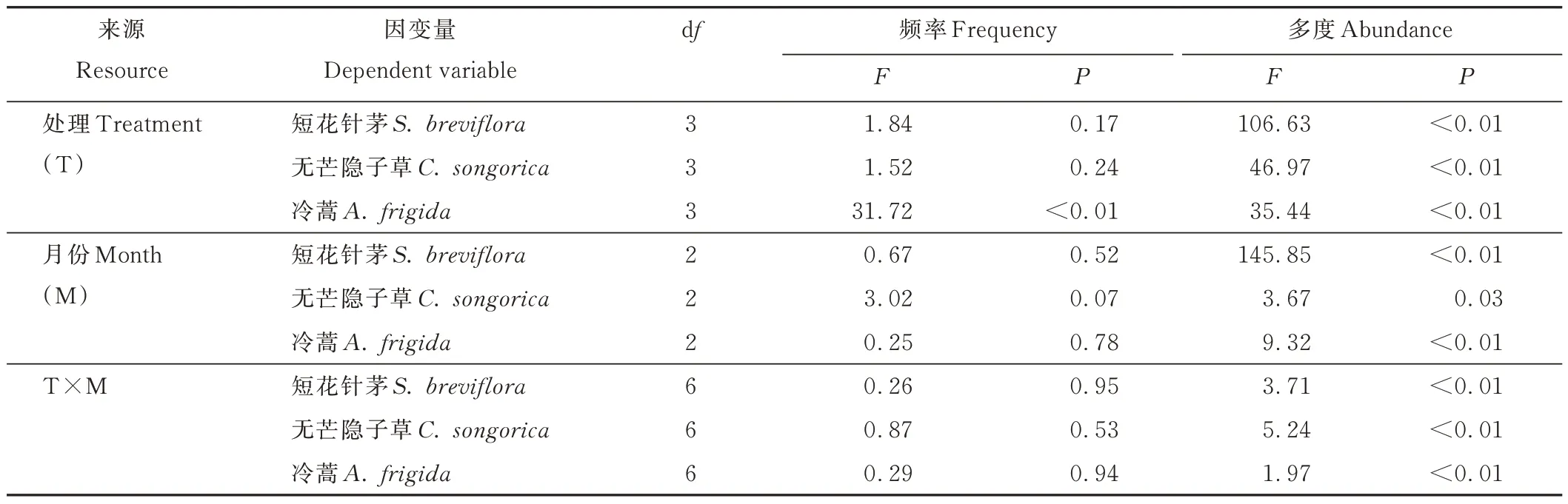
表1 優(yōu)勢種群頻度及多度的雙因素方差分析Table 1 Two-way analysis of variance for frequency and abundance of dominant population
2.2 優(yōu)勢種群出現(xiàn)頻率
不同放牧處理及月份下,短花針茅及無芒隱子草種群出現(xiàn)頻率分別為91.67%~100.00%和85.33%~100.00%(P>0.05)。不同放牧處理下冷蒿種群出現(xiàn)頻率存在顯著差異(P<0.05);與CK相比,LG僅在7月時顯著降低了50.54%(P<0.05),而MG與HG在7-9月均顯著降低(P<0.05),MG在7-9月分別降低了76.92%、77.78%和78.57%,HG則分別降低了100.00%、92.59%和100.00%;但各放牧處理在不同月份間無顯著差異(P>0.05)(圖2)。
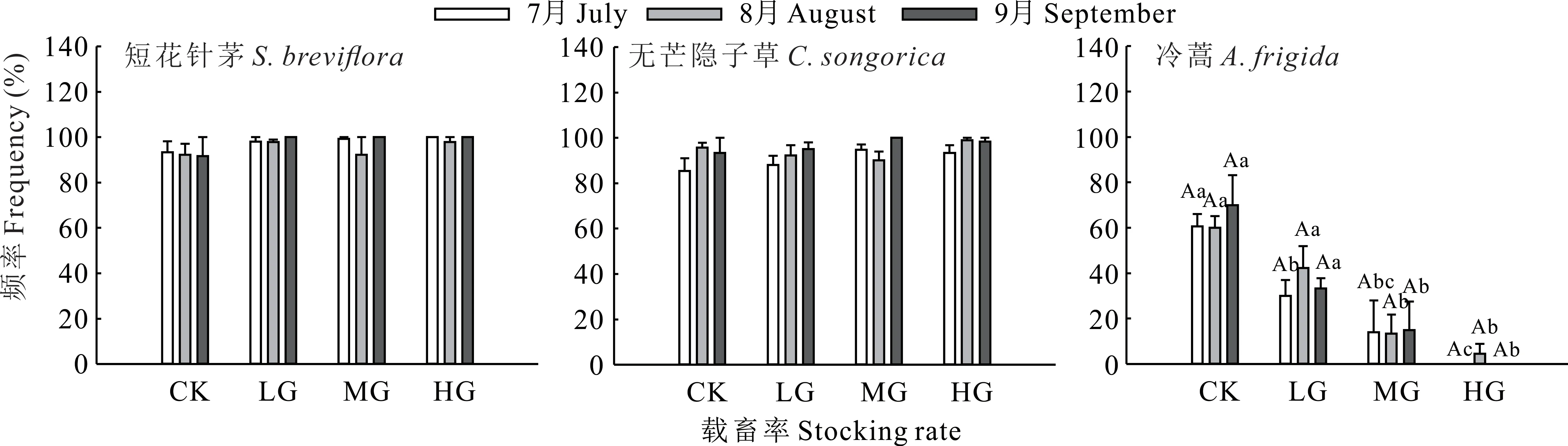
圖2 不同載畜率下優(yōu)勢物種出現(xiàn)頻率Fig.2 Frequency of dominant species under different stocking rates
2.3 種群多度
對短花針茅而言,與CK相比,種群多度在7-9月均顯著增加(P<0.05),LG、MG和HG分別在7-9月平均增加約37.34%、75.72%和99.32%;但與7月相比,8和9月短花針茅種群多度均顯著降低(P<0.05)。對無芒隱子草而言,與CK相比,LG在7和8月時顯著增加(P<0.05),MG和HG在7-9月時均顯著增加(P<0.05);與7月相比,LG和MG在8月時種群多度顯著增加(P<0.05)。對于冷蒿而言,與CK相比,各處理種群多度在7-9月均顯著降低(P<0.05),LG、MG和HG分別在7-9月平均降低約61.26%、66.32%和99.31%;與7月相比,CK、MG和HG在8月時種群多度均顯著增加(P<0.05)(圖3)。
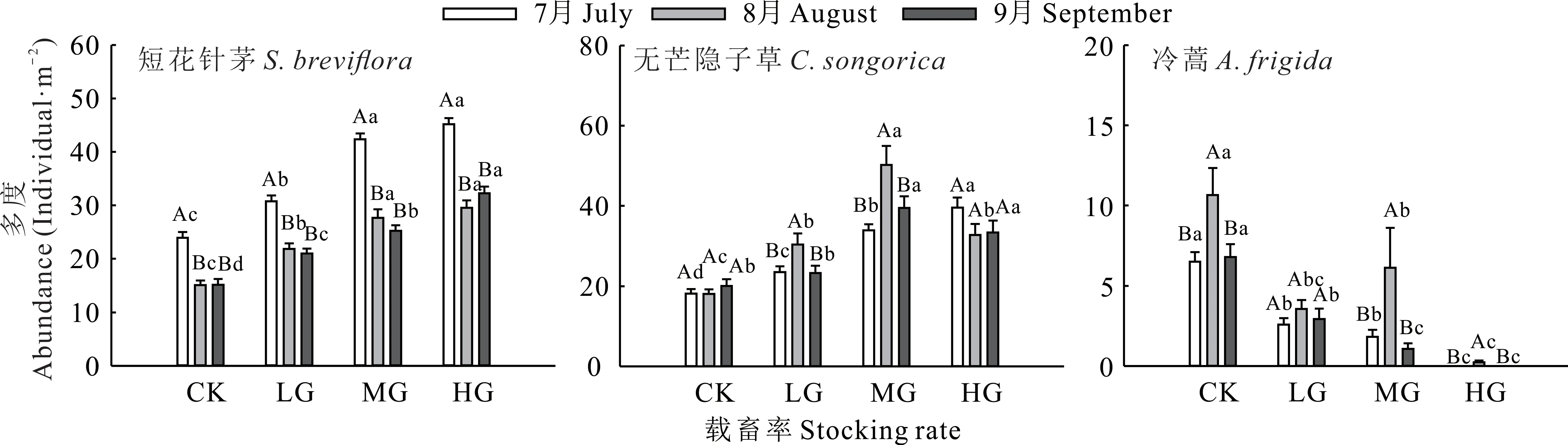
圖3 不同載畜率下優(yōu)勢物種多度Fig.3 Abundance of dominant species under different stocking rates
2.4 建群種與優(yōu)勢種間的回歸分析
7月時,短花針茅與無芒隱子草間在CK中存在顯著正相關(guān)關(guān)系(P<0.05);8月時,在MG和HG中存在顯著正相關(guān)關(guān)系(P<0.05);9月時,在CK中存在顯著正相關(guān)關(guān)系(P<0.05),在HG中存在顯著負相關(guān)關(guān)系(P<0.05)。然而,7月時,短花針茅與冷蒿間在CK和MG中存在顯著負相關(guān)關(guān)系(P<0.05);8月時,在CK、LG和MG中存在顯著負相關(guān)關(guān)系(P<0.05);9月時,在CK和LG中存在顯著負相關(guān)關(guān)系(P<0.05)(表2)。
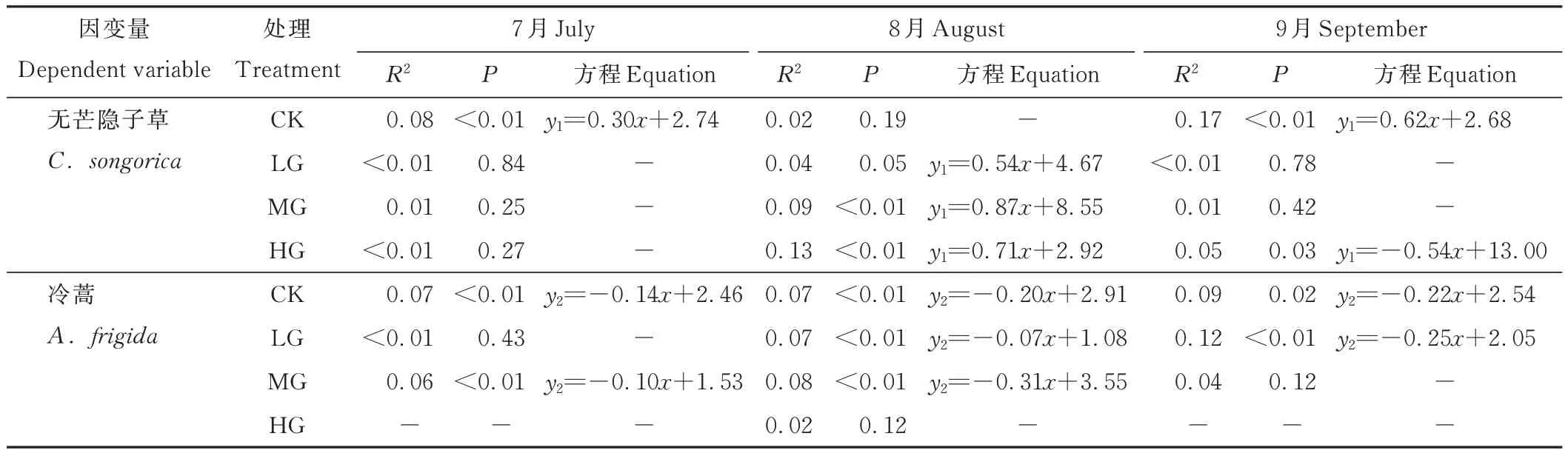
表2 建群種與優(yōu)勢種間的回歸分析Table 2 Regression analysis between the constructive species and the dominant species
2.5 物種間的同分布指數(shù)
對短花針茅、無芒隱子草和冷蒿進行兩兩物種對同分布指數(shù)分析表明,短花針茅與無芒隱子草對冷蒿的作用強度較大,而冷蒿對短花針茅和無芒隱子草的作用強度較小;此外,短花針茅、無芒隱子草和冷蒿之間的相互作用主要集中在7和8月。與CK相比,7和8月時,LG下短花針茅對無芒隱子草和冷蒿作用強度增大。與CK相比,8月時,LG和MG下無芒隱子草對短花針茅和冷蒿作用強度降低,HG下無芒隱子草對短花針茅作用強度增大。與CK相比,冷蒿對短花針茅和無芒隱子草的作用強度變化較小(表3)。
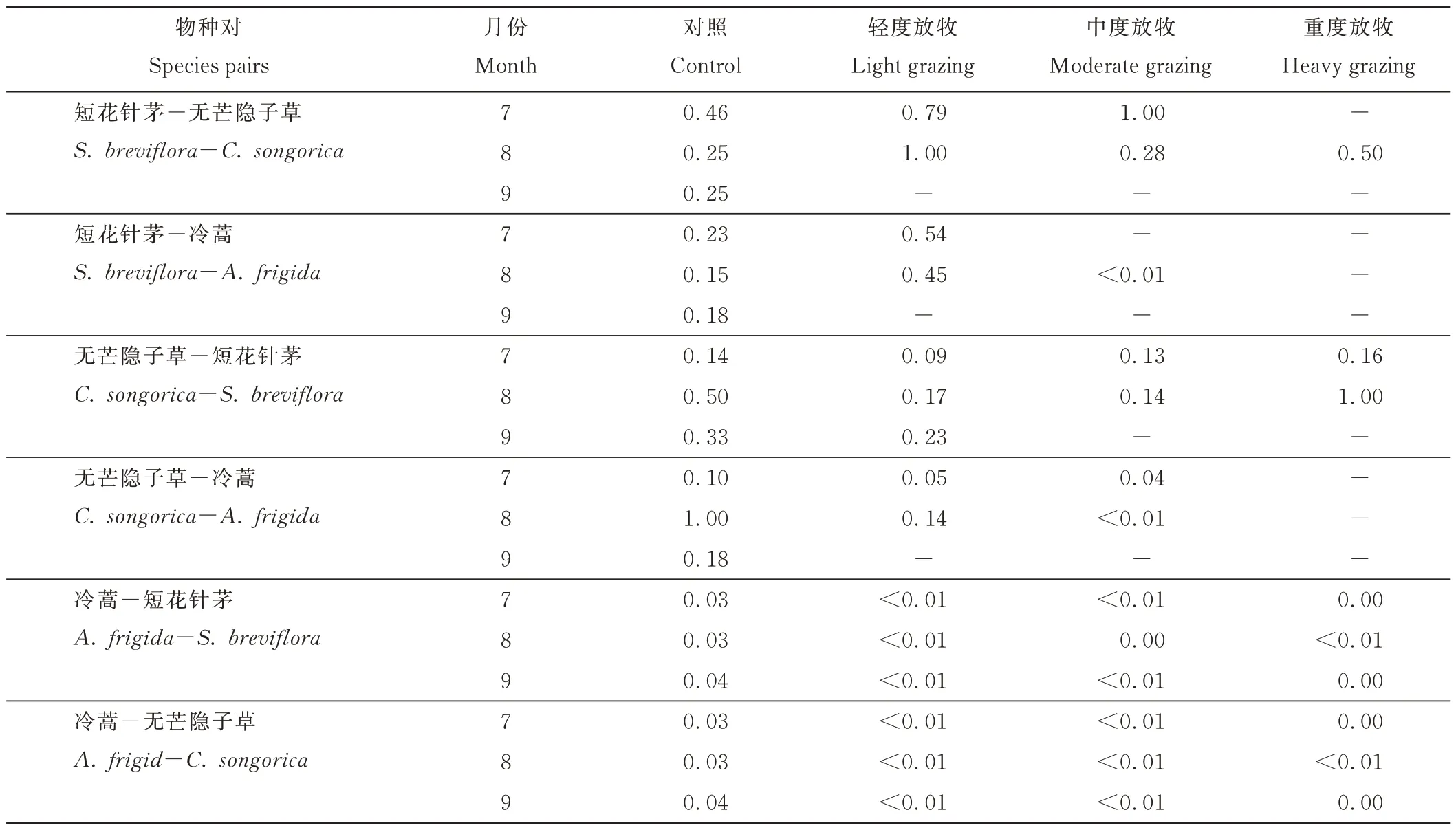
表3 物種間的同分布指數(shù)Table 3 Co-occurrence index among species
2.6 建群種與優(yōu)勢種間的空間分布
9月時,HG下無芒隱子草與短花針茅種群多度存在負相關(guān)關(guān)系(表2),而其他月份或處理中,短花針茅種群多度均與無芒隱子草種群多度為正相關(guān)關(guān)系,與冷蒿種群多度為負相關(guān)關(guān)系,故主要對9月的空間分布進行分析。結(jié)果表明,CK中,短花針茅種群多度隨無芒隱子草種群多度增加而呈增加的趨勢,隨冷蒿種群多度增加而呈降低的趨勢。LG中,當冷蒿種群多度較低時,隨著無芒隱子草種群多度的增加,短花針茅呈先增加后降低,而后增加再降低的趨勢;當冷蒿種群多度較高時,隨著無芒隱子草種群多度增加,短花針茅種群多度呈先降低后增加的趨勢。MG中,當冷蒿種群多度較低時,隨著無芒隱子草種群多度增加,短花針茅種群多度呈先增加后降低的趨勢;當冷蒿種群多度較高時,隨著無芒隱子草種群多度增加,短花針茅種群多度呈逐漸降低的趨勢。HG中,由于冷蒿出現(xiàn)較少、密度較低,導致其種群分布較少,故其種群密度在圖中形成窄條,其多度不隨短花針茅與無芒隱子草種群多度的變化而變化,隨著無芒隱子草種群多度增加,短花針茅種群多度呈先增加后降低,而再增加后再降低的趨勢,整體上呈波動下降的趨勢(圖4)。
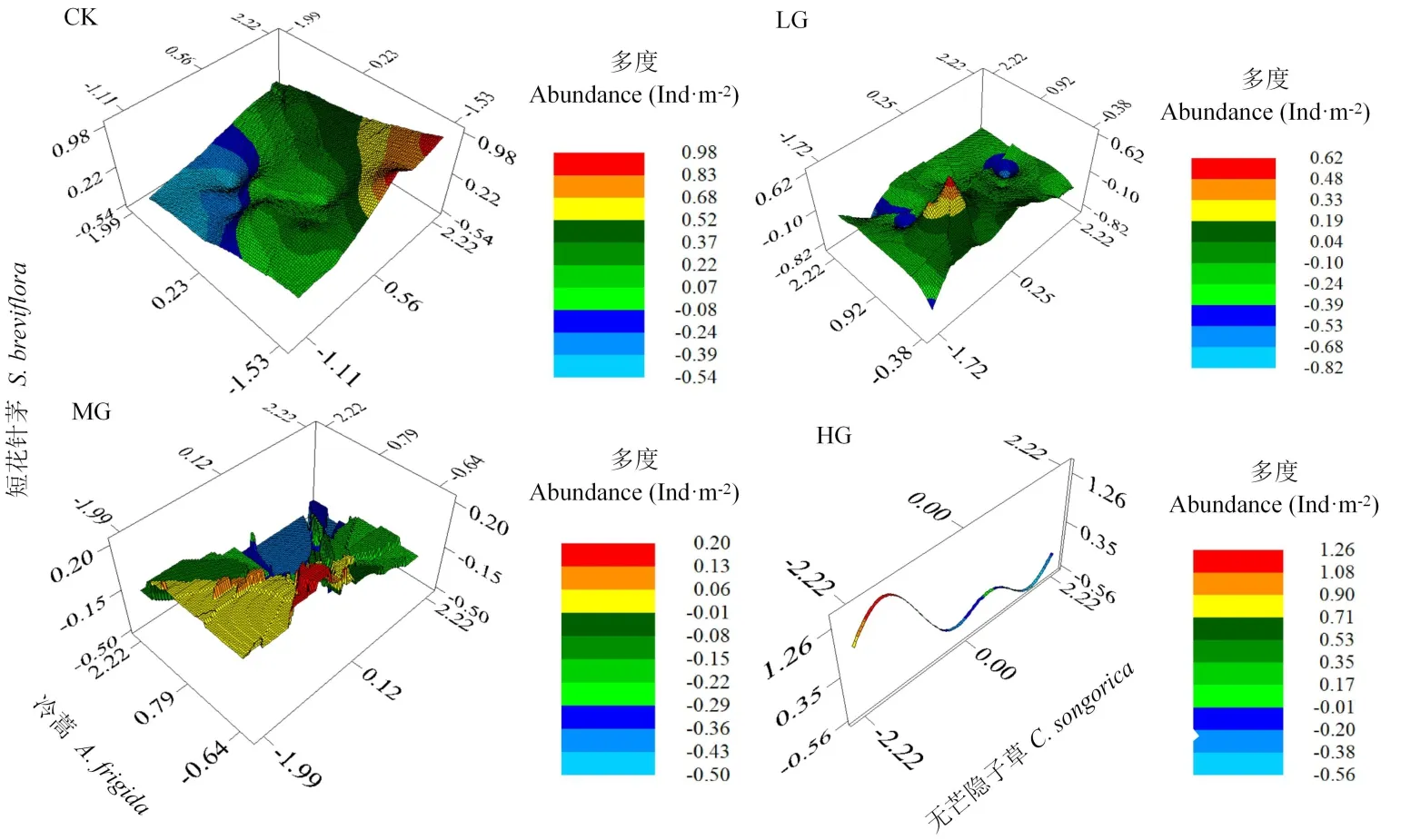
圖4 不同載畜率下建群種與優(yōu)勢種間空間分布特征Fig.4 Spatial distr ibution characteristics of constr uctive species and dominant species under different stocking r ates
3 討論
在長期放牧過程中,群落結(jié)構(gòu)及種群分布會發(fā)生改變[18]。本研究中,放牧導致短花針茅和無芒隱子草種群多度增加,冷蒿種群多度降低。一方面由于放牧會移除一部分植株而降低頂端優(yōu)勢,因而促進叢生禾草分蘗,造成株叢增大或植株密度增加[19];另一方面由于在長期放牧過程中,隨放牧強度增大,短花針茅大株叢逐漸形成若干個小株叢,脫離母株后進行營養(yǎng)繁殖使種群密度增大[20-21]。古琛等[22]在對荒漠草原建群種短花針茅生長和繁殖策略的研究也表明,隨放牧強度的增大,短花針茅種群受家畜踐踏趨于破碎化;張峰等[14]在對大針茅典型草原建群種大針茅和優(yōu)勢種羊草(Leymus chinensis)及糙隱子草(Cleistogenes squarrosa)的研究中發(fā)現(xiàn),其三者在放牧下種群密度顯著增加。本研究中,短花針茅和無芒隱子草均為多年生叢生禾草,受家畜采食和踐踏作用,使得株叢破碎、株幅變小,使其種群密度增大;一方面由于冷蒿適口性高于短花針茅和無芒隱子草,家畜喜食冷蒿造成其光合面積減少及再生能力下降,進而造成其種群多度及出現(xiàn)頻率降低;另一方面,冷蒿生殖方式與短花針茅與無芒隱子草存在差異,短花針茅與無芒隱子草受放牧干擾時出現(xiàn)矮型化、破碎化,而冷蒿主要以匍匐地面的方式降低株高以躲避家畜采食。此外,放牧家畜的過度踐踏使土壤緊實度和容重增大[23],間接阻礙了根系的伸長與擴展[24],也會對冷蒿生長與繁殖造成一定的影響。
希吉日塔娜等[25]對不同放牧制度下短花針茅荒漠草原種間關(guān)系的分析表明,種間關(guān)系在兩物種間是不對等的,種間親和關(guān)系會因放牧制度的改變而改變;本研究也發(fā)現(xiàn),短花針茅與無芒隱子草、冷蒿種群多度之間的相關(guān)性存在動態(tài)變化;此外,短花針茅與無芒隱子草對冷蒿的作用強度較大,而冷蒿對短花針茅和無芒隱子草的作用強度較小。此外,本研究發(fā)現(xiàn),短花針茅主要與無芒隱子草表現(xiàn)為親和關(guān)系,與冷蒿表現(xiàn)為競爭關(guān)系;但9月時短花針茅與無芒隱子草間關(guān)系從對照下的親和關(guān)系轉(zhuǎn)化為重度放牧下的競爭關(guān)系。吳艷玲等[26]對不同放牧強度下物種間關(guān)系研究表明,重度放牧造成物種對正相關(guān)對數(shù)減少,種間競爭與親和作用可以相互轉(zhuǎn)化。方楷等[27]對不同放牧制度下荒漠草原種間關(guān)系研究也表明,輕度放牧不會改變植物的種間關(guān)系,但重度放牧加劇了資源競爭,還會引起群落逆向演替。但張爽[28]在短花針茅荒漠草原種間關(guān)系對放牧響應(yīng)的研究中表明,放牧使得主要物種間關(guān)系向正關(guān)聯(lián)方向發(fā)展;此外,不同放牧處理下,物種間親和或競爭關(guān)系均存在不對稱性;放牧造成種內(nèi)競爭強度增加,但種間競爭強度減弱。劉菊紅等[29]對重度放牧下荒漠草原主要植物種間關(guān)系的研究表明,重牧使群落物種間關(guān)系由競爭轉(zhuǎn)變?yōu)橛H和。由于種間關(guān)系存在多度效應(yīng),因干擾條件或物種不同,植物種間關(guān)系存在多種表現(xiàn)形式[30]。
由于各物種在放牧干擾下存在不同的生存繁殖策略,而其繁殖策略是植物在其生活史中對外界干擾的響應(yīng),在不同生境中的繁殖對策反映了其對環(huán)境的適應(yīng)能力和生殖潛力[31]。短花針茅和無芒隱子草均為多年生叢生禾草,在長期放牧過程中,短花針茅和無芒隱子草株叢傾向于破碎化,種群高度傾向于矮型化[20-22];隨放牧強度增大,冷蒿通過莖的斜倚程度甚至匍匐于地面使得種群矮化來躲避家畜采食[32]。此外,草地群落中大多植物同時采取有性和無性繁殖方式來實現(xiàn)種群生長和繁殖,植物在擾動大、競爭較弱的環(huán)境中傾向于有性繁殖,而在相對穩(wěn)定的環(huán)境中傾向于無性繁殖[33-34]。隨著載畜率增大,冷蒿有性繁殖分配減少、生殖枝數(shù)和種子產(chǎn)量降低,重度放牧下有性生殖可能消失[35];而短花針茅也傾向于分蘗減少、種子減少和降低幼苗存活率。故在本研究中,重度放牧下短花針茅與無芒隱子草關(guān)系從親和變?yōu)楦偁帲砻髦囟确拍翆е路N間關(guān)系發(fā)生變化。
4 結(jié)論
放牧降低了冷蒿的出現(xiàn)頻率,但未改變短花針茅和無芒隱子草的出現(xiàn)頻率;放牧使得短花針茅和無芒隱子草種群多度增加,造成冷蒿種群多度降低。其次,短花針茅主要與無芒隱子草表現(xiàn)為親和關(guān)系,與冷蒿表現(xiàn)為競爭關(guān)系;但9月時短花針茅與無芒隱子草間關(guān)系從對照下的親和關(guān)系轉(zhuǎn)化為重度放牧下的競爭關(guān)系,短花針茅與冷蒿間關(guān)系無變化。此外,通過對9月物種間空間分布進行模擬也發(fā)現(xiàn)短花針茅與無芒隱子草之間關(guān)系由對照中的親和關(guān)系轉(zhuǎn)變?yōu)橹囟确拍料碌母偁庩P(guān)系,表明利用地統(tǒng)計學對其物種間空間分布分析較合理。
猜你喜歡
課堂內(nèi)外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創(chuàng)造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(shù)(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
